Fine-Scale Distribution Patterns of Phragmites australis Populations Across an Environmental Gradient in the Salt Marsh Wetland of Dunhuang, China

Abstract
:1. Introduction
2. Materials and Methods
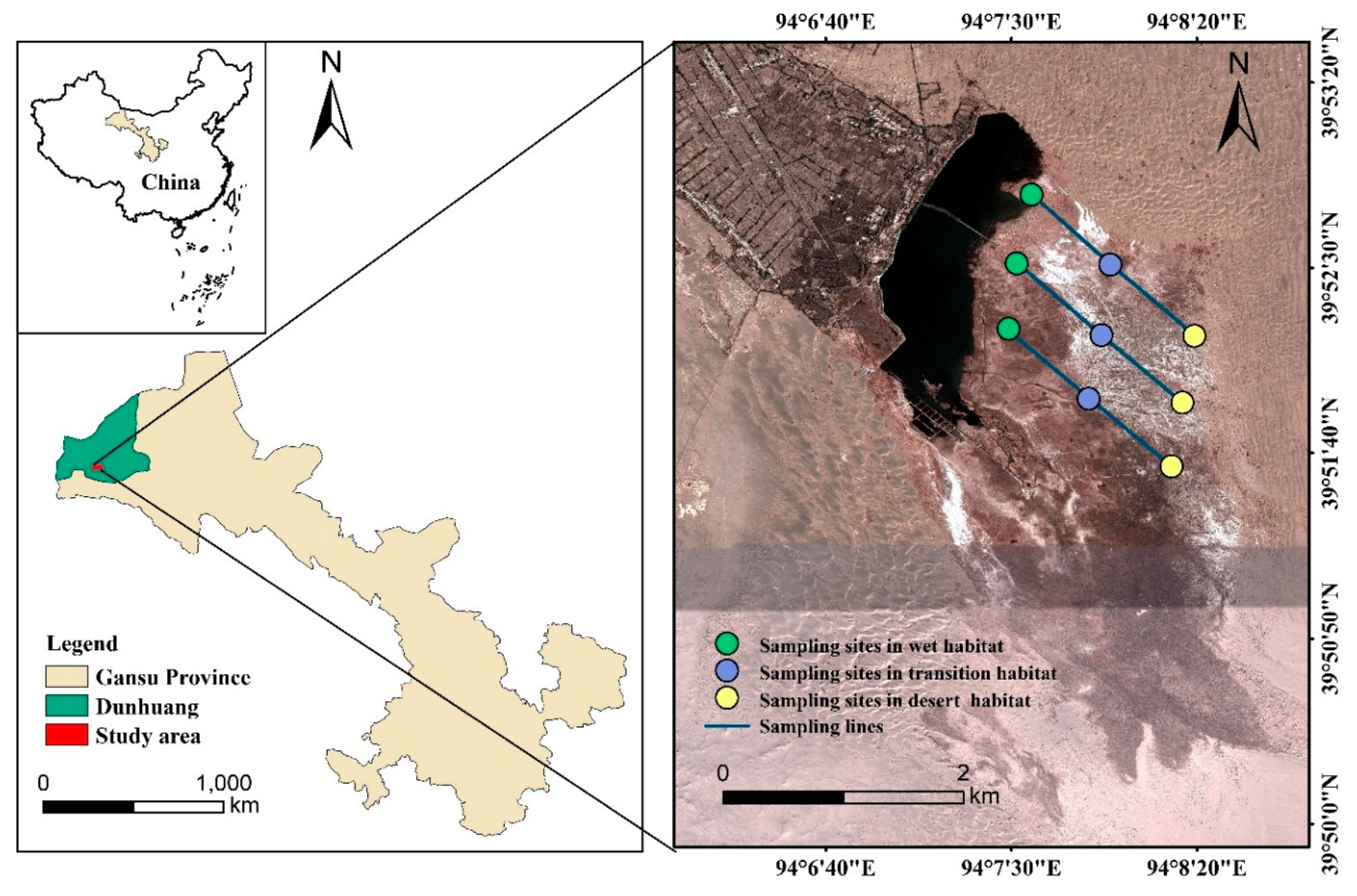
2.1. Overview of the Study Area
2.2. Plant Sampling and Measurement
2.3. Data processing
2.3.1. Point Pattern Analysis
2.3.2. Data Treatment and Statistical Analysis
3. Results
3.1. Analysis of the Soil Environmental Factors in the Study Area
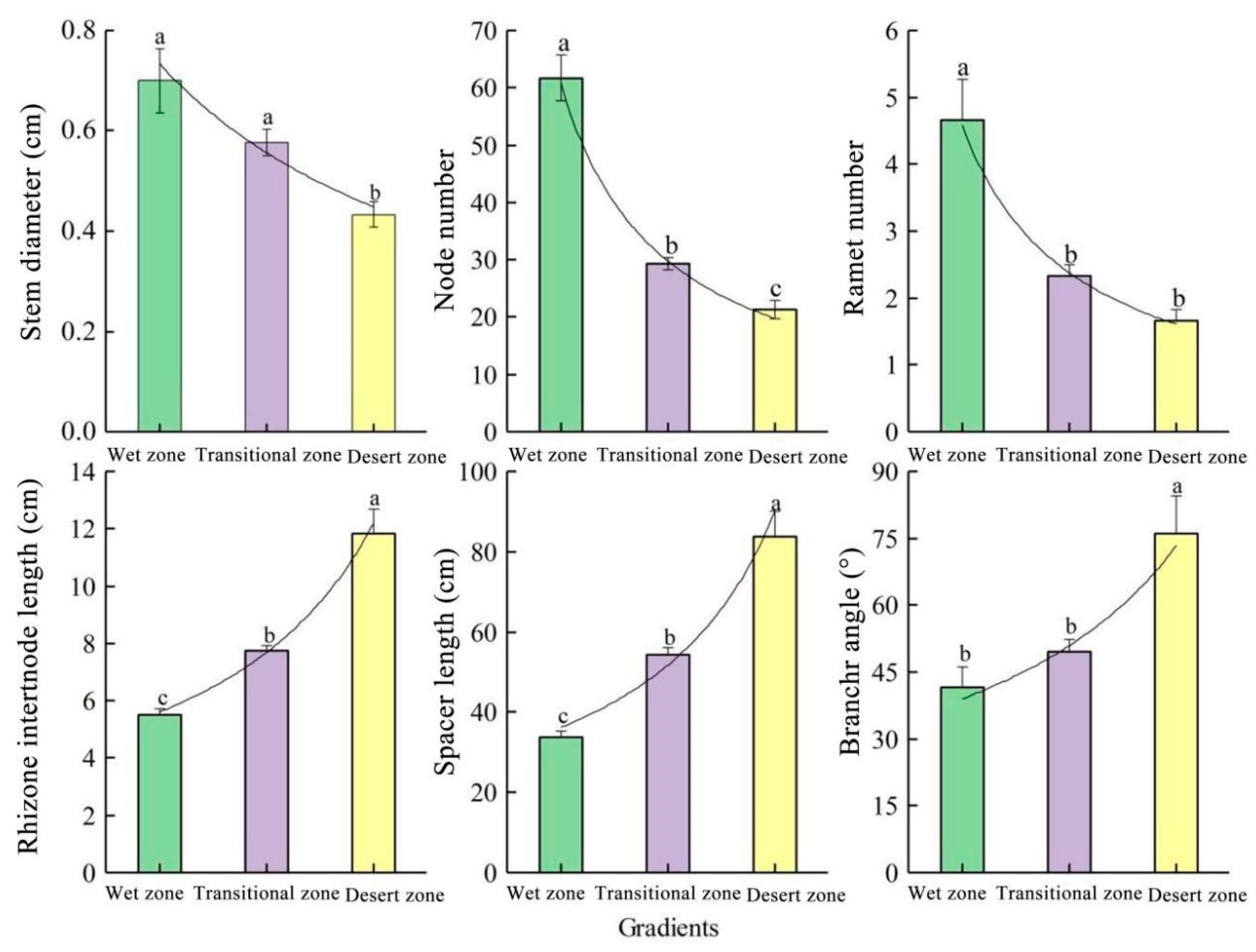
3.2. Analysis of the Clonal Characteristics of Phragmites australis
3.3. Analysis of the Population Characteristics of Phragmites australis
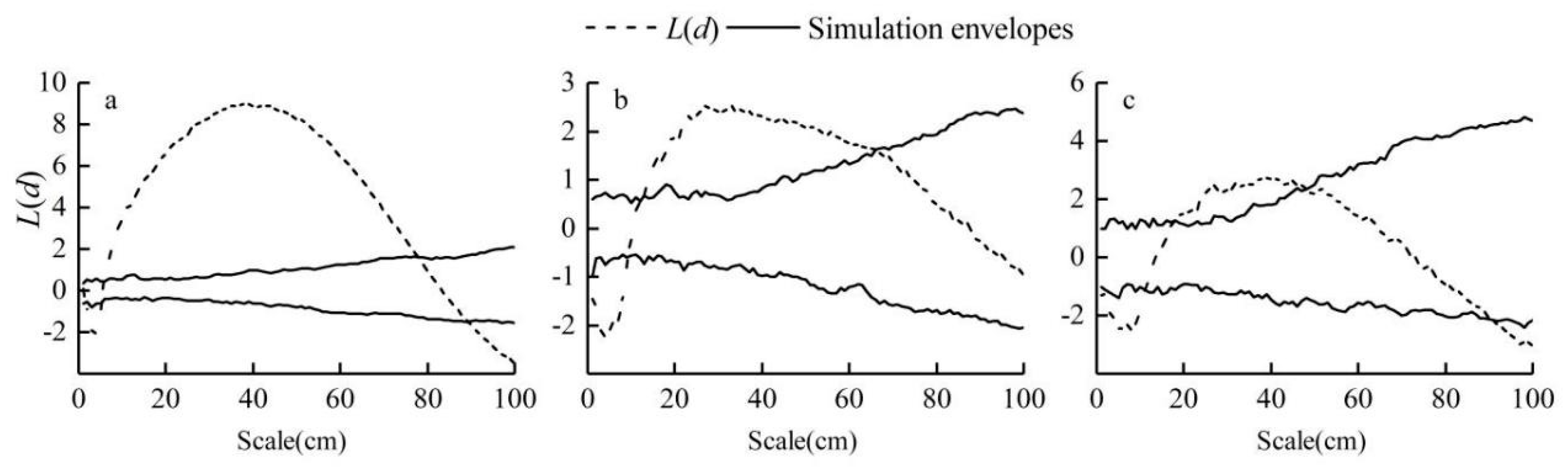
3.4. Analysis of the Fine-Scale Spatial Point Pattern of Phragmites australis
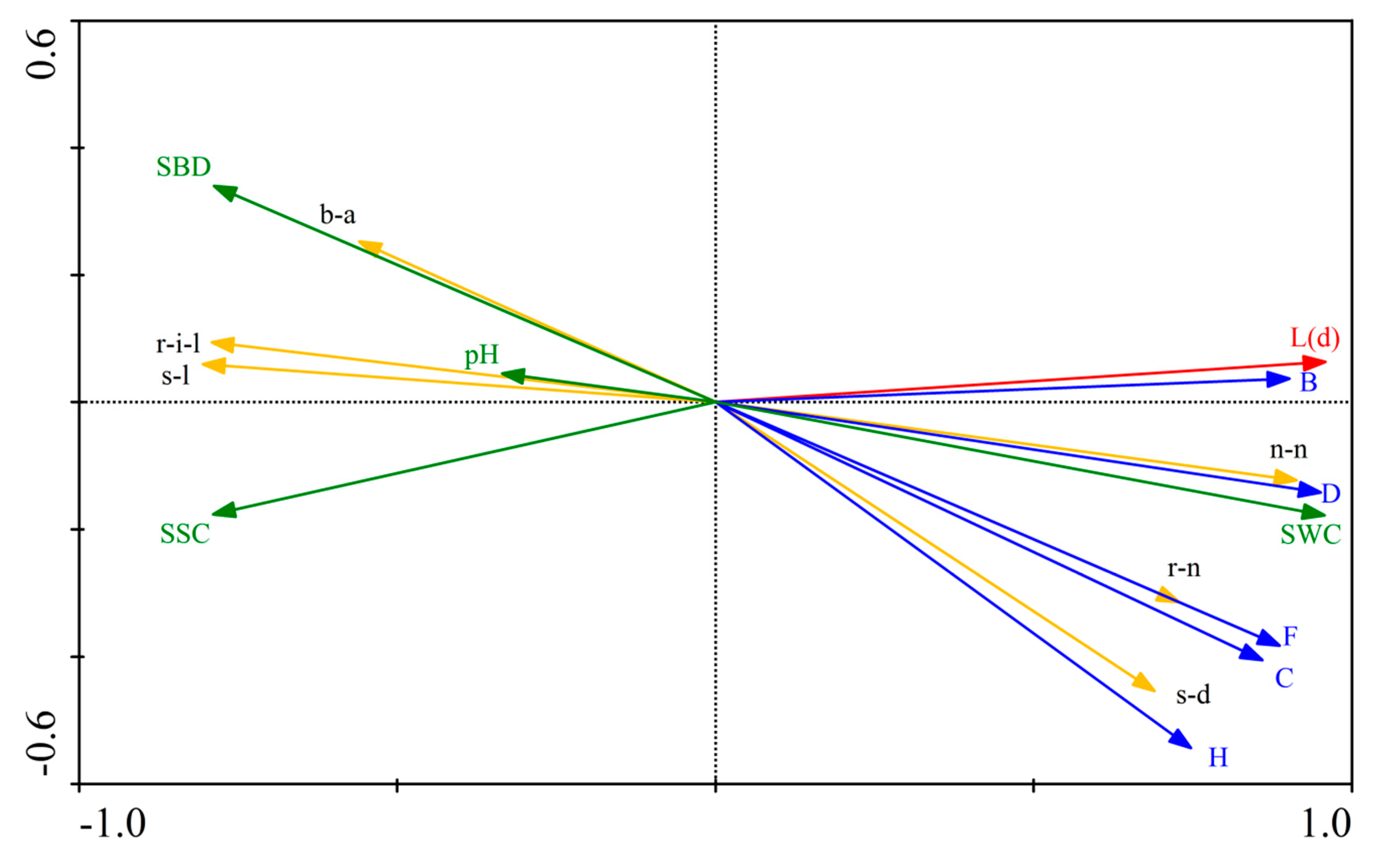
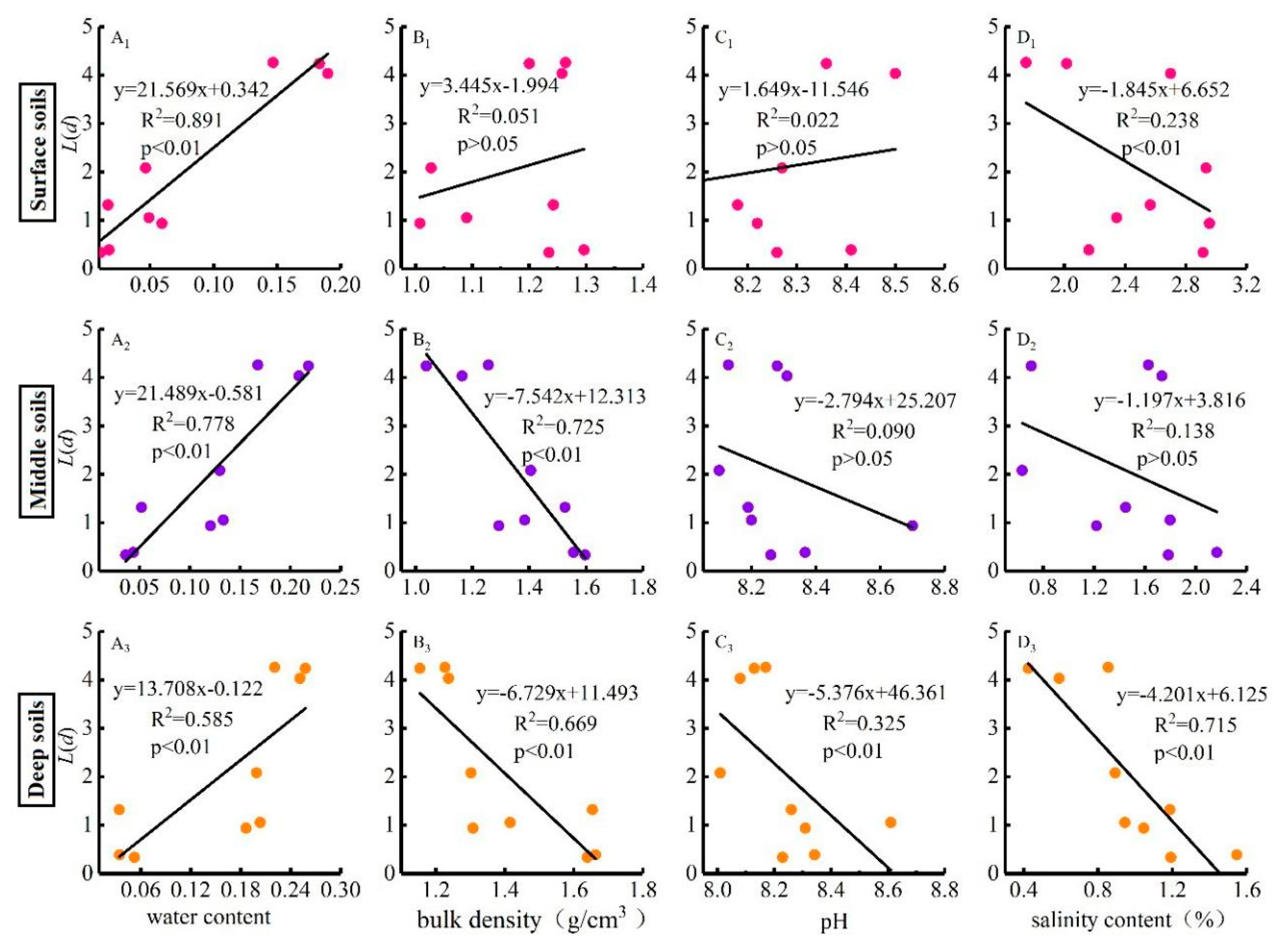
3.5. Relationships of the Spatial Distribution Pattern of Phragmites australis with the Environmental Factors, Population Characteristics and Clonal Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holm, B.; Heinsoo, K. Municipal wastewater application to short rotation coppice of willows–treatment efficiency and clone response in Estonian case study. Biomass Bioenergy 2013, 57, 126–135. [Google Scholar] [CrossRef]
- Pinno, B.D.; Wilson, S.D. Nitrogen translocation between clonal mother and daughter trees at a grassland–forest boundary. Plant Ecol. 2014, 215, 347–354. [Google Scholar] [CrossRef]
- Dong, M.; Yu, F.H.; Alpert, P. Ecological consequences of plant clonality. Ann. Bot. 2014, 114, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Dong, M. Clonal plants and plant species diversity in wetland ecosystems in China. J. Veg. Sci. 2002, 13, 237–244. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prach, K.; Pysek, P.; Smilauer, P. Changes in species traits during succession: A search for pattern. Oikos 1997, 79, 201–205. [Google Scholar] [CrossRef]
- Yu, F.H.; Wang, N.; He, W.M.; Dong, M. Effects of clonal integration on species composition and biomass of sand dune communities. J. Arid Environ. 2010, 74, 632–637. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Song, Y.B.; Yu, F.H.; Dong, M. Plant traits and ecosystem effects of clonality: A new research agenda. Ann. Bot. 2014, 114, 369–376. [Google Scholar] [CrossRef] [Green Version]
