Environmental Risk Assessment of Living Modified Microorganisms (LMM) on the Indigenous Microbial Community


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Condition
2.2. Collection of Field Soil Samples
2.3. Physicochemical Characterization of the Collected Soil Samples
2.4. Analysis of LMM (SEM002) Survival in the Soil and Effect on the Soil Microbial Community
2.5. Analysis of Changes in the Density of Major Soil Microbial Communities
2.6. Detection of Remained Genetic Material of SEM002 and Wild-Type Strains
2.7. Analysis of Changes of Soil Microbial Communities by Real-Time PCR-DGGE
3. Results
3.1. Characteristics of Soil Samples
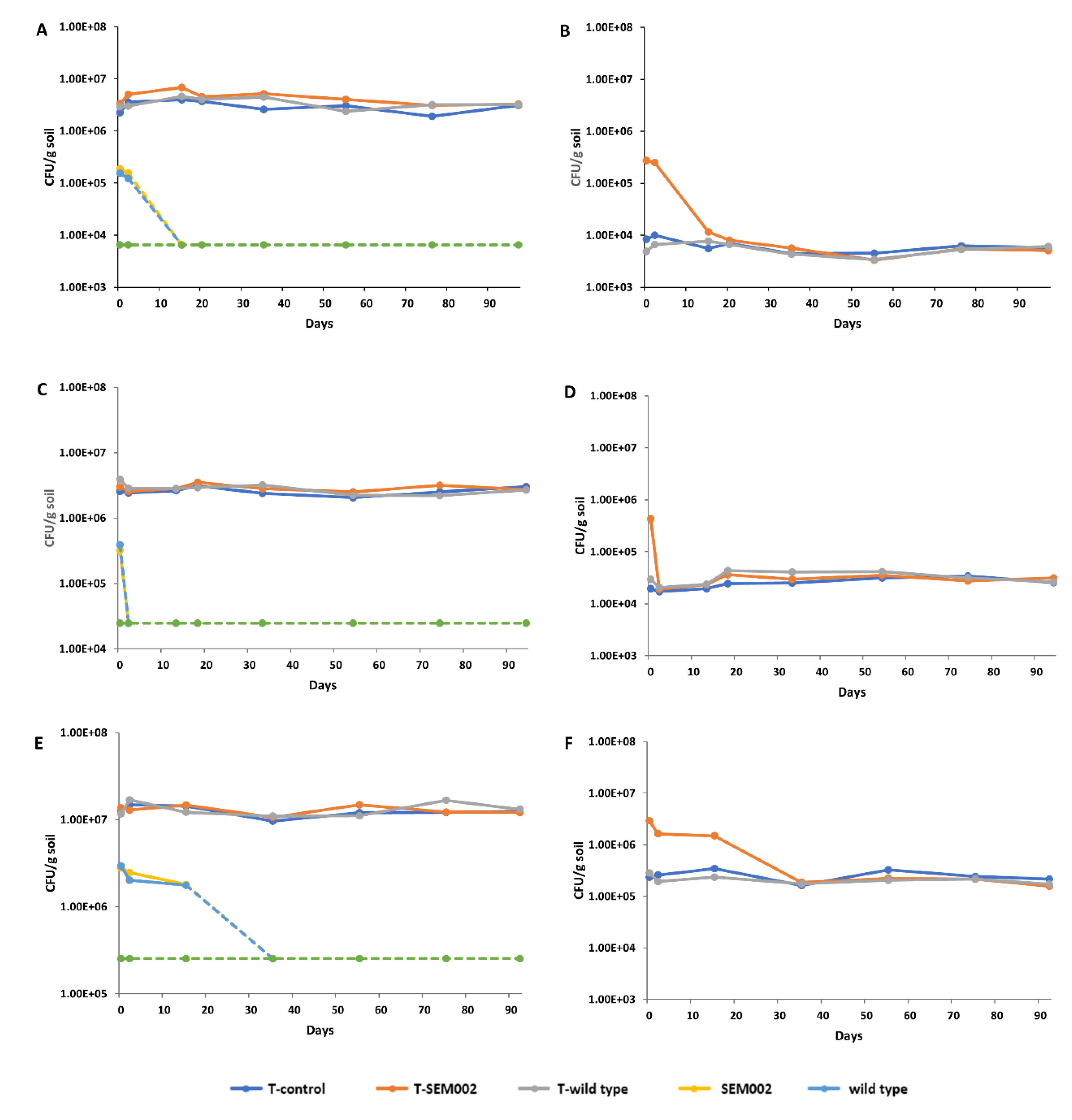
3.2. Survival of LMMs in Soil Samples
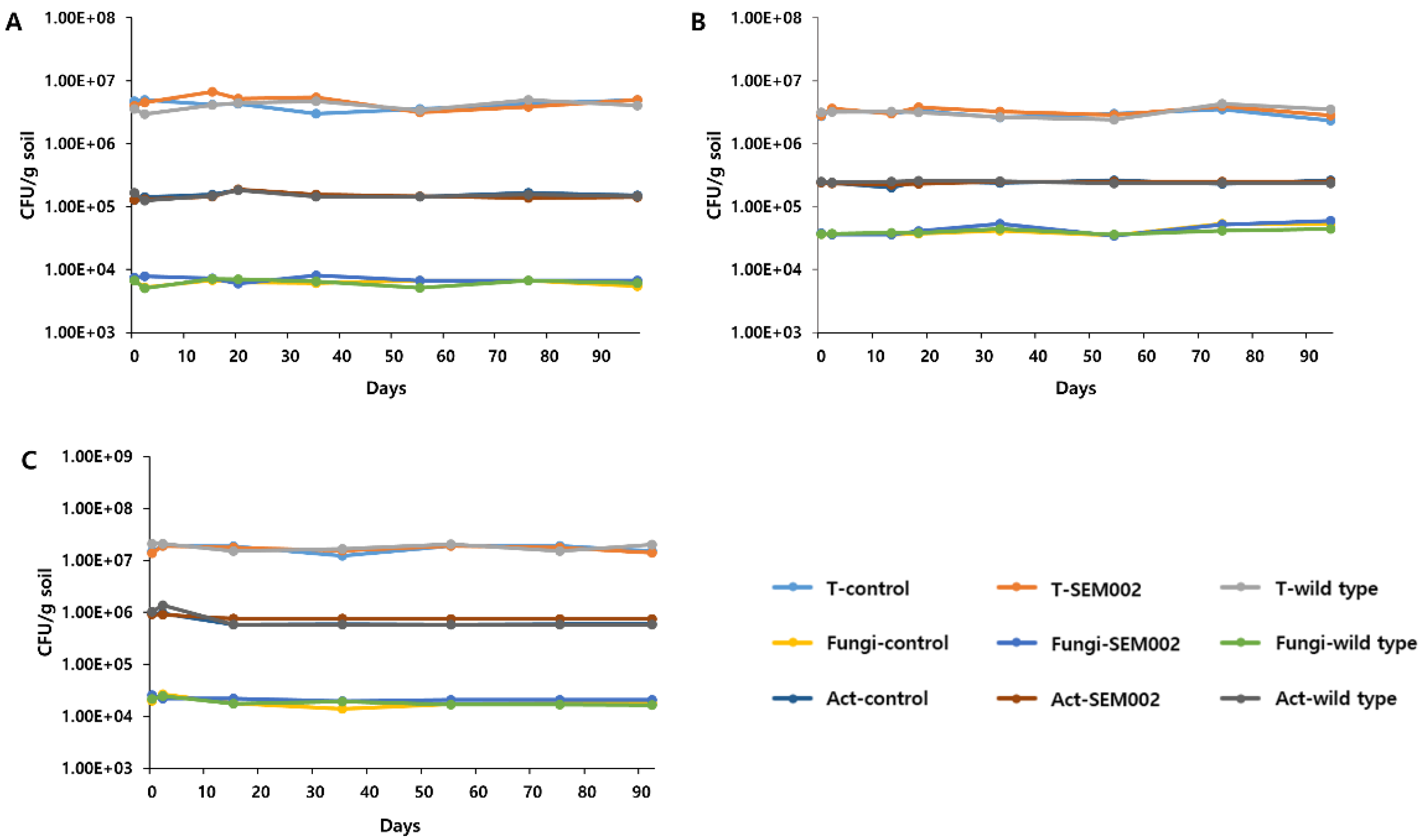
3.3. Changes in the Density of the Major Soil Microorganisms Upon LMM Release
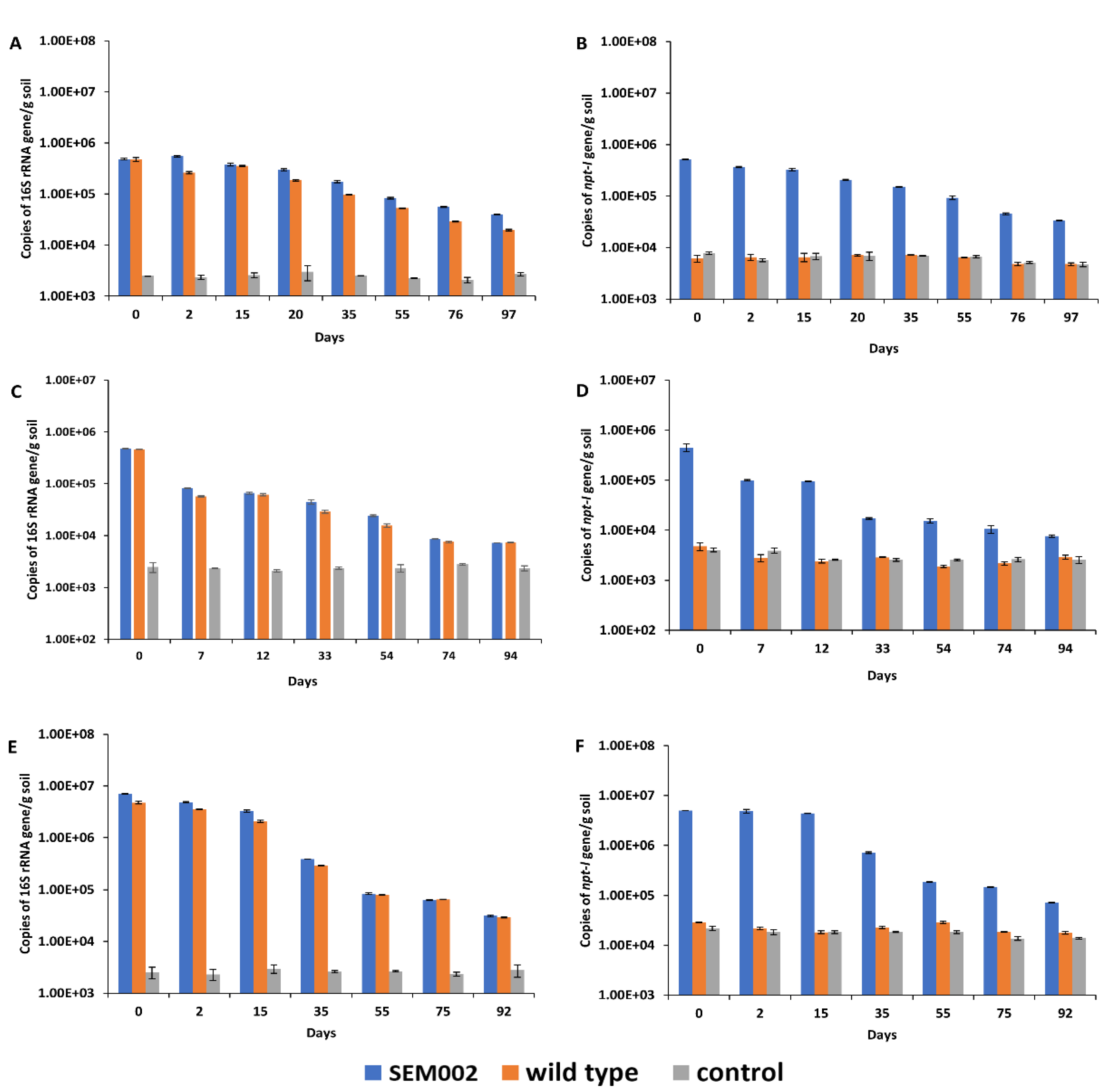
3.4. Changes in the Copy Number of the 16S rRNA and Kanamycin Resistance Gene in Inoculated Microcosm Soil
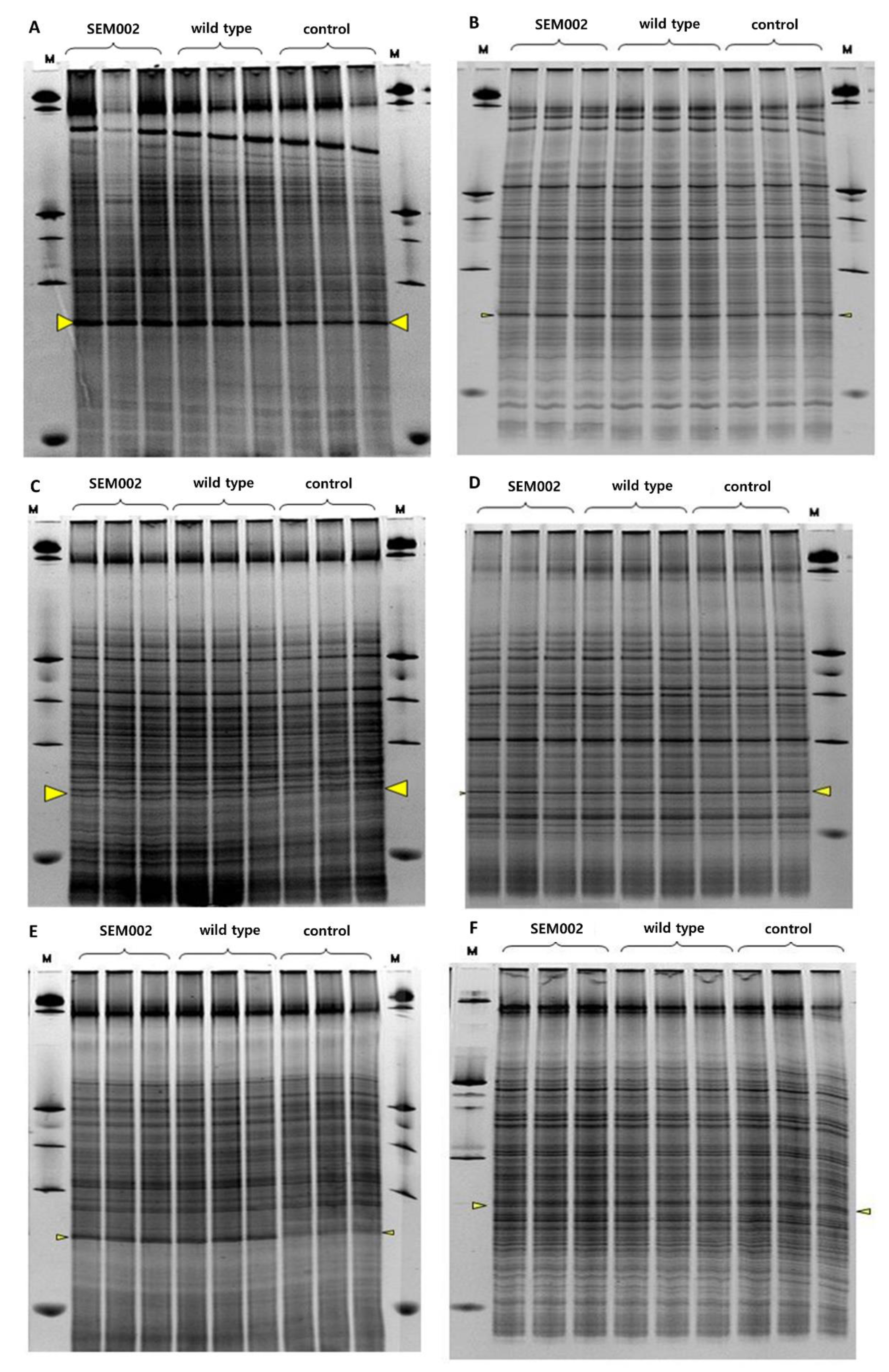
3.5. Effect of LMM on the Indigenous Microbial Community Structure in Soil
