Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production





1
College of Animal Husbandry & Veterinary Sciences, Abdul Wali Khan University, Mardan 23200, Pakistan
2
Faculty of Animal Husbandry & Veterinary Sciences, College of Veterinary Sciences, The University of Agriculture, Peshawar 25130, Pakistan
3
Department of DETO, Section of Veterinary Science and Animal Production, University of Bari ‘Aldo Moro’, Valenzano, 70010 Bari, Italy
*
Authors to whom correspondence should be addressed.
Sustainability 2020, 12(24), 10693; https://0-doi-org.brum.beds.ac.uk/10.3390/su122410693
Submission received: 24 November 2020
/
Revised: 16 December 2020
/
Accepted: 17 December 2020
/
Published: 21 December 2020
(This article belongs to the Special Issue Animal Nutrition and Welfare in Sustainable Production Systems)
Abstract
:This review article discusses the importance and effects of Selenium (Se) in sustainable dairy cows’ production. The Se is an important micronutrient in dairy cows. It is one of the important feed derived antioxidants. It participates in important enzymes and enzyme reactions to improve metabolism, growth, and the defense system of the body, which results in the improved health of animals, particularly that of the mammary gland and reproductive system, thereby improving productive and reproductive performance. The Se is usually deficient in soil due to current extensive farming strategies, so its supplementation is generally advised. Supplementation of Se in organic form is generally preferred over inorganic form due to its better incorporation and uptake, resulting in improved performance. Kidneys, liver, testis, and lungs are important sites for Se storage. The Se is excreted in urine, feces, exhaled breath, loss of skin, and hair cells. Although Se supplementation plays an important role in the profitability of dairy cows, its excess intake is toxic and should be avoided.
1. Introduction
Selenium (Se) is a nonmetal with the atomic number 34. In the periodic table, the element is located in the fourth period [1,2]. Se was discovered by a Swedish chemist, Jons Jacob Berzelius, in the year 1817 [3]. It is regarded as trace element due to its low content (0.05 ppm) in the earth’s crust [4,5]. The Se can occur in organic as well as inorganic form [6]. Inorganic forms of Se include metal selenides, elemental selenium, and selenates (IV) and (VI). Organic forms of Se include selenium amino acids, methyl compounds, selenoproteins, selenocysteine, and selenomethionine [7].
Se was first reported as an essential nutrient for animals by Schwarz and Foltz [8]. In biological samples, Se is present in the form of selenomethionine and selenocysteine [9]. Se plays an important role in animals and human health [10]. In the human body, Se plays a significant role in various biochemical and physiological processes [11].
In farm animals, adequate Se intake prevents various disorders like white muscle disease, mulberry heart disease, dietary necrotic liver degeneration, parturition problems, retention of placenta, post-parturient paraplegia, and early embryonic mortality [12]. In cattle, Se supplementation reduces postpartum reproductive disorders like ovarian cysts and metritis [13]. Se also plays an important role in udder health, preventing clinical and subclinical mastitis [14]. Se is a structural component of important proteins involved in defense mechanisms [15]. Further, Se occupies an important part in these proteins [16]. The body cannot produce Se by itself. Forage is a natural source of Se for animals [17]. Selenium is usually deficient in the soil, and the current extensive farming strategies tend to favor Se deficiency [18]. The soil Se concentration differs to a great extent even within small areas.
2. Absorption, Distribution, Metabolism, and Excretion of Se
Mechanisms of Se homeostasis are important because the element is potentially toxic as well as an essential micronutrient [21]. The Se is absorbed from the small intestine. Organic Se is obtained by the body from selenized yeast and basal feed ingredients in the form of seleno-amino acids (i.e., selenocysteine and selenomethionine), while inorganic selenium supplementation provides selenate and selenite. Seleno-methionine (Se-Methionine) is absorbed via methionine transporter system, selenate is absorbed by active transport system while absorption of selenite mostly takes place by passive diffusion [22]. Rate of absorption of organic Se is higher than inorganic Se [23]. Inorganic Se is mostly reduced to insoluble elemental Se or is readily absorbed into feed particles in the digestive tract [24], resulting in excretion of most of the Se [25]. In contrast, most of organic Se (in form of Se-Methionine) is incorporated in rumen microorganisms so less elemental Se is formed.
Oral bioavailability of Se-Methionine is, therefore, greater than that of inorganic Se [26]. In blood, Se is bound to low density lipoprotein (LDL), very low-density lipoprotein (VLDL), albumin, and α and β-globulins [27]. Most of the Se in body fluids and tissues is present either in the form of Se-methionine or selenocysteine [28]. In cattle, kidneys are the site of the highest density of Se, and muscles are the site of the highest concentration of Se [29]. The highest amount of Se is stored in the kidneys followed by the liver, testis, and lungs [30]. Liver, heart, and skeletal muscles are most sensitive to Se deficiency [29]. The liver is considered the Se storage organ to which a considerable amount of Se is directed during absorption for accumulation [31]. In cattle, concentration of Se rises in serum two to six days following increased supplementation in diet [32].
Various organic and inorganic Se sources are first transformed to inorganic selenide before synthesis of Se-cysteine which, in turn, contributes to the bioactive component of Selenoproteins. Following absorption, Se-Methionine can be found in plasma methionine pool and blood proteins as it is transported to body tissues. One of example of the transportation is extraction of large amounts of methionine by the mammary gland for milk protein synthesis, resulting in a large quantity of Se in milk, which is beneficial to the neonate and human consumers [33]. With an increase in Se intake in Se deficient animals, the concentration of Se in tissues and the whole body rises sharply due to accumulation of Se in tissues as selenoproteins until adequate body Se status is achieved. However, the rate of Se excretion increases once the body requirement for Se has been fulfilled, thereby reducing its further accumulation [34]. The liver plays an important role in Se excretion [35]. Excretion of Se is important for regulation of whole-body Se [34]. Se is excreted in urine, feces, exhaled breath, loss of skin, and hair cells [36]. Polymethylation of selenide takes place before excretion. Dimethyl selenide is excreted in feces and breath, while the cation (CH3)3Se+ is excreted in urine [31]. With an increase in the amount of dietary Se supplementation, the concentration of Se excretion in urine and feces increases. However, the form of Se intake does not influence the concentration of Se excreted [37].