- López, R.P.; Larrea-Alcázar, D.; Zenteno-Ruiz, F. Spatial pattern analysis of dominant species in the Prepuna: Gaining insight into community dynamics in the semi-arid, subtropical Andes. J. Arid Environ. 2010, 74, 1534–1539. [Google Scholar] [CrossRef]
- Lee, K.; Kim, S.; Shin, Y.; Choung, Y. Spatial pattern and association of tree species in a mixed Abies holophylla-broadleaved deciduous forest in Odaesan National Park. J. Plant Biol. 2012, 55, 242–250. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166. [Google Scholar] [CrossRef]
- Gopal, B. Future of wetlands in tropical and subtropical Asia, especially in the face of climate change. Aquat. Sci. 2013, 75, 39–61. [Google Scholar] [CrossRef]
- Weis, J.S.; Weis, P. Metal uptake, transport and release by wetland plants: Implications for phytoremediation and restoration. Environ. Int. 2004, 30, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Windham, L.; Weis, J.S.; Weis, P. Lead uptake, distribution, and effects in two dominant salt marsh macrophytes, Spartina alterniflora (cordgrass) and Phragmites australis (common reed). Mar. Pollut. Bull. 2001, 42, 811–816. [Google Scholar] [CrossRef]
- Hansen, D.L.; Lambertini, C.; Jampeetong, A.; Brix, H. Clone-specific differences in Phragmites australis: Effects of ploidy level and geographic origin. Aquat. Bot. 2007, 86, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Saltonstall, K. Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc. Nat. Acad. Sci. USA 2002, 99, 2445–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douhovnikoff, V.; Hazelton, E.L.G. Clonal growth: Invasion or stability? A comparative study of clonal architecture and diversity in native and introduced lineages of Phragmites australis (Poaceae). Am. J. Bot. 2014, 101, 1577–1584. [Google Scholar] [CrossRef]
- Wang, C.; Xiao, R.; Cui, Y.; Ma, Z.; Guo, Y.; Wang, Q.; Xiu, Y.J.; Zhang, M. Photosynthate-13C allocation in the plant-soil system after 13C–pulse labeling of Phragmites australis in different salt marshes. Geoderma 2019, 347, 252–261. [Google Scholar] [CrossRef]
