Potential Distribution of the Critically Endangered Chinese Pangolin (Manis pentadactyla) in Different Land Covers of Nepal: Implications for Conservation

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
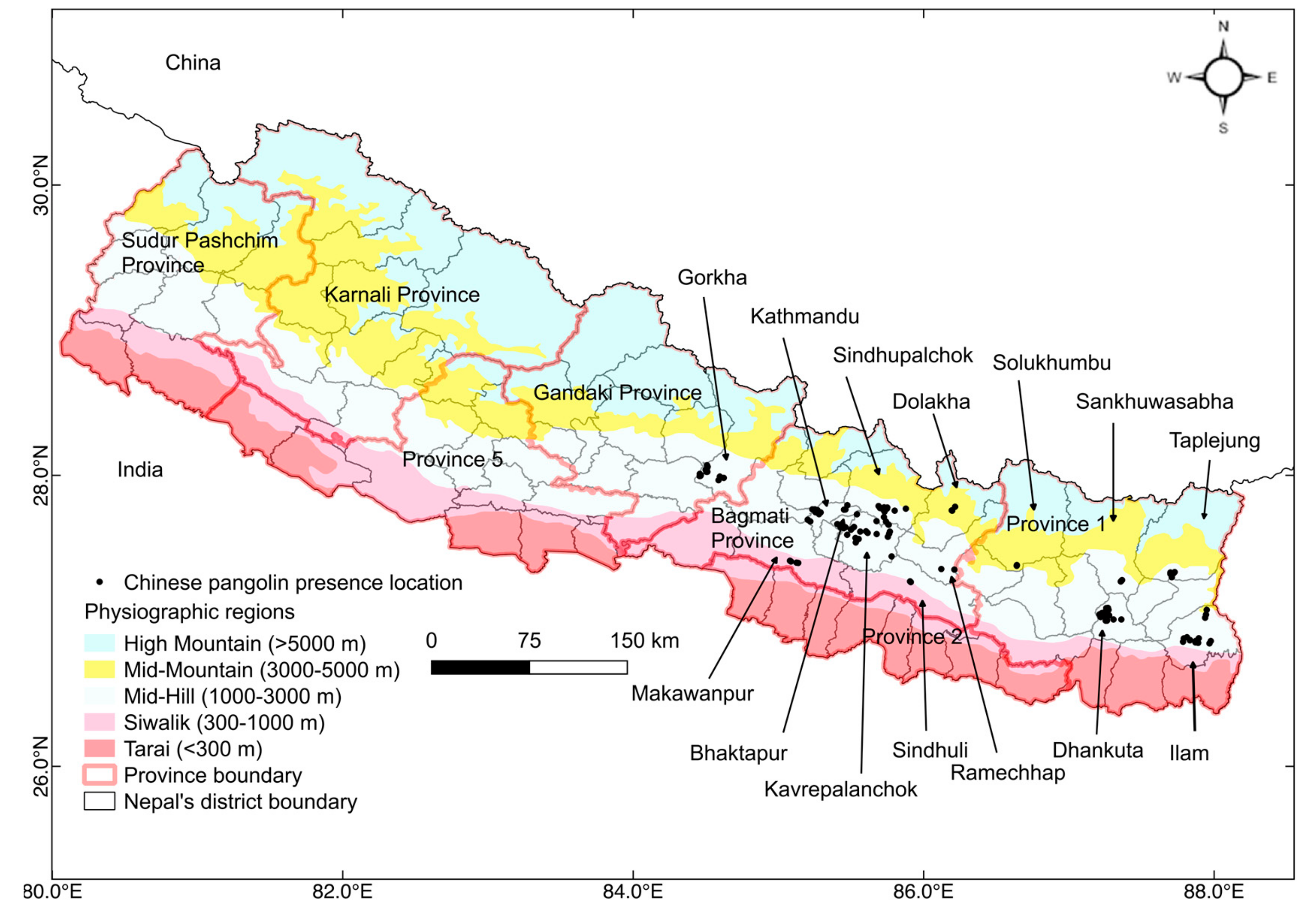
2.1. Study Area
2.2. Data Collection and Management