3. Role of Se in Antioxidant Defense Mechanism
Oxidation is a process in which loss of electrons occurs [38]. Oxidation reactions are very important for life, but they can also have detrimental effects [39]. These reactions may produce reactive oxygen species (ROS) [40]. ROS can cause oxidative damage and cell death in case of excessive production [41]. ROS damage macromolecules of the cells leading to lipid peroxidation, nucleic acid, and protein alterations. Formation of ROS is considered as a pathobiochemical mechanism involved in progression or initiation phase of various diseases. To maintain correct cell signaling, radical scavenging enzymes must maintain a threshold level ROS inside the cell. Otherwise, increased ROS production not only causes excessive signals to the cells, but also directly damages key components in signaling pathways [42]. Animals with high productive performance are more susceptible to oxidative stress, resulting in their lower productive and reproductive performance [43].
Under stress conditions, the antioxidant system of the body requires dietary antioxidant supplementation via water/feed, as it cannot deal with excessive ROS formation properly [44]. Antioxidant is a natural or man-made substance that may prevent or delay some types of cell damage [39]. Antioxidants play a significant role in the body’s defense system against reactive oxygen species (ROS) [45]. Se is one of the important feed derived antioxidants [43]. Se is an important part of 25 selenoproteins identified in animals [43]. More than half of these proteins are involved in redox balance and antioxidant defense of the body [44]. Important families of selenoproteins include iodothyronine deiodinases, thioredoxin reductases, and glutathione peroxidases (GPX) [45]. The GPX, for example, protects cells against oxidative injury [46]. Activity of this enzyme is better in cows fed Se supplemented diets compared to cows fed Se deficient diets [47,48,49].
To kill phagocytized bacteria, neutrophils must provide a high oxidizing intracellular environment, but at the same time they must also maintain a balance between reactive oxygen metabolites (H2O2 and superoxide [O2−]) so that cell damage and death can be avoided [33]. In neutrophils of Se deficient cattle, GPX activity and oxygen consumption is lower than normal [50]. This results in oxidative injury of body cells [46]. Due to healthy GPX activity, destruction of cellular proteins and necrosis can be avoided, as it neutralizes the effects of lipid hydroperoxide and hydrogen peroxide [51]. Organic Se is superior to inorganic Se in improving antioxidant status in dairy animals [44].
4. Role of Se in Milk Production
General health of the animal, health of the mammary gland, environmental conditions such as insufficient nutrient intake, intensity of production system, and low corporal condition directly influence milk production [52]. Production can be improved in lactating dairy cows by supplementation of organic trace minerals [53]. Se is one of the trace minerals [54]. It is an essential element in ruminant nutrition [54,55]. Deficiency of Se impairs the immune function, leading to a higher risk of illness including that of the udder, which results in lower milk production [56].
Se helps to decrease linear somatic cell count scores [14]. Se affects innate and adaptive immune responses of the mammary gland through humoral and cellular activities [57]. It improves the bactericidal effects of milk and blood neutrophils [58]. It, therefore, prevents the incidence of mastitis [59]. In short, higher milk production in response to supplementation of Se is due to improvement in immune system brought about by Se [60].
Se yeast also improves feed digestibility, resulting in improved milk production [61]. There is a better incorporation and uptake of selenium when it is supplemented as selenium yeast [56]. Yeast based Se retains higher in tissues than inorganic Se and ensures sufficient Se availability for disease reduction [62]. Supplementation of Se in yeast form increases milk production by 24.8% [63]. When Se is supplemented along with vit E, a payback of 0.21 US cents/animal/day can be increased [64].
5. Role of Se in Feed Intake, Feed Utilization, and Body Growth
Feed intake is a critical factor for milk production of dairy cattle. The intake generally declines under heat stress to reduce metabolic heat production [65]. Feeding management is one of the key factors to enhance feed intake of dairy cows. [66]. Supplementation of diet with supranutritional Se and vit E enhances feed intake in heat stress condition by improving antioxidant status and thyroid hormone activity [67]. Se supplementation also improves growth performance in heifers, especially during the early stages of growth [68]. Se participates in important enzymes and enzyme reactions [69,70]. For example, Se helps in the expression of iodothyronine deiodinases, which regulate active T3 hormone production in peripheral tissues and the thyroid gland to improve metabolism and growth [71]. Thyroid peroxidase is also a selenoenzyme important for the process of iodization of globulin, thus avoiding damage to the thyroid epithelial cell membrane [72,73].
Supplementing Se enhances organic matter (OM) and crude protein (CP) intake [74]. Se supplementation improves total digestible nutrients (TDN) intake [68]. A higher TDN intake improves microbial efficiency in cattle. Due to differences in metabolism, the organic form of Se has better bioavailability compared to the inorganic form [75]. Supplementation of Se in yeast form improves digestibility of dry matter (DM), organic matter (OM), crude protein, neutral detergent fiber (NDF), and acid detergent fiber [76]. Se yeast improves feed digestibility in lactating dairy cows [61].