- Ali, N.A.; Bernal, M.P.; Ater, M. Tolerance and bioaccumulation of copper in Phragmites australis and Zea mays. Plant Soil 2002, 239, 103–111. [Google Scholar] [CrossRef]
- Pagter, M.; Bragato, C.; Brix, H. Tolerance and physiological responses of Phragmites australis to water deficit. Aquat. Bot. 2005, 81, 285–299. [Google Scholar] [CrossRef]
- Amsberry, L.; Baker, M.A.; Bertness, E.M.D. Clonal integration and the expansion of Phragmites australis. Ecol. Appl. 2000, 10, 1110–1118. [Google Scholar] [CrossRef]
- Reijers, V.C.; van den Akker, M.; Cruijsen, P.M.; Lamers, L.P.; van der Heide, T. Intraspecific facilitation explains the persistence of Phragmites australis in modified coastal wetlands. Ecosphere 2019, 10, e02842. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhang, J.; Zhang, C.; Yue, Q.; Li, Y.; Li, C. Equilibrium and kinetic studies of methyl orange and methyl violet adsorption on activated carbon derived from Phragmites australis. Desalination 2010, 252, 149–156. [Google Scholar] [CrossRef]
- Bonanno, G.; Giudice, R.L. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. Ecol. Indic. 2010, 10, 639–645. [Google Scholar] [CrossRef]
- Rezania, S.; Park, J.; Rupani, P.F.; Darajeh, N.; Xu, X.; Shahrokhishahraki, R. Phytoremediation potential and control of Phragmites australis as a green phytomass: An overview. Environ. Sci. Pollut. Res. 2019, 26, 7428–7441. [Google Scholar] [CrossRef]
- Houle, G. A multivariate analysis of fine-scale species density in the plant communities of a saltwater lagoon—The importance of disturbance intensity. Oikos 2005, 111, 465–472. [Google Scholar] [CrossRef]
- Cheng, D.L.; Wang, G.X.; Chen, B.M.; Wei, X.P. Positive interactions: Crucial organizers in a plant community. J. Integr. Plant Biol. 2006, 48, 128–136. [Google Scholar] [CrossRef]