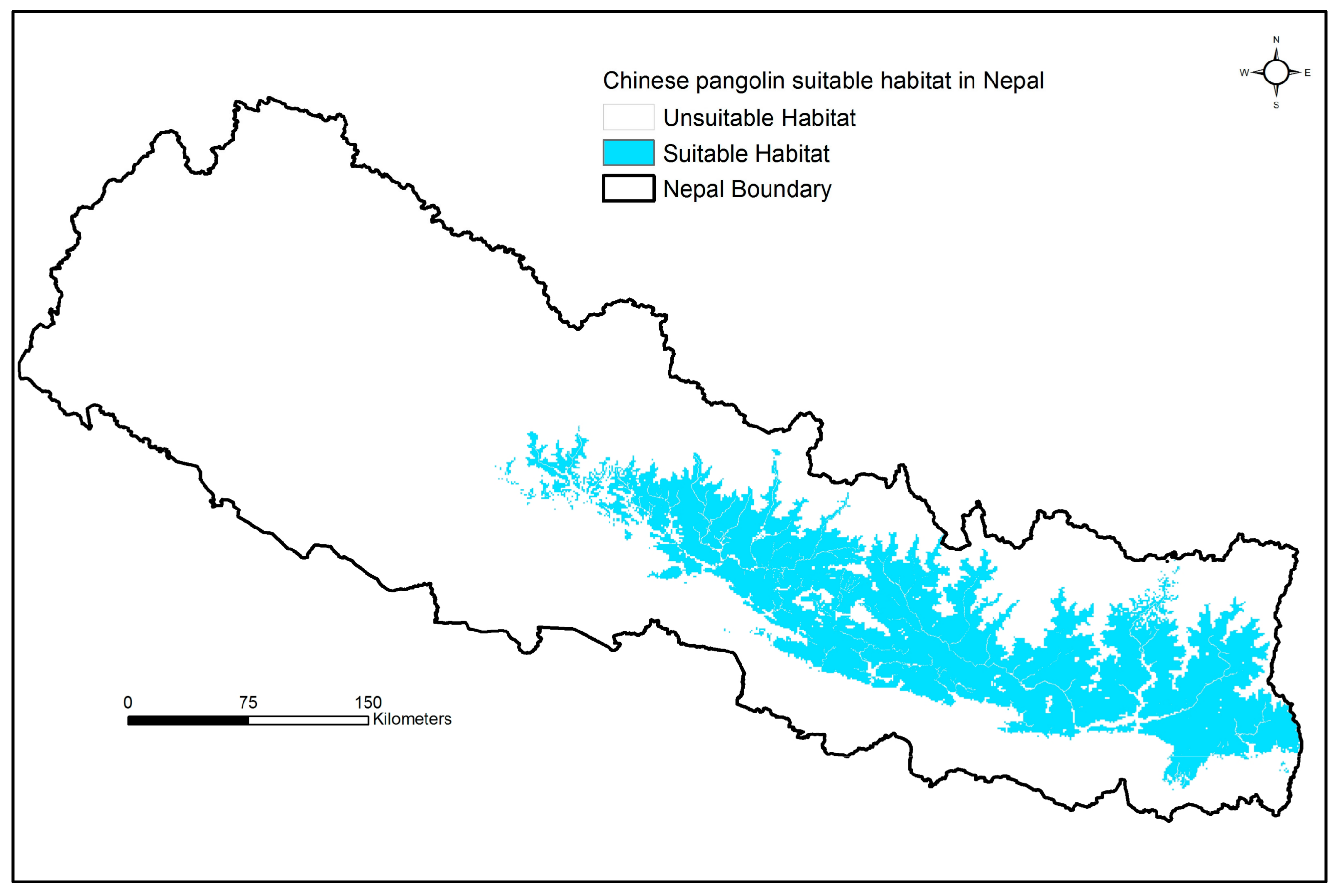
2.3. Spatial Modeling
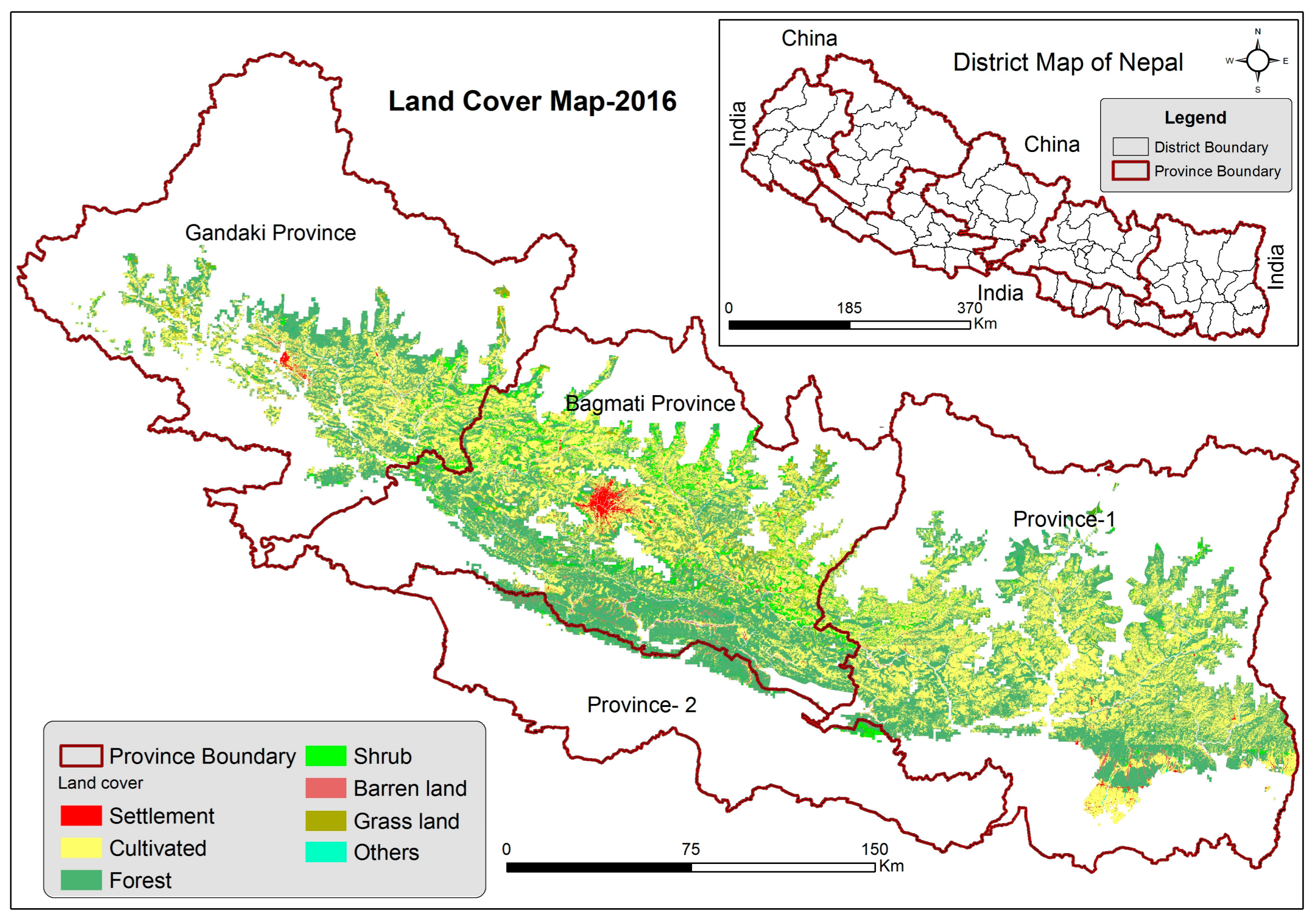
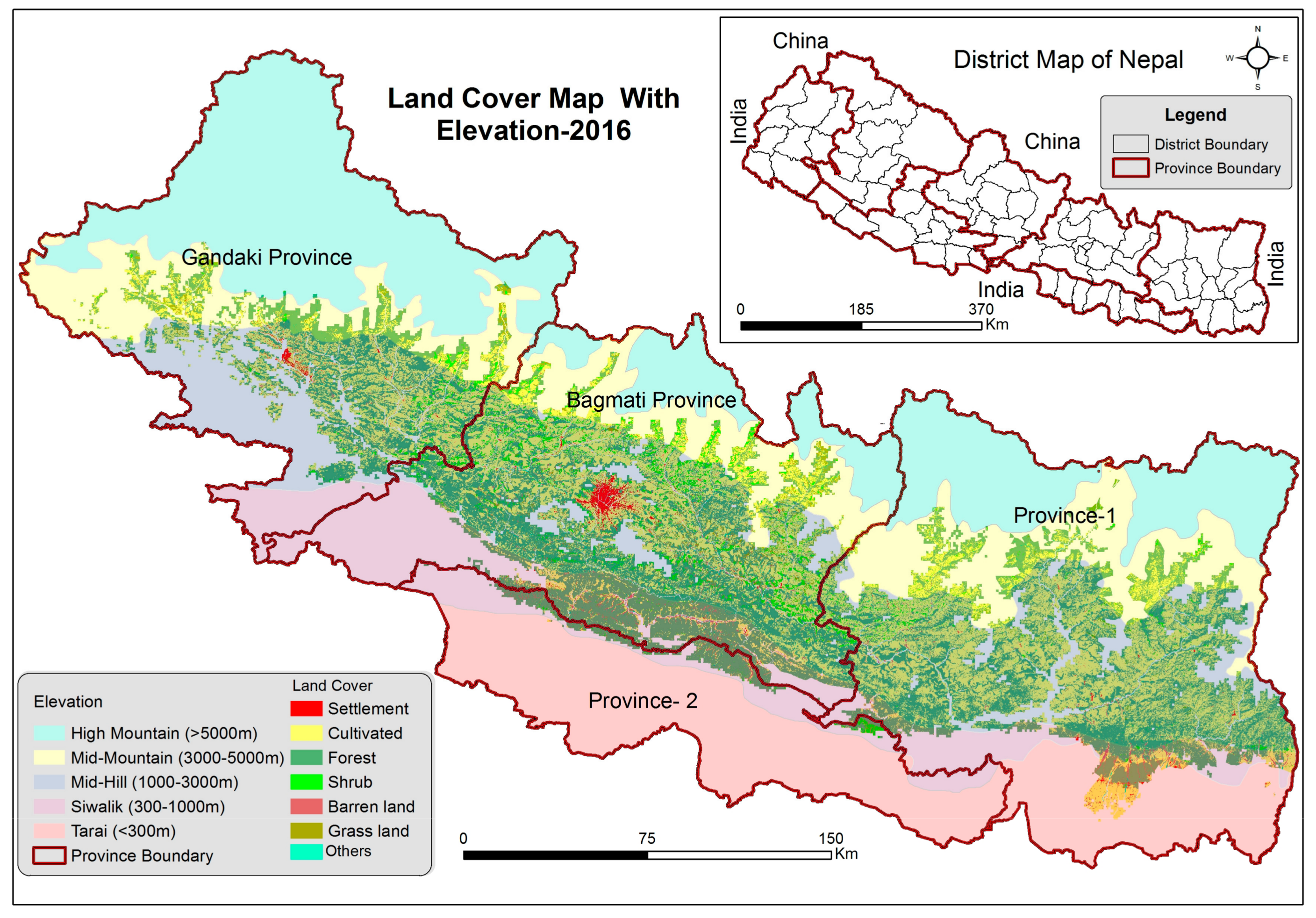
2.4. Land Cover Classification
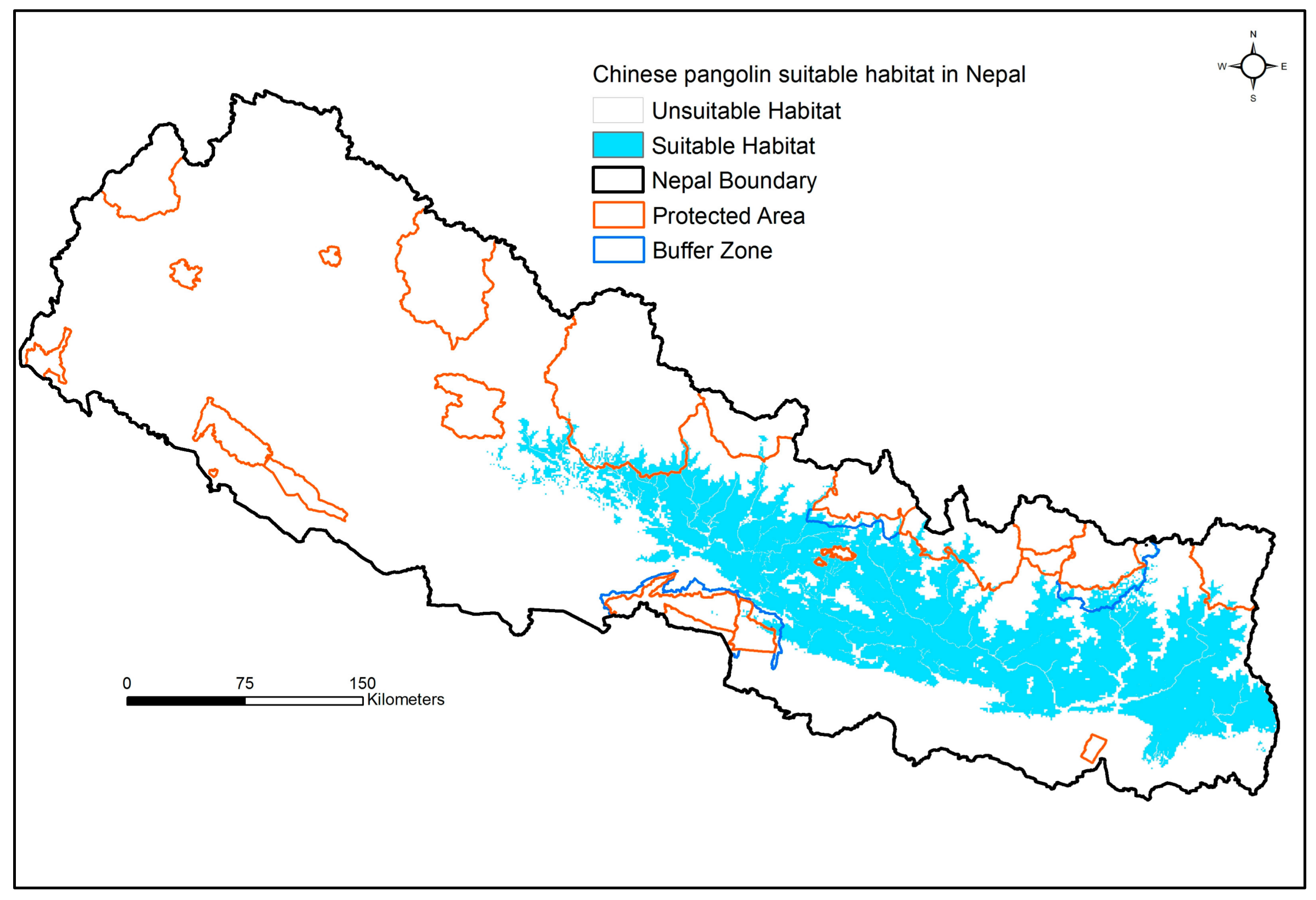
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Owens, I.P.F.; Bennett, P.M. Ecological basis of extinction risk in birds: Habitat loss versus human persecution and introduced predators. Proc. Natl. Acad. Sci. 2000, 97, 12144–12148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [PubMed]
- Sharma, H.P.; Belant, J.L.; Swenson, J.E. Effects of livestock on occurrence of the vulnerable red panda Ailurus fulgens in Rara National Park, Nepal. Oryx 2014, 48, 228–231. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: entering the sixth mass extinction. Sci. Adv. 2015, 1, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WWF [World Wildlife Fund]. In Living Planet Report 2016. Risk and Resilience in a New Era; WWF International: Gland, Switzerland, 2016.