6. Role of Se in Cattle Reproductive Performance
Reproductive performance, such as age at first calving and calving interval, are important for improving productivity and profitability of dairy cows [77]. Trace minerals play very important roles in improving reproductive health and performance [78]. Fertility in dairy cows is getting worse in recent years, so enhancement of the conception rate has become an important issue. Although multiple factors are responsible for the declining fertility, improving the function of corpus luteum (CL) is one of the focuses of recent studies [79]. The function of CL is very important for reproduction according to many researchers [79]. For example, it was found that a reduced level of progesterone (P4) in plasma during the ovulatory follicles’ growth is associated with decreased conception rate [80], and a low level of plasma progesterone in the blood was associated with low survivability of the embryo during early pregnancy [81]. Progesterone concentration in blood plasma of dairy cows supplemented with selenium enriched yeast shows an early increase compared to cows to which no selenium is fed [79]. Se increases proliferation of bovine corpus luteal cells by degrading lipid peroxides resulting in increased progesterone concentration [82]. This improved luteal function helps in preventing early embryonic death [83]. So, Se supplementation improves fertility by reducing embryonic death during the first month of gestation [13].
Se supplementation also improves estrous percentage and results in lower age at first conception [84]. Supplementation of diet with organic Se in lactation and transition period improves the immune function and health of the uterus followed by improved reproductive performance [85]. Pregnancy enhances oxidative stress and vitamin E, and Se supplementation reduces the stress [86]. Deficiency of Se is usually associated with a high risk of metritis, retention of placenta, abortion, increased susceptibility to infections, and lower fertility [87]. Se supplementation improves the rate of reproduction. However, results are best when Se is supplemented as Se yeast. There is a better incorporation and uptake of selenium when it is supplemented as Selenium yeast [56].
7. Toxicity and Concluding Remarks
Se is an indispensable micronutrient in animals and is a fundamental part of seleno-enzymes. However, its excess is toxic for animals [88]. Toxicity of Se was described long before its actual discovery. In 1295, Marco Polo observed that in the north-eastern region of China, hooves of animals consuming a particular type of plant became deformed [89]. At the turn of the nineteenth century, it was discovered by US researchers that similar situation arose in cattle due to eating plants containing Se in large quantities [10,90].
Different factors, like routes of administration of Se, species of animals, and chemical forms of the element, influence the severity of Se poisoning. The soluble salts of Se (Na2SeO3 and Na2SeO4) are among the more toxic compounds; seleno-aminoacids and Se inherent in grains are relatively moderately toxic; the poorly soluble forms (e.g., diphenyl selenide, SeS2, Na2Se, and elemental Se) are among the least toxic of the Se compounds. Oral administration of Se poses less chances of toxicity than parental administration [91].
Chronic selenosis, often called alkali disease and acute selenosis, popularly known as blind staggers, are the most common form of selenosis [12]. Animals consuming feed containing Se at the rate of 5–8 mg/kg of dry matter are at risk of Se toxicity [92,93]. Livestock within 1 km of toxic farms is at risk of the toxicity. Therefore, blood samples from the livestock within 5 miles of known toxic farms should be tested for levels of Se before supplementation with the mineral [94].
Acute poisoning is not common due to the unpalatability of plants with high Se levels. However, hungry animals may eat such plants and thus suffer from acute poisoning. In cattle, acute poisoning is characterized by watery diarrhea, abnormal posture and movement, labored respiration, indications of abdominal pain, elevated temperature, prostration, and death. Chronic selenium poisoning may result from consumption of seleniferous feed over prolonged periods such as weeks or months. Important signs of the poisoning in cattle include hoof malformations, emaciation, loss of hair, and lameness [95]. In advanced cases, atrophy of the heart, anemia, and liver cirrhosis occur [91]. Excess Se intake impairs some immunological functions [96].
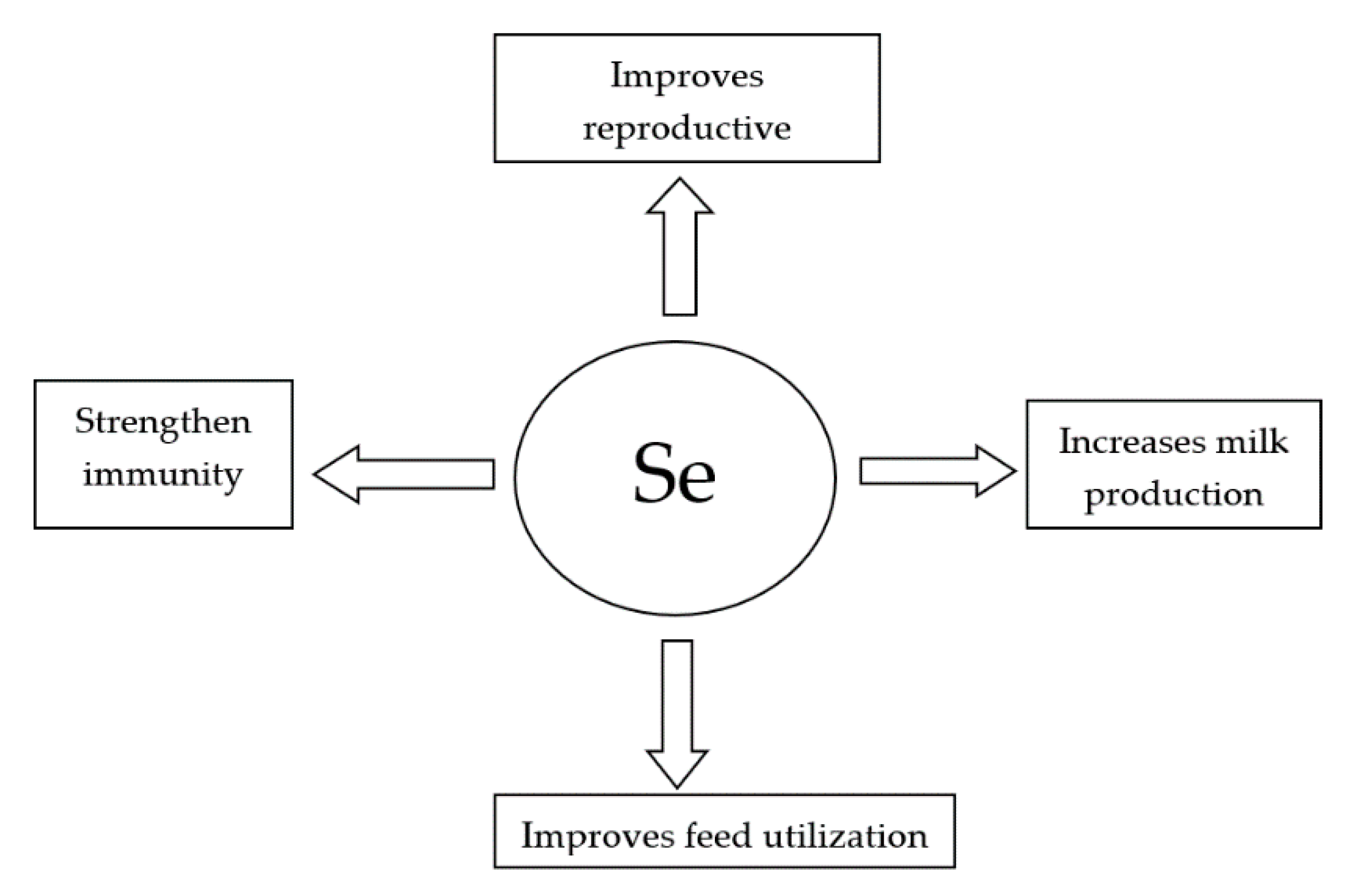
It is concluded from the present review that Se supplementation, especially in organic form, improves immunity, feed utilization, reproductive performance, and milk production in dairy cattle (Figure 1).
