Influences of Nutrient Sources on the Alternation of Nutrient Limitations and Phytoplankton Community in Jiaozhou Bay, Southern Yellow Sea of China


Abstract
:1. Introduction

2. Methods
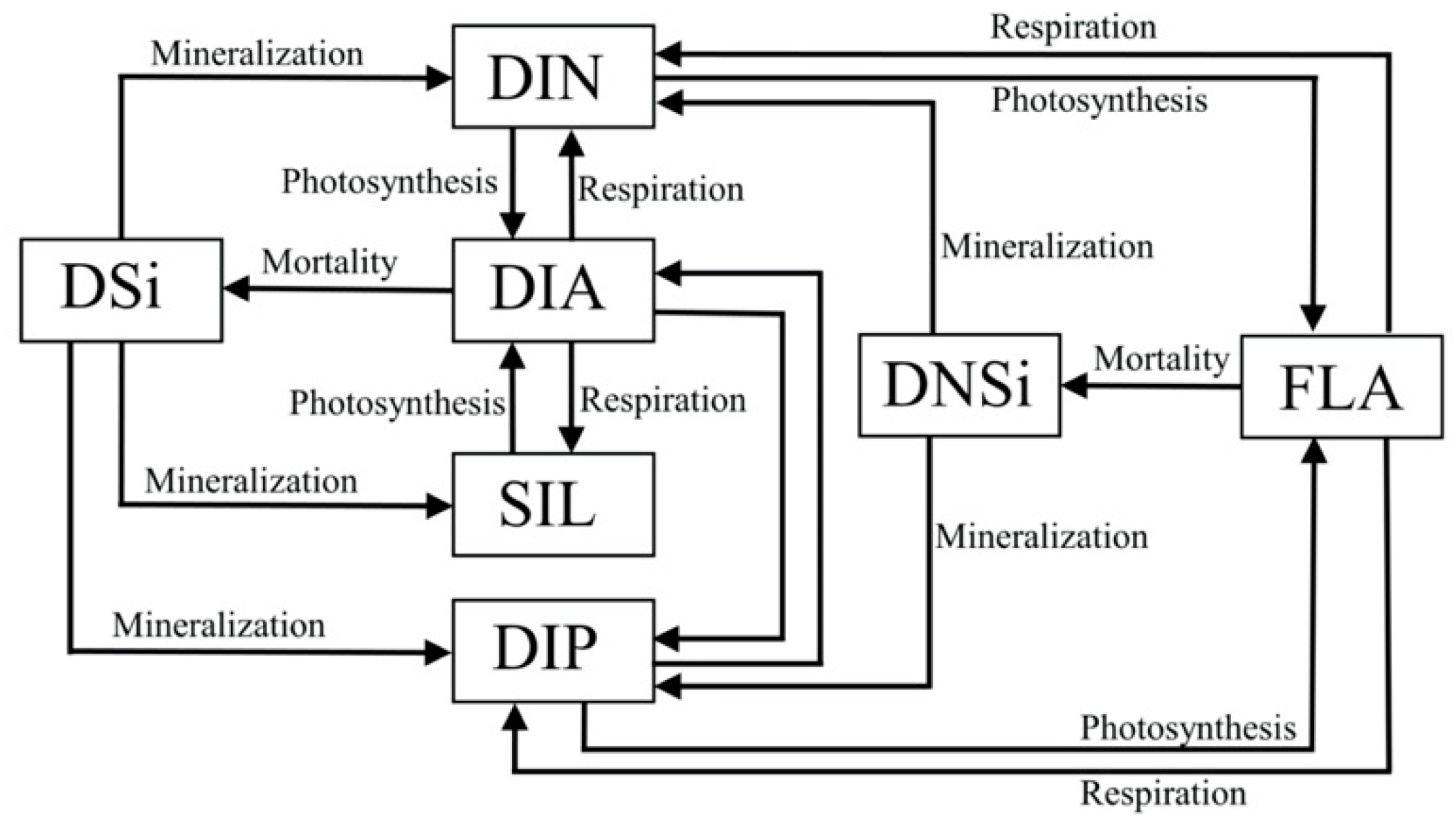
2.1. Model Descriptions
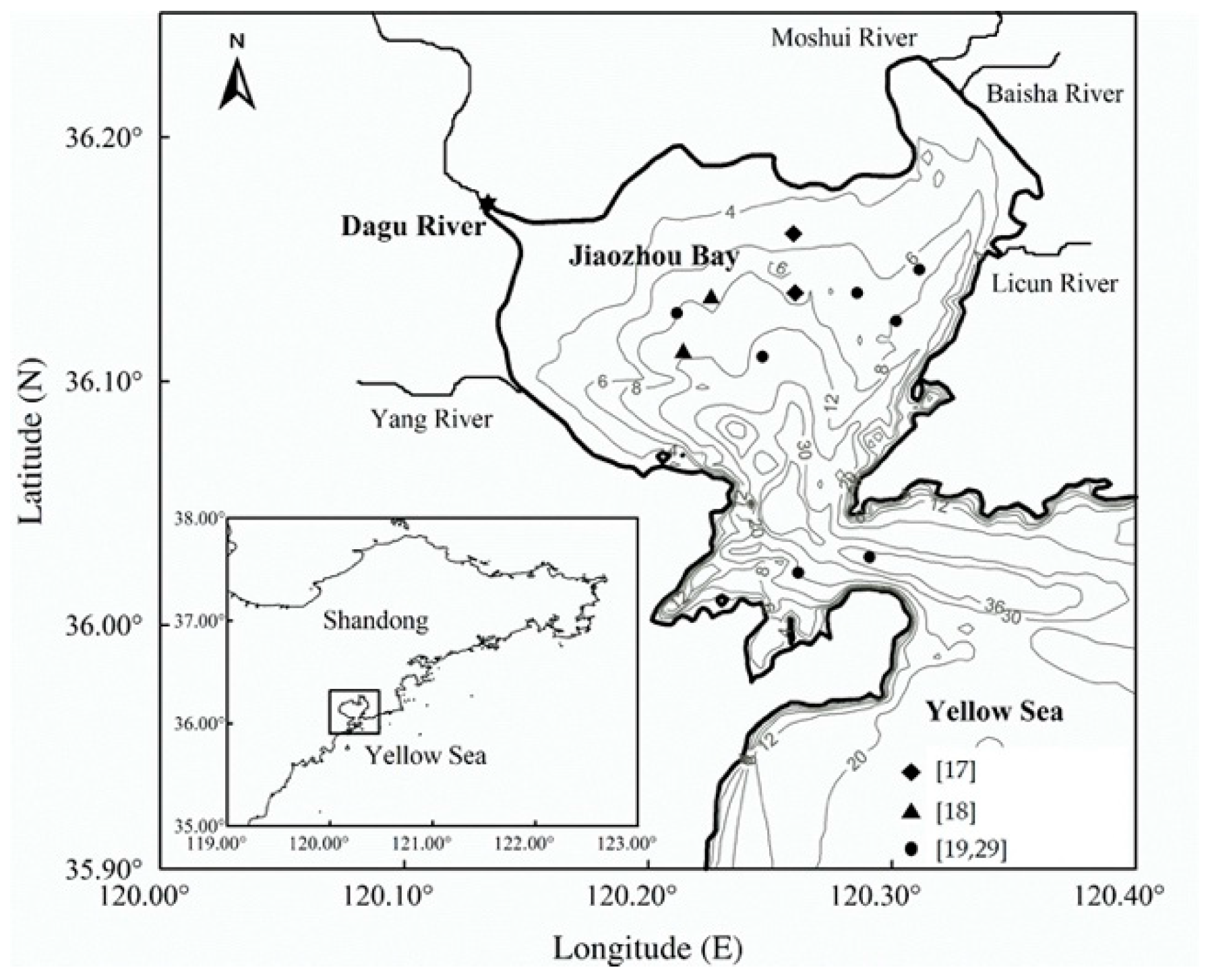
2.2. Model Configuration and Data Sources
2.2.1. CONTROL Run (the 2000s)
2.2.2. NORI, NOWA, NOAQ, and NOAT Runs