- Rodrigues, A.S.L.; Brooks, T.M.; Butchart, S.H.M.; Chanson, J.; Cox, N.; Hoffmann, M.; Stuart, S.N. Spatially explicit trends in the global conservation status of vertebrates. PLoS ONE 2014, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sands, J.P.; DeMaso, S.J.; Schnupp, M.J.; Brennan, L.A. Wildlife Science: Connecting Research with Management. CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Greggor, A.L.; Berger-Tal, O.; Blumstein, D.T.; Angeloni, L.; Bessa-Gomes, C.; Blackwell, B.F.; St Clair, C.C.; Crooks, K.; de Silva, S.; Fernández-Juricic, E.; et al. Research Priorities from Animal Behaviour for Maximising Conservation Progress. Trends Ecol. Evol. 2016, 31, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Gavashelishvili, A.; Lukarevskiy, V. Modelling the habitat requirements of leopard Panthera pardus in west and central Asia. J. Appl. Ecol. 2008, 45, 579–588. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Larson, M.A.; Thompson, F.R.; Millspaugh, J.J.; Dijak, W.D.; Shifley, S.R. Linking population viability, habitat suitability, and landscape simulation models for conservation planning. Ecol. Modell. 2004, 180, 103–118. [Google Scholar] [CrossRef]
- Martínez-Meyer, E.; Peterson, A.T. Conservatism of ecological niche characteristics in North American plant species over the Pleistocene-to-Recent transition. J. Biogeogr. 2006, 33, 1779–1789. [Google Scholar] [CrossRef]
- Austin, M.P.; Meyers, J.A. Current approaches to modelling the environmental niche of eucalypts: Implication for management of forest biodiversity. For. Ecol. Manage. 1996, 85, 95–106. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Srinivasulu, B. Checklist of scandents and pholidots (Mammalia: Scandentia and Pholidota) of South Asia. Zoos’ Print J. 2013, 19, 1372–1374. [Google Scholar] [CrossRef]
- Challender, D.W.S.; Baillie, J.; Ades, G.; Kaspal, P.; Chan, B.; Khatiwada, A.; Xu, L.; Chin, S.; KC, R.; Nash, H.; et al. Manis pentadactyla. The IUCN Red List of Threatened Species: e.T12764A45222544. 2014. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2014-2.RLTS.T12764A45222544.en (accessed on 25 October 2019).
- Challender, D.W.S.; Waterman, C.; Baillie, J.E.M. Scaling up pangolin conservationIUCN SSC Pangolin Specialist Group Conservation Action Plan. 24 July 2014. Available online: https://www.iucn.org/downloads/scaling_up_pangolin_conservation_280714_v4_1.pdf (accessed on 25 October 2019).
- Jnawali, S.R.; Baral, H.S.; Lee, S.; Subedi, N.; Acharya, K.P.; Upadhyay, G.P.; Pandey, M.; Shrestha, R.; Joshi, D.; Lamichhane, B.R.; et al. The Status of Nepal’s Mammals: the National Red List Series. Department of National Parks and Wildlife Conservation: Kathmandu, Nepal, 2011. [Google Scholar]
- Corlett, R.T. The Impact of Hunting on the Mammalian Fauna of Tropical Asian Forests. Biotropica 2007, 39, 292–303. [Google Scholar] [CrossRef]
- Challender, D.; Waterman, C. Implementation of CITES Decision2 17.239 b) and 17.240 on Pangolins (Manis spp.). Prepared by IUCN for the CITES SecretariatSC69 Doc, 57. 2017. Available online: https://cites.org/sites/default/files/eng/com/sc/69/E-SC69-57-A.pdf (accessed on 26 October 2019).
- Heinrich, S.; Wittman, T.A.; Ross, J.V.; Shepherd, C.R.; Challender, D.W.S.; Cassey, P. The Global Trafficking of Pangolins: A comprehensive summary of seizures and trafficking routes from 2010–2015. TRAFFIC. Southeast Asia Regional Office: Petaling Jaya, Selangor, Malaysia, 2017. [Google Scholar]
- Acharya, S.; Rayamajhi, S.; Sharma, S.; Upadhaya, S.; Joshi, S.; Lamichhane, S. Anthropogenic threats to survival of the critically endangered Chinese pangolins (Manis pentadactyla) and their habitat in Kavrepalanchowk Nepal. J. Biodiver. Endanger. Species 2018, 6, 218. [Google Scholar]
- Katuwal, H.B.; Neupane, K.R.; Adhikari, D.; Sharma, M.; Thapa, S. Pangolins in eastern Nepal: trade and ethno-medicinal importance. J. Threat. Taxa 2015, 7, 7563–7567. [Google Scholar] [CrossRef]
- Kaspal, P.; Shah, K.B.; Baral, H.S. Saalak (i.e., Pangolin); Himalayan Nature: Kathmandu, Nepal, 2016. [Google Scholar]
- Mohapatra, R.K.; Panda, S.; Acharjyo, L.N.; Nair, M.V.; Challender, D.W. A note on the illegal trade and use of pangolin body parts in India. Traffic Bullet. 2015, 27, 33–40. [Google Scholar]
- Newton, P.; Van Thai, N.; Roberton, S.; Bell, D. Pangolins in peril: Using local hunters’ knowledge to conserve elusive species in Vietnam. Endanger. Species Res. 2008, 6, 41–53. [Google Scholar] [CrossRef]
- Gurung, J.B. A pangolin survey in Royal Nagarjung Forest in Kathmandu, Nepal. Tiger Paper 1996, 23, 29–32. [Google Scholar]