Lastly, excessive antioxidant supplementation generally can lead to an increased production of ROS [97,98]. Inorganic Se (selenite) is more toxic than the organic form. Inorganic Se reacts with glutathione, resulting in formation of selenotrisulphides which, in turn, react with other thiols and thus lead to the generation of oxygen free radicals, for example, superoxide anion. Organic Se is converted into selenols. This also results in the generation of oxygen free radicals inducing oxidative stress [88]. Mineral sulphates given in high doses can antagonize Se [94]. Deficiency of Se [99] is a much more common problem than its toxicity.
Table 1 summarizes the effects of the different forms of Se supplementation on productive and reproductive parameters, as well as the health status of dairy cows.
Author Contributions
All the authors equally contributed and commented on early and final versions of manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Authors extend thanks to their respected institutes and universities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Butterman, W.C.; Brown, R.D. Mineral Commodity Profiles—Selenium; U.S. USGS Numbered Series; USGS Publisher: Reston, Virginia, 2004; p. 20. [Google Scholar]
- Perrone, D.; Monteiro, M.; Nunes, J.C. The Chemistry of Selenium. R. Soc. Chem. 2015, 3–15. [Google Scholar] [CrossRef]
- Akl, M.A.; Ismael, D.S.; El-Asmy, A.A. Precipitate flotation-separation, speciation and hydride generation atomic absorption spectrometric determination of selenium (IV) in food stuffs. Microchem. J. 2006, 83, 61–69. [Google Scholar] [CrossRef]
- Shrivas, K.; Patel, D.K. Ultrasound assisted-hollow fibre liquid-phase microextraction for the determination of selenium in vegetable and fruit samples by using GF-AAS. Food Chem. 2011, 124, 1673–1677. [Google Scholar] [CrossRef]
- Fan, A.M.; Kizer, K.W. Selenium-nutritional, toxicologic, and clinical aspects. West. J. Med. 1990, 153, 160–167. [Google Scholar] [PubMed]
- Vonderheide, A.P.; Wrobel, K.; Kannamkumarath, S.S.; B’Hymer, C.; Montez-Bay’on, M.; Ponce de Leon, C.; Caruso, J.A. Characterization of selenium species in Brazil nuts by HPLC-ICP-MS and ES-MS. J. Agric. Food Chem. 2002, 50, 5722–5728. [Google Scholar] [CrossRef] [PubMed]
- Najafi, N.M.; Seidi, S.; Alizadeh, R.; Tavakoli, H. Inorganic selenium speciation in environmental samples using selective electrodeposition coupled with electrothermal atomic absorption spectrometry. Spectrochim. Acta B 2010, 65, 334–339. [Google Scholar] [CrossRef]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor-3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292. [Google Scholar] [CrossRef]
- Gergely, V.; Montes-Bayon, M.; Fodor, P.; Sanz-Medel, A. Selenium species in aqueous extracts of alfalfa sprouts by two-dimensional liquid chromatography coupled to inductively coupled plasma mass spectrometry and electrospray mass spectrometry detection. J. Agric. Food Chem. 2006, 54, 4524–4530. [Google Scholar] [CrossRef]
- Bodnar, M.; Konieczka, P.; Namieśnik, J. The Properties, Functions, and Use of Selenium Compounds in Living Organisms. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2012, 30, 225–252. [Google Scholar] [CrossRef]
- Puzanowska-Tarasiewicz, H.; Kuzmicka, L.; Tarasiewicz, M. Biological functions of selected elements and their compounds. 3, Zinc—An enzyme component and activator. Pol. Med. Merkur. Organ Pol. Med. Soc. 2009, 27, 419–422. [Google Scholar]
- Zarczy’nska, K.; Sobiech, P.; Radwi ´nska, J.; Rękawek, W. Effects of selenium on animal health. J. Elementol. 2013, 18, 329–340. [Google Scholar]
- Mehdi, Y.; Dufrasne, I. Selenium in Cattle: A Review. Molecules 2016, 21, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, V.S.; Bicalho, M.L.; Pereira, R.V.; Caixeta, L.S.; Knauer, W.A.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. Effect of an injectable trace mineral supplement containing selenium, copper, zinc, and manganese on the health and production of lactating Holstein cows. Vet. J. 2013, 197, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Alonso, M. Trace minerals and livestock: Not too much not too little. Int. Sch. Res. Netw. Vet. Sci. 2012, 704825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöne, F.; Steinhöfel, O.; Weigel, K.; Bergmann, H.; Herzog, E.; Dunkel, S.; Kirmse, R.; Leiterer, M. Selenium in feedstuffs and rations for dairy cows including a view of the food chain up to the consumer. J. Verbrauch. Lebensm. 2013, 8, 271–280. [Google Scholar]
- Lu, J.; Holmgren, A. Selenoproteins. J. Biol. Chem. 2009, 284, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Slavik, P.; Illek, J.; Brix, M.; Hlavicoca, J.; Rajmon, R.; Jilek, F. Influence of organic versus inorganic dietary selenium supplementation on the concentration of selenium in colostrum, milk and blood of beef cows. Acta Vet. Scand. 2008, 50, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.D.; Stewart, R.D. Metabolic studies of (75Se) selenomethionine and (75Se) selenite in the rat. Br. J. Nutr. 1973, 30, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Weiss, W.P. Selenium nutrition of dairy cows: Comparing responses to organic and inorganic selenium forms. In Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s; Lyons, P.T., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 12–14 May 2003; pp. 333–343. [Google Scholar]