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, C.; Wohlhueter, R.; Zhang, H. Genetically modified foods: A critical review of their promise and problems. Food Sci. Human Wellness 2016, 5, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Schiemann, J.; Dietz-Pfeilstetter, A.; Hartung, F.; Kohl, C.; Romeis, J.; Sprink, T. Risk Assessment and Regulation of Plants Modified by Modern Biotechniques: Current Status and Future Challenges. Annu. Rev. Plant Biol. 2019, 70, 699–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallikarjuna, N.; Yellamma, K. Genetic and Metabolic Engineering of Microorganisms for the Production of Various Food Products. In Recent Developments in Applied Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 2019; pp. 167–182. [Google Scholar]
- Engwa, A.G. Genetic engineering on microorganism: The ecological and bioethical implications. Eur. J. Biotechnol. 2014, 1, 27–33. [Google Scholar]
- Ando, A.; Suzuki, C.; Shima, J. Survival of genetically modified and self-cloned strains of commercial baker’s yeast in simulated natural environments: Environmental risk assessment. Appl. Environ. Microbiol. 2005, 71, 7075–7082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, I.; Lee, T.K.; Han, J.; Doan, T.V.; Kim, S.B.; Park, J. Improved detection of microbial risk of releasing genetically modified bacteria in soil by using massive sequencing and antibiotic resistance selection. J. Hazard. Mater. 