- Tu, H.R.; Li, J.F.; Liu, R.H.; Liang, S.C.; Lan, Z.N.; Zhang, X.Y.; Kang, X.D.; Jiang, Y. Spatial distribution patterns and association of Loropetalum chinense population in karst hills of Guilin, Southwest China. J. Appl. Ecol. 2019, 30, 2621–2630. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, J.; Wiegand, T.; Traveset, A. Adult proximity and frugivore’s activity structure the spatial pattern in an endangered plant. Funct. Ecol. 2012, 26, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Lilleskov, E.A.; Bruns, T.D.; Horton, T.R.; Lee Taylor, D.; Grogan, P. Detection of forest stand-level spatial structure in ectomycorrhizal fungal communities. Fems Microbiol. Ecol. 2004, 49, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Wilschut, L.I.; Laudisoit, A.; Hughes, N.K.; Addink, E.A.; de Jong, S.M.; Heesterbeek, H.A.; Begon, M. Spatial distribution patterns of plague hosts: Point pattern analysis of the burrows of great gerbils in Kazakhstan. J. Biogeogr. 2015, 42, 1281–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Xiao, R.; Shortridge, A.; Wu, J. Spatial Point Pattern Analysis of Human Settlements and Geographical Associations in Eastern Coastal China—A Case Study. Int. J. Environ. Res. Public Health 2014, 11, 2818–2833. [Google Scholar] [CrossRef] [PubMed]
- Haltigin, T.; Pollard, W.; Dutilleul, P. Comparison of ground-and aerial-based approaches for quantifying polygonal terrain network geometry on Earth and Mars via spatial point pattern analysis. Planet. Space Sci. 2010, 58, 1636–1649. [Google Scholar] [CrossRef]
- Kui, L.; Li, F.; Moore, G.; West, J. Can the riparian invader, a rundo donax, benefit from clonal integration? Weed Res. 2013, 53, 370–377. [Google Scholar] [CrossRef]
- Wiegand, T.; Kissling, W.D.; Cipriotti, P.A.; Aguiar, M.R. Extending point pattern analysis for objects of finite size and irregular shape. J. Ecol. 2006, 94, 825–837. [Google Scholar] [CrossRef]
- Mack, R.N.; Harper, J.L. Interference in dune annuals: Spatial pattern and neighbourhood effects. J. Ecol. 1977, 65, 345–363. [Google Scholar] [CrossRef] [Green Version]
- Silander, J.A.; Pacala, S.W. Neighborhood predictors of plant performance. Oecologia 1985, 66, 256–263. [Google Scholar] [CrossRef]
- Ripley, B.D. Modelling spatial patterns. J. R. Stat. Soc. Ser. B 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Gatrell, A.C.; Bailey, T.C.; Diggle, P.J.; Rowlingson, B.S. Spatial point pattern analysis and its application in geographical epidemiology. Trans. Inst. Br. Geogr. 1996, 21, 256–274. [Google Scholar] [CrossRef]