- Tufarelli, V.; Ceci, E.; Laudadio, V. 2-Hydroxy-4-methylselenobutanoic acid as new organic selenium dietary supplement to produce selenium-enriched eggs. Biol. Trace Elem. Res. 2016, 171, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Podoll, K.L.; Bernard, J.B.; Ullrey, D.E.; DeBar, S.R.; Ku, P.K.; Magee, W.T. Dietary selenate versus selenite for cattle, sheep, and horses. J. Anim. Sci. 1992, 70, 1965–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, P.L.; Preston, R.L.; Pfander, W.H. Wholebody retention, tissue distribution and excretion of selenium after oral and intravenous administration in lambs fed varying selenium intakes. J. Nutr. 1969, 97, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbraith, M.L.; Vorachek, W.R.; Estill, C.T.; Whanger, P.D.; Bobe, G.; Davis, T.Z.; Hall, J.A. Rumen microorganisms decrease bioavailability of inorganic selenium supplements. Biol. Trace Elem. Res. 2015, 171, 338–343. [Google Scholar] [CrossRef]
- Schrauzer, G.N. Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 2000, 130, 1653–1656. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.T.; Ogra, Y. Metabolic pathway for selenium in the body: Speciation by HPLC-ICP MS with enriched Se. Food Addit. Contam. 2002, 19, 974–983. [Google Scholar] [CrossRef]
- Meschy, F. Nutrition Minérale des Ruminants; Editions Quae: Versaille, France, 2010; p. 208. [Google Scholar]
- Juniper, D.T.; Phipps, R.H.; Ramos-Morales, E.; Bertin, G. Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle. J. Anim. Sci. 2008, 86, 3100–3109. [Google Scholar] [CrossRef]
- Combs, G.F. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.G.; Herdt, T.H.; Stowe, H.D. Physical, hematologic, biochemical, and immunologic effects of supranutritional supplementation with dietary selenium in Holstein cows. Am. J. Vet. Res. 1997, 58, 760–764. [Google Scholar]
- Silvestre, F.T.; Rutigliano, H.M.; Thatcher, W.W.; Santos, J.E.P.; Staples, C.R. Florida Ruminant Nutrition Symposium; University of Florida: Gainesville, FL, USA, 2007; pp. 48–61. [Google Scholar]
- Lucia, F.; Pedrosa, C.; Motley, A.K.; Stevenson, T.D.; Hill, K.E. Fecal Selenium Excretion Is Regulated by Dietary Selenium Intake. Biol. Trace Elem. Res. 2012, 149, 377–381. [Google Scholar]
- Burk, R.F.; Hill, K.E. Regulation of Selenium Metabolism and Transport. Ann. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, L.L., Jr.; Pope, A.L.; Baumann, C.A. Distribution of microgram quantities of selenium in the tissues of the rat, effects of previous selenium intake. J. Nutr. 1966, 88, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Juniper, D.T.; Phipps, R.H.; Jones, A.K.; Bertin, G. Selenium Supplementation of Lactating Dairy Cows: Effect on Selenium Concentration in Blood, Milk, Urine, and Feces. J. Dairy Sci. 2006, 89, 3544–3551. [Google Scholar] [CrossRef]
- Hrbac, J.; Kohen, R. Biological redox activity: Its importance, methods for its quantification and implication for health and disease. Drug Dev. Res. 2000, 50, 516–527. [Google Scholar] [CrossRef]
- Yadav, A.; Kumari, R.; Yadav, A.; Mishra, J.P.; Srivatva, S.; Prabha, S. Antioxidants and its functions in human body—A Review. Res. Environ. Life Sci. 2016, 9, 1328–1331. [Google Scholar]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of Reactive Oxygen Species in Cell Signaling Pathways. Biochem. Biomed. Asp. Oxidative Modif. 2001, 29, 345–350. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Adwas, A.A.; Elsayed, A.; Azab, A.E.; Quwaydir, F.A. Oxidative stress and antioxidant mechanisms in human body. J. Appl. Biotechnol. 2019, 6, 43–47. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Juniper, D.T. Revisiting Oxidative Stress and the Use of Organic Selenium in Dairy Cow Nutrition. Animals 2019, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Selenium in Poultry Nutrition and Health; Academic Publishers: Wageningen, The Netherlands, 2018. [Google Scholar]
- Boxin, O.U.; Dejian, H.; Maureen, A.F.; Elizabeth, K.D. Analysis of antioxidant activities of common vegetables employing oxygen radical Absorbance Capacity (ORAC) and Ferric Reducing Antioxidant Power (FRAP) Assays: A comparative study. J. Agric. Food Chem. 2002, 5, 223–228. [Google Scholar]
- Burk, R.F.; Hill, K.E.; Motley, A.K. Selenoprotein Metabolism and Function: Evidence for More than One Function for Selenoprotein. Pak. J. Nutr. 2003, 133, 1517S–1520S. [Google Scholar] [CrossRef] [PubMed]
- Mezzetti, A.; Ilio, C.D.; Calafiore, A.M.; Aceto, A.; Marzio, L.; Frederici, G.; Cuccurullo, F. Glutathione peroxidase, glutathione reductase and glutathione transferase activities in the human artery, vein and heart. J. Mol. Cardiol. 1990, 22, 935–938. [Google Scholar] [CrossRef]