- Bhandari, N.; Chalise, M.K. Habitat and distribution of Chinese pangolin (Manis pentadactyla Linnaeus, 1758) in Nagarjun Forest of Shivapuri-Nagarjun National Park, Nepal. Nepalese J. Zool. 2014, 2, 18–25. [Google Scholar]
- Katuwal, H.B.; Sharma, H.P.; Parajuli, K. Anthropogenic impacts on the occurrence of the critically endangered Chinese pangolin (Manis pentadactyla) in Nepal. J. Mammal. 2017, 98, 1667–1673. [Google Scholar] [CrossRef]
- Paudel, B.; Zhang, Y.l.; Li, S.-C.; Liu, L.-S.; Wu, X.; Khanal, N.R. Review of studies on land use and land cover change in Nepal. J. Mt. Sci. 2016, 13, 643–660. [Google Scholar] [CrossRef]
- DNPWC; DoF. Pangolin Conservation Action Plan for Nepal (2018-2022); Department of National Parks and Wildlife Conservation and Department of Forests: Kathmandu, Nepal, 2018. [Google Scholar]
- Benítez-López, A.; Alkemade, R.; Verweij, P.A. The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol. Conserv. 2010, 143, 1307–1316. [Google Scholar] [CrossRef] [Green Version]
- Canova, L.; Balestrieri, A. Long-term monitoring by roadkill counts of mammal populations living in intensively cultivated landscapes. Biodivers. Conserv. 2019, 28, 97–113. [Google Scholar] [CrossRef]
- Schtickzelle, N.; Baguette, M. Behavioural responses to habitat patch boundaries restrict dispersal and generate emigration - patch area relationships in fragmented landscapes. J. Anim. Ecol. 2003, 72, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 6, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the 21st International Conference on Machine Learning; ACM Press: New York, NY, USA, 2004; p. 655. Available online: https://www.cs.princeton.edu/~schapire/papers/maxent_icml.pdf (accessed on 26 October 2019).
- Philipps, S.J.; Dudı´k, M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Wilson, K.A.; Carwardine, J.; Possingham, H.P. Setting conservation priorities. Ann. N. Y. Acad. Sci. 2009, 1162, 237–264. [Google Scholar] [CrossRef]
- LRMP. Land Resources Mapping Project. Kathmandu, Nepal: Survey Department, His majesty Government of Nepal and Kenting Earth Sciences; Government of Nepal and Government of Canada, Land Resources Mapping Project: Kathmandu, Nepal, 1986.
- Baral, H.S.; Shah, K.B. Wild Mammals of Nepal, Himalayan Nature: Kathmandu, Nepal, 2008.
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological niches and geographic distributions (MPB-49). Princeton University Press: Princeton, NJ, USA, 2011; Volume 56. [Google Scholar]
- Phillips, S.J.; Dudik, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Receiver operating characteristic (ROC) curve analysis for medical diagnostic test evaluation. Caspian. J. Intern. Med. 2013, 4, 627–635. [Google Scholar]
- Lobo, J.M.; Jiménez-valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- R Development Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2017. [Google Scholar]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Rimal, B.; Zhang, L.; Keshtkar, H.; Haack, B.; Rijal, S.; Zhang, P. Land use/land cover dynamics and modeling of urban land expansion by the integration of cellular automata and markov chain. ISPRS Int. J. Geo-Inf. 2018, 7, 154. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A. Monitoring land cover change in urban and peri-urban areas using dense time stacks of Landsat satellite data and a data mining approach. Remote Sens. Environ. 2012, 124, 689–704. [Google Scholar] [CrossRef]
- Kavzoglu, T.; Colkesen, I. A kernel functions analysis for support vector machines for land cover classification. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 352–359. [Google Scholar] [CrossRef]
- Sothe, C.; de Almeida, C.M.; Liesenberg, V.; Schimalski, M.B. Evaluating Sentinel-2 and Landsat-8 data to map sucessional forest stages in a subtropical forest in Southern Brazil. Remote Sens. 2017, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Thenkabail, P.S.; Tilton, J.C.; Gumma, M.K.; Teluguntla, P.; Id, A.O.; Id, R.G.C.; Id, K.Y.; Gorelick, N. Nominal 30-m cropland extent map of continental Africa by integrating pixel-based and object-based algorithms using sentinel-2 and landsat-8 data on Google Earth Engine. Remote Sens. 2017, 9, 1065. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.R.; Hardy, E.E.; Roach, J.T.; Witmer, R.E. A Land Use and Land Cover Classification System for Use with Remote Sensor Data; U.S. Geological Survey, 604 South Pickett Street, Alexandria, VA 22304; 1976. Available online: https://pubs.usgs.gov/pp/0964/report.pdf (accessed on 26 October 2019).