- Molofsky, J. The effect of nutrients and spacing on neighbor relations in Cardamine pensylvanica. Oikos 1999, 84, 506–514. [Google Scholar] [CrossRef]
- Perelman, S.B.R.; León, R.J.C.; Oesterheld, M. Cross-scale vegetation patterns of Flooding Pampa grasslands. J. Ecol. 2001, 89, 562–577. [Google Scholar] [CrossRef] [Green Version]
- Purves, D.W.; Law, R. Fine-scale spatial structure in a grassland community: Quantifying the plant’s-eye view. J. Ecol. 2002, 90, 121–129. [Google Scholar] [CrossRef]
- Benot, M.L.; Bittebiere, A.K.; Ernoult, A.; Clement, B.; Mony, C. Fine-scale spatial patterns in grassland communities depend on species clonal dispersal ability and interactions with neighbours. J. Ecol. 2013, 101, 626–636. [Google Scholar] [CrossRef]
- De Kroons, H.; Hutchings, M.J. Morphological plasticity in clonal plants: The foraging concept reconsidered. J. Ecol. 1995, 83, 143–152. [Google Scholar] [CrossRef]
- Diggle, P.J. Statistical Analysis of Spatial Point Patterns, 1st ed.; Academic Press: London, UK, 1983; pp. 136–245. [Google Scholar]
- Loosmore, N.B.; Ford, E.D. Statistical inference using the G or K point pattern spatial statistics. Ecology 2006, 87, 1925–1931. [Google Scholar] [CrossRef] [Green Version]
- Morris, C. Multivariate analysis of ecological data using CANOCO 5. Afr. J. Range Forage Sci. 2015, 32, 289–290. [Google Scholar] [CrossRef]
- Zou, K.H.; Tuncali, K.; Silverman, S.G. Correlation and simple linear regression. Radiology 2003, 227, 617–628. [Google Scholar] [CrossRef]
- Hirose, T. A vegetative plant growth model: Adaptive significance of phenotypic plasticity in matter partitioning. Funct. Ecol. 1987, 1, 195–202. [Google Scholar] [CrossRef]
- Sterner, R.W.; Ribic, C.A.; Schatz, G.E. Testing for life historical changes in spatial patterns of four tropical tree species. J. Ecol. 1986, 74, 621–633. [Google Scholar] [CrossRef]
- Dale, M.R.T. Spatial Pattern Analysis in Plant Ecology, 1st ed.; Cambridge University Press: Cambridge, UK, 2000; pp. 168–205. [Google Scholar]