2012, 227–228, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Janczyk, P.; Pieper, R.; Smidt, H.; Wolfgang, B. Souffrant effect of alginate and inulin on intestinal microbial ecology of weanling pigs reared under different husbandry conditions. FEMS Microbiol. Ecol. 2010, 72, 132–142. [Google Scholar] [CrossRef] [PubMed]
- De Vos, B.; Lettens, S.; Muys, B.; Deckers, J.A. Walkley–Black analysis of forest soil organic carbon: Recovery, limitations and uncertainty. Soil Use Manag. 2007, 23, 221–229. [Google Scholar] [CrossRef]
- Guebel, D.V.; Nudel, B.C.; Giulietti, A.M. A Simple and rapid micro-Kjeldahl method for total nitrogen analysis. Biotechnol. Tech. 1991, 5, 427–430. [Google Scholar] [CrossRef]
- Ruiz-Calero, V.; Galcera, M.T. Ion chromatographic separations of phosphorus species: A review. Talanta 2005, 66, 376–410. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.L. Actinomycetes in Danish soils. Soil Sci. 1930, 30, 59–77. [Google Scholar] [CrossRef]
- Ahn, J.H.; Kim, Y.J.; Kim, T.S.; Song, H.G.; Kang, C.H.; Ka, J.O. Quantitative improvement of 16S rDNA DGGE analysis for soil bacterial community using real-time PCR. J. Microbiol. Methods 2009, 78, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N. Bulk and Rhizosphere Soil Bacterial Communities Studied by Denaturing Gradient Gel Electrophoresis: Plant-Dependent