- Grasso, P.J.; Scholz, R.W.; Erskine, R.J.; Eberhart, R.J. Phagocytosis, bactericidal activity, and oxidative metabolism of mammary neutrophils from dairy cows fed selenium-adequate and selenium-deficient diets. Am. J. Vet. Res. 1990, 51, 269–274. [Google Scholar] [PubMed]
- Gunter, S.A.; Beck, P.A.; Phillips, J.M. Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 2003, 81, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.R.; Boyne, R. Superoxide dismutase and glutathione peroxidase activities in neutrophils from selenium deficient and copper deficient cattle. Life Sci. 1985, 36, 1569–1575. [Google Scholar] [CrossRef]
- Awadeh, F.T.; Kincaid, R.L.; Johnson, K.A. Effect of level and sources of dietary selenium on concentrations of thyroid hormones and immunoglobulins in beef cows and calves. J. Anim. Sci. 1998, 76, 1204–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, W.R. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 2000, 60–61, 449–457. [Google Scholar] [CrossRef]
- Rabiee, A.R.; Lean, I.J.; Stevenson, M.A.; Socha, M.T. Effects of feeding organic trace minerals on milk production and reproductive performance in lactating dairy cows: A meta-analysis. J. Dairy Sci. 2010, 93, 4239–4251. [Google Scholar] [CrossRef] [Green Version]
- Oltramari, C.E.; Pinheiro, M.G.; Miranda, M.S.; Arcaro, J.R.P.; Castelani, L.; Toledo, L.C.; Ambrósio, L.A.; Leme, P.R.; Manella, M.Q.; Júnior, I.A. Selenium Sources in the Diet of Dairy Cows and Their Effects on Milk Production and Quality, on Udder Health and on Physiological Indicators of Heat Stress. Ital. J. Anim. Sci. 2014, 13, 2921. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.B. Serum selenium concentrations and glutathione peroxidase activities in cattle grazing forages of various selenium concentrations. Am. J. Vet. Res. 1985, 46, 1556–1561. [Google Scholar]
- Tufarelli, V.; Laudadio, V. Dietary supplementation with selenium and vitamin E improves milk yield, composition and rheological properties of dairy Jonica goats. J. Dairy Res. 2011, 78, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Salman, S.; Khol-Parisini, A.; Schafft, H.; Lahrssen-Wiederholt, M.; Hulan, H.W.; Dinse, D.; Zentek, J. The role of dietary selenium in bovine mammary gland health and immune function. Anim. Health Res. Rev. 2009, 10, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Souza, F.N.; Blagitz, M.G.; Latorre, A.O.; Mori, C.S.; Sucupira, M.C.A.; Libera, A.M.M.P.D. Effects of in vitro selenium supplementation on blood and milk neutrophils from dairy cows. Pesqui. Vet. Bras. 2012, 32, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, H.H.; Safwat, M.A.; El-Sysy, M.A.I.; Al-Metwaly, M.A. Effect of selenium and vitamin E supplementation as a nutritional treatment for some physiological and productive traits of Holstein dairy cows under Egyptian summer conditions. J. Egypt. Acad. Soc. Environ. Dev. 2016, 17, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Cordova-Izquierdo, C.G.R.L.; Campos, V.M.X.; Suarez, S.C.; Cordova-Jimenez, C.A.; Cordova-Jimenez, M.S. Effect of gonadotophin releasing factor and antioxidants on the rate of estrous repetition dairy cows. Aust. J. Basic Appl. Sci. 2010, 408, 1282–1287. [Google Scholar]
- Wang, C.; Liu, Q.; Yang, W.Z.; Dong, Q.; Yang, X.M.; He, D.C.; Huang, Y.X. Effects of selenium yeast on rumen fermentation, lactation performance and feed digestibilities in lactating dairy cows. Livest. Sci. 2009, 126, 239–244. [Google Scholar] [CrossRef]
- Wilde, D. Influence of macro and micro minerals in the peri-parturient period on fertility in dairy cattle. Anim. Reprod. Sci. 2006, 96, 240–249. [Google Scholar] [CrossRef]
- Ullah, H.; Khan, R.U.; Mobashar, M.; Ahmad, S.; Sajid, A.; Khan, N.U.; Usman, T.; Khattak, I.; Hamayun, K. Effect of yeast-based selenium on blood progesterone, metabolites and milk yield in Achai dairy cows. Ital. J. Anim. Sci. 2019, 18, 1445–1450. [Google Scholar] [CrossRef] [Green Version]
- Eulogio, G.L.J.; Alberto, S.O.J.; Hugo, C.V.; Antonio, C.N.; Alejandro, C.; Juan, M.Q. Effects of Selenium and vitamin E in the production, physiochemical composition and somatic cell count in milk of Ayrshire cows. J. Anim. Vet. Adv. 2012, 11, 687–691. [Google Scholar]
- Chauhana, S.S.; Ponnampalama, E.N.; Celi, P.; Hopkins, D.L.; Leurya, B.J.; Dunsheaa, F.R. High dietary vitamin E and selenium improves feed intake and weight gain of finisher lambs and maintains redox homeostasis under hot conditions. Small Rumin. Res. 2016, 137, 17–23. [Google Scholar] [CrossRef]
- Smith, J.F.; Brouk, M.J. Factors affecting dry matter intake by lactating dairy cows. Kans. Agric. Exp. Stn. Res. Rep. 2000, 2, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Alhidary, I.; Shini, S.; Al Jassim, R.; Gaughan, J. Effect of various doses of injected selenium on performance and physiological responses of sheep to heat load. J. Anim. Sci. 2012, 90, 2988–2994. [Google Scholar] [CrossRef] [PubMed]