- Eccles, N.S.; Esler, K.J.; Cowling, R.M. Spatial pattern analysis in Namaqualand desert plant communities: Evidence for general positive interactions. Plant Ecol. 1999, 142, 71–85. [Google Scholar] [CrossRef]
- Bisigato, A.J.; Villagra, P.E.; Ares, J.O.; Rossi, B.E. Vegetation heterogeneity in Monte Desert ecosystems: A multi-scale approach linking patterns and processes. J. Arid Environ. 2009, 73, 182–191. [Google Scholar] [CrossRef]
- De Kroon, H.; Knops, J. Habitat exploration through morphological plasticity in two chalk grassland perennials. Oikos 1990, 59, 39–49. [Google Scholar] [CrossRef]
- Ye, X.H.; Yu, F.H.; Dong, M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann. Bot. 2006, 98, 187–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, J.J.; Zhao, C.Z.; Dong, X.G.; Yang, Q.; Cha, G.D. Response of Kobresia tibetica and Kobresia humilis population spatial pattern to soil moisture in Chinese. Chin. J. Ecol. 2014, 33, 1734–1740. [Google Scholar] [CrossRef]
- Wang, X.P.; Zhao, C.Z.; Wang, J.W.; Zhao, L.C.; Wen, J. Aggregated spatial patterns and characteristics of Suaeda corniculata in Qin wangchuan salt marsh in Chinese. Acta Ecol. Sin. 2018, 38, 3943–3951. [Google Scholar] [CrossRef]
- Pottier, J.; Evette, A. Spatial pattern and process at the plant neighbourhood scale: Insights from communities dominated by the clonal grass Elymus repens (L.) Gould. J. Veg. Sci. 2011, 22, 973–982.61. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Uria-Diez, J.; Wiegand, K.; Michalet, R. Spatial distribution and association patterns in a tropical evergreen broad-leaved forest of north-central Vietnam. J. Veg. Sci. 2016, 27, 318–327. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, X.; Guan, H.; Meng, Y.; Ruan, J.; Wang, Z. Effect of Thinning on the Spatial Structure of a Larix gmelinii Rupr. Secondary Forest in the Greater Khingan Mountains. Forests 2018, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- De Araújo, E.J.G.; Lima, D.O.; Curto, R.D.A.; Silva, E.V.; Júnior, J.C.A. Space pattern of forest species and its relationship with agricultural factors agroflorestal System. Floresta 2019, 49, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Bliss, K.M.; Jones, R.H.; Mitchell, R.J.; Mou, P.P. Are competitive interactions influenced by spatial nutrient heterogeneity and root foraging behavior? New Phytol. 2002, 154, 409–417. [Google Scholar] [CrossRef]