Enrichment and Seasonal Shifts Revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.C.; Ahn, J.H.; Shin, H.C.; Kim, T.; Ryu, T.H.; Kim, D.H.; Song, H.G.; Lee, G.H.; Ka, J.O. Molecular Analysis of Bacterial Community Structures in Paddy Soils for Environmental Risk Assessment with Two Varieties of Genetically Modified Rice, Iksan 483 and Milyang 204. J. Microbiol. Biotechnol. 2008, 18, 207–218. [Google Scholar] [PubMed]
- Bruinsma, M.; Kowalchuk, G.A.; Van Veen, J.A. Effects of genetically modified plants on microbial communities and processes in soil. Biol. Fertil. Soils 2003, 37, 329–337. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Yu, C.; Bao, W.; Dong, S.; Yu, R.; Chen, M.; Wang, C. Genetically modified WYMV-resistant wheat exerts little influence on rhizosphere microbial communities. Appl. Soil Ecol. 2016, 105, 169–176. [Google Scholar] [CrossRef]
- Ma, B.L.; Blackshaw, R.E.; Roy, J.; He, T. Investigation on gene transfer from genetically modified corn (Zea mays L.) plants to soil bacteria. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2011, 46, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Hagi, T.; Minagawa, A.; Shima, J. Dynamics of genetically modified Lactococcus lactis in simulated natural environments and impacts on microbial communities. J. Biosci. Bioeng. 2009, 107, 339–343. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factory Soil | Forest Soil | Sedimentary Soil | |