- Campbell, J.B.; Wynne, R.H. Introduction to Remote Sensing, 5th ed.Guilford Press: New York, NY, USA, 2011. [Google Scholar]
- GoN. Topographical Map. Min Bhawan: Kathmandu, Nepal, 1995. [Google Scholar]
- Yang, C.W.; Guo, J.C.; Li, Z.W.; Yuan, X.W.; Cai, Y.L.; Fan, Z.Y. Studies on Taiwan Chinese pangolin. Taibei Zoo: Taibei, China, 2001. [Google Scholar]
- Heath, M.E.; Hammel, H.T. Body temperature and rate of O2 consumption in Chinese pangolins. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1986, 250, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Nikol’skii, A.A. Relative effects of soil and surface air on mammal burrow temperature: a study of the bobac burrow as an example. Gener. Biol. 2002, 382, 25–27. [Google Scholar]
- Nikol’skii, A.A.; Khutorskoi, M.D. Thermal characteristics of mammalian burrows in summer (using a burrow of the steppe marmot as an example). Gener. Biol. 2001, 378, 240–243. [Google Scholar]
- Nikol’skii, A.A.; Savchenko, G.A. Air temperature changes in a steppe marmot burrow in the summer-autumn period. Russian J. Ecol. 2002, 33, 109–114. [Google Scholar] [CrossRef]
- Wu, S.B.; Liu, N.F.; Ma, G.Z.; Xu, Z.R.; Chen, H. Habitat selection by Chinese pangolin (Manis pentadactyla) in winter in Dawuling Natural Reserve. Mammalia 2003, 67, 493–501. [Google Scholar] [CrossRef]
- Bao, F.; Wu, S.; Su, C.; Yang, L.; Zhang, F.; Ma, G. Air temperature changes in a burrow of Chinese pangolin, Manis pentadactyla, in winter. Folia Zool. 2013, 62, 42–48. [Google Scholar] [CrossRef]
- Okwakol, M.J.N.; Sekamatte, M.B. Review article soil macrofauna research in ecosystems in Uganda. Afr. J. Ecol. 2007, 45, 2–8. [Google Scholar] [CrossRef]
- Ellwood, M.D.; Jones, D.T.; Foster, W.A. Canopy ferns in lowland dipterocarp forest support a prolific abundance of ants, termites, and other invertebrates. Biotropica 2002, 34, 575–583. [Google Scholar] [CrossRef]
- Eggleton, P.; Bignell, D.E.; Sands, W.A.; Mawdsley, N.A.; Lawton, J.H.; Wood, T.G.; Bignell, N.C. The diversity, abundance and biomass of termites under differing levels of disturbance in the Mbalmayo Forest Reserve, southern Cameroon. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 51–68. [Google Scholar]
- Lee, R.H.; Cheung, K.; Fellowes, J.R.; Guenard, B. Insights Into the Chinese Pangolin’s (Manis pentadactyla) diet in a peri-urban habitat: a case study from Hong Kong. Trop. Conserv. Sci. 2017, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, U.B.; Shrestha, B.B.; Shrestha, S. Biodiversity conservation in community forests of Nepal: Rhetoric and reality. Int. J. Biodivers. Conserv. 2010, 2, 98–104. [Google Scholar]
- Kunwar, R.M.; Sharma, S.P. Quantitative analysis of tree species in two community forests of Dolpa district, mid-west Nepal. Himalayan J. Sci. 2004, 2, 23–28. [Google Scholar] [CrossRef]
- Reddy, C.S.; Vazeed, P.S.; Satish, K.V.; Saranya, K.R.L.; Jha, C.S.; Krishna Murthy, Y.V.N. Quantifying nationwide land cover and historical changes in forests of Nepal (1930–2014): implications on forest fragmentation. Biodivers. Conserv. 2018, 27, 91–107. [Google Scholar] [CrossRef]
- Karawita, H.; Perera, P.; Dayawansa, N.; Dias, S. Dietary composition and foraging habitats of the Indian Pangolin (Manis crassicaudata) in a tropical lowland forest-associated landscape in southwest Sri Lanka. Glob. Ecol. Conserv. 2020, e00880. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Saunders, D.A. (Eds.) Reintegrating Fragmented Landscapes: Towards Sustainable Production and Nature Conservation; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Carter, A.S.; Gilmour, D.A. Increase in tree cover in private farmland in central Nepal. Mt. Res. Dev. 1989, 9, 381–391. [Google Scholar] [CrossRef]
- Acharya, K.P. Linking trees on farms with biodiversity conservation in subsistence farming systems in Nepal. Bioderv. Conserv. 2006, 15, 631–646. [Google Scholar] [CrossRef]
- Rimal, B.; Sharma, R.; Kunwar, R.; Keshtkar, H.; Stork, N.E.; Rijal, S.; Rahman, S.A.; Baral, H. Effects of land use and land cover change on ecosystem services in the Koshi River Basin, Eastern Nepal. Ecosyst. Serv. 2019, 38, 100963. [Google Scholar] [CrossRef]
- Dhakal, B.; Bigsby, H.; Cullen, R. Impacts of community forestry development on livestock-based livelihood in Nepal. J. Forest Livelihood 2005, 4, 43–49. [Google Scholar]