- McNickle, G.G.; Clair, C.C.S.; Cahill Jr, J.F. Focusing the metaphor: Plant root foraging behaviour. Trends Ecol. Evol. 2009, 24, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Wang, W.; Liang, C.Z. Changes in the population spatial distribution pattern of Leymus chinensis in degraded steppe community during restorative succession in Inner Mongolia, China. Chin. J. Plant Ecol. 2009, 33, 63–70. [Google Scholar] [CrossRef]
- Gao, F.Y.; Zhao, C.Z. In the process of grassland degradation the spatial pattern and spatial association of dominant species. Acta Ecol. Sin. 2012, 32, 6661–6669. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Berninger, F.; Koskela, J.; Sonninen, E. Drought responses of Eucalyptus microtheca provenances depend on seasonality of rainfall in their place of origin. Funct. Plant Biol. 2000, 27, 231–238. [Google Scholar] [CrossRef]
- Bargali, K.; Tewari, A. Growth and water relation parameters in drought-stressed Coriaria nepalensis seedlings. J. Arid Environ. 2004, 58, 505–512. [Google Scholar] [CrossRef]
- Shi, L.X.; Guo, J.X. Changes in photosynthetic and growth characteristics of Leymus chinensis community along the retrogression on the Songnen grassland in northeastern China. Photosynthetica 2006, 44, 542. [Google Scholar] [CrossRef]
- Zhang, H.; Irving, L.J.; McGill, C.; Matthew, C.; Zhou, D.; Kemp, P. The effects of salinity and osmotic stress on barley germination rate: Sodium as an osmotic regulator. Ann. Bot. 2010, 106, 1027–1035. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer Depth | Wet Zone | Transition Zone | Desert Zone | |
|---|---|---|---|---|
| Soil water content (%) | 0–30cm | 17.34 ± 0.68 a | 6.49 ± 0.20 b | 1.54 ± 0.10 c |
| 30–60cm | 19.8 ± 0.77 a | 12.81 ± 0.19 b | 4.42 ± 0.23 c | |
| 60–100cm | 24.32. ± 0.57 a | 19.63 ± 0.25 b | 4.04 ± 0.30 c | |
| 0–100cm | 20.49 ± 0.66 a | 12.54 ± 0.09 b | 3.33 ± 0.03 c | |