|---|---|---|---|
| pH | 7.83 | 4.37 | 6.95 |
| Organic content (%) | 1.03 | 7.21 | 3.88 |
| T-N (%) | 0.0449 | 0.2679 | 0.1657 |
| Exchangeable Ca (mg/kg) | 871.50 | 159.30 | 755.3 |
| Exchangeable Mg (mg/kg) | 31.52 | 21.34 | 58.56 |
| Exchangeable K (mg/kg) | 93.51 | 94.72 | 80.76 |
| Exchangeable Na (mg/kg) | 501.50 | 456.70 | 500.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Kim, D.-U.; Son, J.; Kim, S.-B.; Ka, J.-O. Environmental Risk Assessment of Living Modified Microorganisms (LMM) on the Indigenous Microbial Community. Sustainability 2020, 12, 5566. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145566
Lee H, Kim D-U, Son J, Kim S-B, Ka J-O. Environmental Risk Assessment of Living Modified Microorganisms (LMM) on the Indigenous Microbial Community. Sustainability. 2020; 12(14):5566. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145566
Chicago/Turabian StyleLee, Hyosun, Dong-Uk Kim, Jigwan Son, Seong-Bo Kim, and Jong-Ok Ka. 2020. "Environmental Risk Assessment of Living Modified Microorganisms (LMM) on the Indigenous Microbial Community" Sustainability 12, no. 14: 5566. https://0-doi-org.brum.beds.ac.uk/10.3390/su12145566