- Ganie, A.A.; Baghel, R.P.S.; Mudgal, V.; Sheikh, G.G. Effect of Selenium Supplementation on Growth and Nutrient Utilization in Buffalo Heifers. Anim. Nutr. Feed Technol. 2010, 10, 255–259. [Google Scholar]
- Surai, P.F. Selenium in poultry nutrition-1. Antioxidant properties, deficiency and toxicity. World Poult. Sci. J. 2002, 58, 333–347. [Google Scholar] [CrossRef]
- Willshire, J.A.; Payne, J.H. Selenium and vitamin E in dairy cows—A review. Cattle Pract. 2011, 19, 22–30. [Google Scholar]
- Rooke, J.A.; Robinson, J.J.; Arthur, J.R. Effects of vitamin E and selenium on the performance and immune status of ewes and lambs. J. Agric. Sci. 2004, 142, 253–262. [Google Scholar] [CrossRef]
- Gartner, R.; Gasnier, B.; Dietrich, J.; Krebs, B.; Angstwurm, M. Selenium supplementation in patients with autoimmune thyroiditis decreases thyroid peroxidase antibodies concentrations. J. Clin. Endocrinol. Metab. 2007, 87, 1687–1691. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L.; Riese, C.; Michaelis, M.; Griebert, E.; Klein, M.; Sapin, R.; Schweizer, U.; Köhrle, J. Synthesis and metabolism of thyroid hormones is preferentially maintained in seleniumdeficient transgenic mice. Endocrinology 2007, 147, 1306–1313. [Google Scholar] [CrossRef] [Green Version]
- Taheri, Z.; Karimi, S.; Mehrban, H.; Moharrery, A. Supplementation of different selenium sources during early lactation of native goats and their effects on nutrient digestibility, nitrogen and energy status. J. Appl. Anim. Res. 2018, 46, 64–68. [Google Scholar] [CrossRef]
- Weiss, W.P. Selenium sources for dairy cattle. In Proceedings of the Tri–State Dairy Nutrition Conference, Fort Wayne, IN, USA, 2–3 May 2005. [Google Scholar]
- Alimohamady, R.; Aliarabi, H.; Bahari, A.; Dezfoulian, A.H. Influence of Different Amounts and Sources of Selenium Supplementation on Performance, Some Blood Parameters, and Nutrient Digestibility in Lambs. Biol. Trace Elem. Res. 2013, 154, 45–54. [Google Scholar] [CrossRef]
- Do, C.; Wasana, N.; Cho, K.; Choi, Y.; Choi, T.; Park, B.; Lee, D. The Effect of Age at First Calving and Calving Interval on Productive Life and Lifetime Profit in Korean Holsteins. Asian Australas. J. Anim. Sci. 2013, 26, 1511–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedaoo, M.; Khllare, K.; Meshram, M.; Sahatpure, S.; Patil, M. Study of some serum trace minerals in cyclic and noncyclic surti buffaloes. Vet. World 2008, 1, 71–72. [Google Scholar]
- Kamada, H. Effects of selenium-rich yeast supplementation on the plasma progesterone levels of postpartum dairy cows. Asian Australas. J. Anim. Sci. 2017, 30, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pursley, J.R.; Martins, J.P. Impact of circulating concentrations of progesterone and antral age of the ovulatory follicle on fertility of high-producing lactating dairy cows. Reprod. Fertil. Dev. 2011, 24, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inskeep, E.K. Preovulatory, and postmaternal recognition effects of concentrations of progesterone on embryonic survival in the cow. J. Anim. Sci. 2004, 82, 24–39. [Google Scholar] [CrossRef]
- Kamada, H.; Ikumo, H. Effect of selenium on cultured bovine luteal cells. Anim. Reprod. Sci. 1997, 46, 203–211. [Google Scholar] [CrossRef]
- Bajaj, N.K.; Sharma, N. Endocrine Causes of Early Embryonic Death: An Overview. Curr. Res. Dairy Sci. 2011, 3, 1–24. [Google Scholar] [CrossRef]
- Ganie, A.A.; Baghel, R.P.S.; Mudgal, V.; Aarif, O.; Sheikh, G.G. Effect of selenium supplementation on reproductive performance and hormonal profile in buffalo heifers. Ind. J. Anim. Res. 2014, 48, 27–30. [Google Scholar] [CrossRef]
- Thatcher, W.; Santos, J.E.P.; Staples, C.R. Dietary manipulations to improve embryonic survival in cattle. Theriogenology 2011, 76, 1619–1631. [Google Scholar] [CrossRef]
- Dimri, U.; Ranjan, R.; Sharma, M.C.; Varshney, V.P. Effect of vitamin E and selenium supplementation on oxidative stress indices and cortisol level in blood in water buffaloes during pregnancy and early postpartum period. Trop. Anim. Health Prod. 2010, 42, 405–410. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, R.; Malevu, T.D.; Sochor, J.; Baron, M.; Melcova, M.; Zidkova, J.; et al. A Summary of New Findings on the Biological Effects of Selenium in Selected Animal Species—A Critical Review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef] [PubMed]
- Mézes, M.; Balogh, K. Prooxidant mechanisms of selenium toxicity—A review. Acta Biol. Szeged. 2009, 53, 15–18. [Google Scholar]
- Hartikainen, H. Biogeochemistry of selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Biogeochemia Pierwiastk’ow‘sladowych; PWN Press: Warsaw, Poland, 1999. [Google Scholar]
- Mihajlović, M. Toksicnost selena kod domaćih zivotinja [Selenium toxicity in domestic animals]. Glas Srp Akad Nauka Med. 1992, 42, 131–144. [Google Scholar]
- Neve, J.; Favier, A. Selenium in medecine and biology. In Proceedings of the Second International Congress on Trace Elements in Medecine and Biology, Avoriaz, France, March 1988. [Google Scholar]