| Soil bulk density (g/cm3) | 0–30cm | 2.15 ± 0.14 b | 2.74 ± 0.10 a | 2.55 ± 0.11 a |
| 30–60cm | 1.36 ± 0.16 b | 1.22 ± 0.17 b | 1.80 ± 0.10 a | |
| 60–100cm | 0.62 ± 0.06 c | 0.96 ± 0.02 b | 1.31 ± 0.06 a | |
| 0–100cm | 1.2 ± 0.02 c | 1.25 ± 0.01 b | 1.49 ± 0.01 a | |
| Soil pH | 0–30cm | 8.31 ± 0.07 a | 8.19 ± 0.03 a | 8.28 ± 0.03 a |
| 30–60cm | 8.24 ± 0.03 a | 8.33 ± 0.10 a | 8.27 ± 0.03 a | |
| 60–100cm | 8.13 ± 0.01 b | 8.31 ± 0.09 a | 8.28 ± 0.02 a | |
| 0–100cm | 8.22 ± 0.03 a | 8.28 ± 0.04 a | 8.28 ± 0.02 a | |
| Soil salinity content (%) | 0–30cm | 1.24 ± 0.01 b | 1.04 ± 0.01 a | 1.26 ± 0.01 a |
| 30–60cm | 1.15 ± 0.03 c | 1.36 ± 0.02 b | 1.56 ± 0.01 a | |
| 60–100cm | 1.21 ± 0.01 c | 1.34 ± 0.02 b | 1.65 ± 0.01 a | |
| 0–100cm | 1.38 ± 0.09 c | 1.64 ± 0.04 b | 1.89 ± 0.04 a |
| Biological Characteristics | Wet Zone | Transition Zone | Desert Zone |
|---|---|---|---|
| Cover (%) | 86.67 ± 2.20 a | 71.67 ± 2.20 b | 20.00 ± 1.44 c |
| Frequency (%) | 85.00 ± 1.44 a | 68.33 ± 2.20 b | 20.00 ± 0.01 c |
| Density (plants/m2) | 94.33 ± 3.93 a | 51.33 ± 3.03 b | 28.00 ± 1.04 c |
| Height (cm) | 157.23 ± 11.07 a | 136.37 ± 3.74 a | 68.00 ± 5.11 b |
| Biomass (g/m2) | 560.01 ± 62.01 a | 316.18 ± 13.56 b | 98.47 ± 42.51 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, L.; Li, F.; Liu, X.; Wang, S.; Zhou, Y. Fine-Scale Distribution Patterns of Phragmites australis Populations Across an Environmental Gradient in the Salt Marsh Wetland of Dunhuang, China. Sustainability 2020, 12, 1671. https://0-doi-org.brum.beds.ac.uk/10.3390/su12041671
Jiao L, Li F, Liu X, Wang S, Zhou Y. Fine-Scale Distribution Patterns of Phragmites australis Populations Across an Environmental Gradient in the Salt Marsh Wetland of Dunhuang, China. Sustainability. 2020; 12(4):1671. https://0-doi-org.brum.beds.ac.uk/10.3390/su12041671
Chicago/Turabian StyleJiao, Liang, Fang Li, Xuerui Liu, Shengjie Wang, and Yi Zhou. 2020. "Fine-Scale Distribution Patterns of Phragmites australis Populations Across an Environmental Gradient in the Salt Marsh Wetland of Dunhuang, China" Sustainability 12, no. 4: 1671. https://0-doi-org.brum.beds.ac.uk/10.3390/su12041671