- Gautam, A.P.; Webb, E.L.; Shivakoti, G.P.; Zoebisch, M.A. Land use dynamics and landscape change pattern in a mountain watershed in Nepal. Agric. Ecosyst. Environ. 2003, 99, 83–96. [Google Scholar] [CrossRef]
- Clements, G.R.; Lynam, A.J.; Gaveau, D.; Yap, W.L.; Lhota, S.; Goosem, M.; Laurance, S.; Laurance, W.F. Where and how are roads endangering mammals in Southeast Asia’s forests? PLOS ONE 2014, 9, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Chen, M.; Challender, D.W.S.; Waterman, C.; Zhang, C.; Huo, Z.; Liu, H.; Luan, X. Historical data for conservation: reconstructing range changes of Chinese pangolin (Manis pentadactyla) in eastern China (1970–2016). Proc. R. Soc. B Biol. Sci. 2018, 285, 20181084. [Google Scholar] [CrossRef] [Green Version]
- Thapa, P.; Khatiwada, A.P.; Nepali, S.C.; Paudel, S. Chinese Pangolin (Manis pentadactyla) in Nangkholyang VDC, Taplejung, Eastern Nepal. Am. J. Zool. Res. 2014, 2, 16–21. [Google Scholar]
- Bajracharya, S.B.; Furley, P.A.; Newton, A.C. Impacts of community-based conservation on local communities in the Annapurna Conservation Area, Nepal. Biodivers. Conserv. 2006, 15, 2765–2786. [Google Scholar] [CrossRef]
- Budhathoki, P. Linking communities with conservation in developing countries: Buffer zone management initiatives in Nepal. Oryx 2004, 38, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.P.; Belant, J.L.; Shaner, P.J.L. Attitudes towards conservation of the Endangered red panda Ailurus fulgens in Nepal: A case study in protected and non-protected areas. Oryx 2019, 53, 542–547. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Cover Type | Chinese Pangolin’s Potential Suitable Habitat (Km2) | Total Area (Km2) | ||||
|---|---|---|---|---|---|---|
| Tarai < 300 m | Siwalik 300–1000 m | Mid-hill 1000–3000 m | Mid-mountain 3000–5000 m | High mountain > 5000 m | ||
| Settlements | 17.54 | 10.68 | 245.60 | 14.01 | 0.00 | 287.83 |
| Cultivated | 260.64 | 578.83 | 9868.28 | 1571.90 | 0.04 | 12279.69 |
| Forest | 189.69 | 2683.96 | 8841.47 | 2223.88 | 6.57 | 13945.56 |
| Shrub | 1.88 | 93.97 | 877.04 | 255.10 | 0.37 | 1228.36 |
| Barren | 24.27 | 143.43 | 179.67 | 15.22 | 0.13 | 362.72 |
| Grassland | 0.13 | 25.24 | 323.27 | 168.10 | 2.22 | 518.96 |
| Others | 7.71 | 21.91 | 86.65 | 28.64 | 0.31 | 145.22 |
| Total | 501.87 | 3558.01 | 20421.98 | 4276.84 | 9.64 | 28768.35 |
| Land Cover Type | Chinese Pangolin’s Suitable Habitat (Km2) | Total Area (km2) | |||
|---|---|---|---|---|---|
| Province 1 | Province 2 | Bagmati Province | Gandaki Province | ||
| Settlement | 35.45 | 0.26 | 187.68 | 64.44 | 287.83 |
| Cultivated | 5452.41 | 29.61 | 4576.03 | 2223.10 | 12281.16 |
| Forest | 5358.27 | 527.41 | 5506.59 | 2551.55 | 13943.83 |
| Bush | 190.04 | 55.46 | 815.11 | 167.95 | 1228.57 |
| Barren | 120.12 | 27.33 | 192.32 | 22.91 | 362.68 |
| Grassland | 69.24 | 2.48 | 341.76 | 105.54 | 519.02 |
| Others | 53.70 | 3.55 | 72.23 | 15.78 | 145.25 |
| Total | 11279.23 | 646.11 | 11691.72 | 5151.29 | 28768.35 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, H.P.; Rimal, B.; Zhang, M.; Sharma, S.; Poudyal, L.P.; Maharjan, S.; Kunwar, R.; Kaspal, P.; Bhandari, N.; Baral, L.; et al. Potential Distribution of the Critically Endangered Chinese Pangolin (Manis pentadactyla) in Different Land Covers of Nepal: Implications for Conservation. Sustainability 2020, 12, 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/su12031282
Sharma HP, Rimal B, Zhang M, Sharma S, Poudyal LP, Maharjan S, Kunwar R, Kaspal P, Bhandari N, Baral L, et al. Potential Distribution of the Critically Endangered Chinese Pangolin (Manis pentadactyla) in Different Land Covers of Nepal: Implications for Conservation. Sustainability. 2020; 12(3):1282. https://0-doi-org.brum.beds.ac.uk/10.3390/su12031282
Chicago/Turabian StyleSharma, Hari Prasad, Bhagawat Rimal, Mingxia Zhang, Sandhya Sharma, Laxman Prasad Poudyal, Sujan Maharjan, Ripu Kunwar, Prativa Kaspal, Namrata Bhandari, Laxmi Baral, and et al. 2020. "Potential Distribution of the Critically Endangered Chinese Pangolin (Manis pentadactyla) in Different Land Covers of Nepal: Implications for Conservation" Sustainability 12, no. 3: 1282. https://0-doi-org.brum.beds.ac.uk/10.3390/su12031282