- Claude, J.B. Introduction à la Nutrition des Animaux Domestiques; Tec & Doc/EM Inter: Paris, France, 2002. [Google Scholar]
- Rogers, P.A.M.; Arora, S.P.; Fleming, G.A.; Crinion, R.A.P.; McLaughlin, J.G. Selenium toxicity in farm animals: Treatment and prevention. Ir. Vet. J. 1990, 43, 151–153. [Google Scholar]
- James, L.F.; James, L. Shupe. Selenium Poisoning in Livestock. Rangelands 1984, 6, 64–67. [Google Scholar]
- Nair, M.P.N.; Schwartz, S.A. Immunoregulation of natural and lymphokine-activated killer cells by selenium. Immunopharmacology 1990, 19, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Abuelo, A.; Alves-Nores, V.; Hernandez, J.; Muiño, R.; Benedito, J.L.; Castillo, C. Effect of parenteral antioxidant supplementation during the dry period on postpartum glucose tolerance in dairy cows. J. Vet. Intern. Med. 2016, 30, 892–898. [Google Scholar]
- Rizzo, A.; Pantaleo, M.; Mutinati, M.; Minoia, G.; Trisolini, C.; Ceci, E.; Sciorsci, R.L. Blood and milk oxidative status after administration of different antioxidants during early postpartum in dairy cows. Res. Vet. Sci. 2013, 95, 1171–1174. [Google Scholar] [CrossRef]
- Szpunar, J.; Lobinski, R.; Prange, A. Hyphenated techniques for elemental speciation in biological systems. Appl. Spectrosc. 2003, 57, 102–112. [Google Scholar] [CrossRef]
- Najafnejad, B.; Aliarabi, H.; Tabatabaei, M.M.; Alimohamady, R. Effects of different forms of Selenium supplementation on production performance and nutrient digestibility in lactating dairy cattle. In Proceedings of the Second International Conference on Agriculture and Natural Resources, Kermanshah, Iran, 25–26 December 2013. [Google Scholar]
- Kamada, H.; Hondate, K. Effect of dietary Selenium supplementation on plasma progesterone concentration in cows. J. Vet. Med. Sci. 1998, 60, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Bobe, G.; Vorachek, W.R.; Kasper, K.; Traber, M.G.; Mosher, W.D.; Pirelli, G.J.; Gamroth, M. Effect of supranutritional organic selenium supplementation on postpartum blood micronutrients, antioxidants, metabolites, and inflammation biomarkers in selenium-replete dairy cows. Biol. Trace Elem. Res. 2014, 161, 272–287. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Benefits of Se supplementation in dairy cattle nutrition.


{kind=link}
Table 1.
Effect of Se supplementation on different parameters of dairy cows. DM: Dry matter; OM: Organic matter; NDF: Neutral detergent fiber; NFC: Non-fiber carbohydrates.
Table 1.
Effect of Se supplementation on different parameters of dairy cows. DM: Dry matter; OM: Organic matter; NDF: Neutral detergent fiber; NFC: Non-fiber carbohydrates.
| Parameter | Form of Se | Dose of Se | Effect | Reference |
|---|---|---|---|---|
| Milk production | Organic | 0.15 mg/kg DMI | Increased | [61] |
| Organic | 0.3 mg/kg DMI | Increased | [61,63,64] | |
| Organic | 0.4 mg/kg DMI | Decreased | [63] | |
| Neutrophil function | Inorganic/sodium selenite | 10 μM | Improved | [58] |
| Organic | 0.3 mg/kg DMI | Increased | [33] | |
| Adaptive immunity | Organic | 0.3 mg/kg DMI | Improved | [33] |
| Somatic cell count | Organic | 0.3 mg/kg DMI | Decreased | [33] |
| Milk fat | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Organic | 0.3 mg/kg DMI | Increased | [33] | |
| Milk protein | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Digestibility (DM, OM, NDF, Ether extract, NFC) | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Plasma P4 | Inorganic | 0.5 mg/kg DMI | Increased | [101] |
| Serum Se | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Plasma Se | Organic | 0.3 mg/kg DMI | Increased | [33] |
| Whole blood Se concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum amyloid A | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Erythrocyte glutathione | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum albumin concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum cholesterol concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Decreased | [102] |
| α-tocopherol/cholesterol ratio | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Uterine health | Organic | 0.3 mg/kg DMI | Improved | [33] |
| Second service pregnancy rate | Organic | 0.3 mg/kg DMI | increased | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ullah, H.; Khan, R.U.; Tufarelli, V.; Laudadio, V. Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production. Sustainability 2020, 12, 10693. https://0-doi-org.brum.beds.ac.uk/10.3390/su122410693
AMA Style
Ullah H, Khan RU, Tufarelli V, Laudadio V. Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production. Sustainability. 2020; 12(24):10693. https://0-doi-org.brum.beds.ac.uk/10.3390/su122410693
Chicago/Turabian StyleUllah, Hammad, Rifat Ullah Khan, Vincenzo Tufarelli, and Vito Laudadio. 2020. "Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production" Sustainability 12, no. 24: 10693. https://0-doi-org.brum.beds.ac.uk/10.3390/su122410693
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.