2.2.3. The 1960s and the 1980s Runs
3. Results
3.1. Parameter Sensitivity Analysis
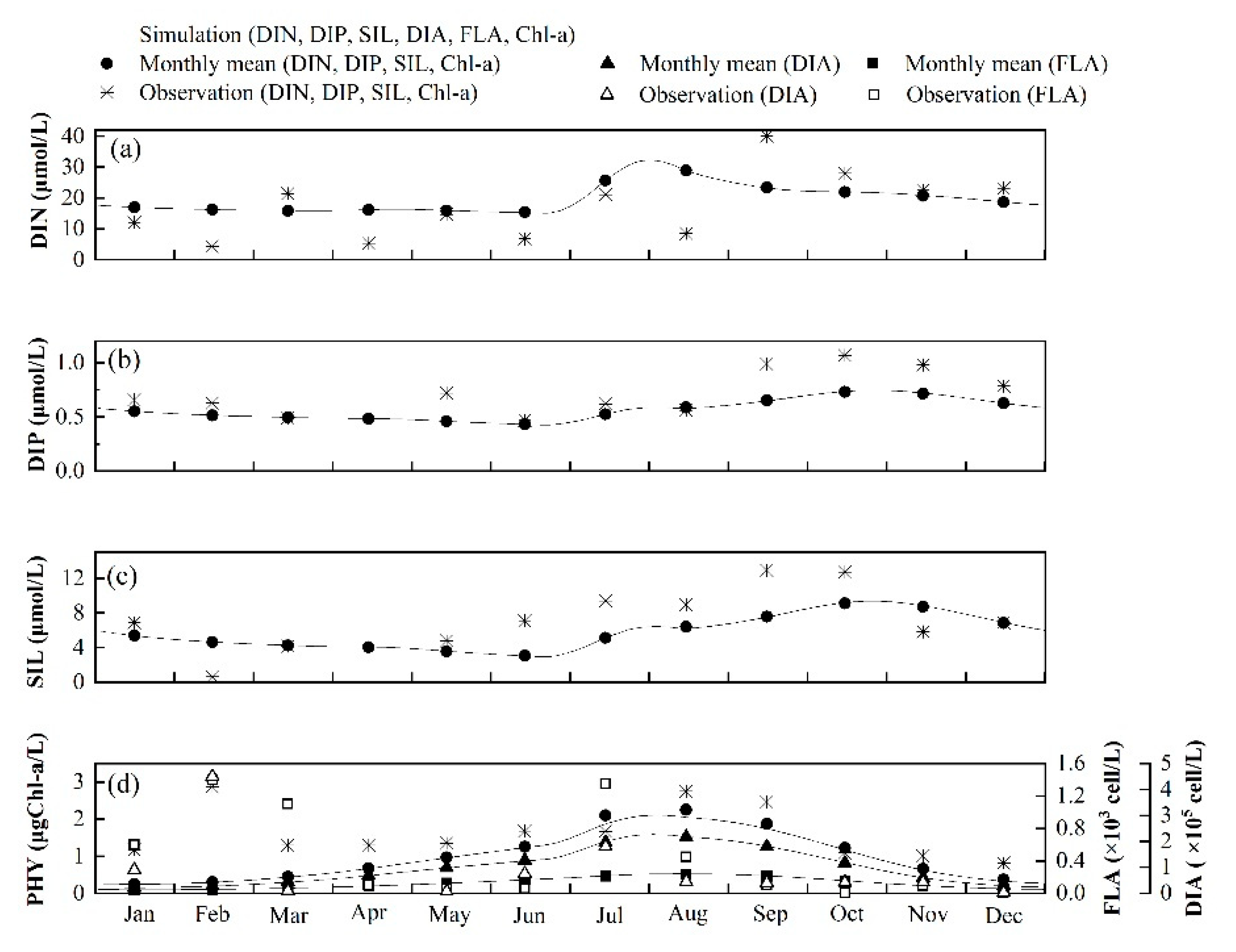
3.2. Seasonal Variations of Nutrient Concentrations and Phytoplankton Biomasses
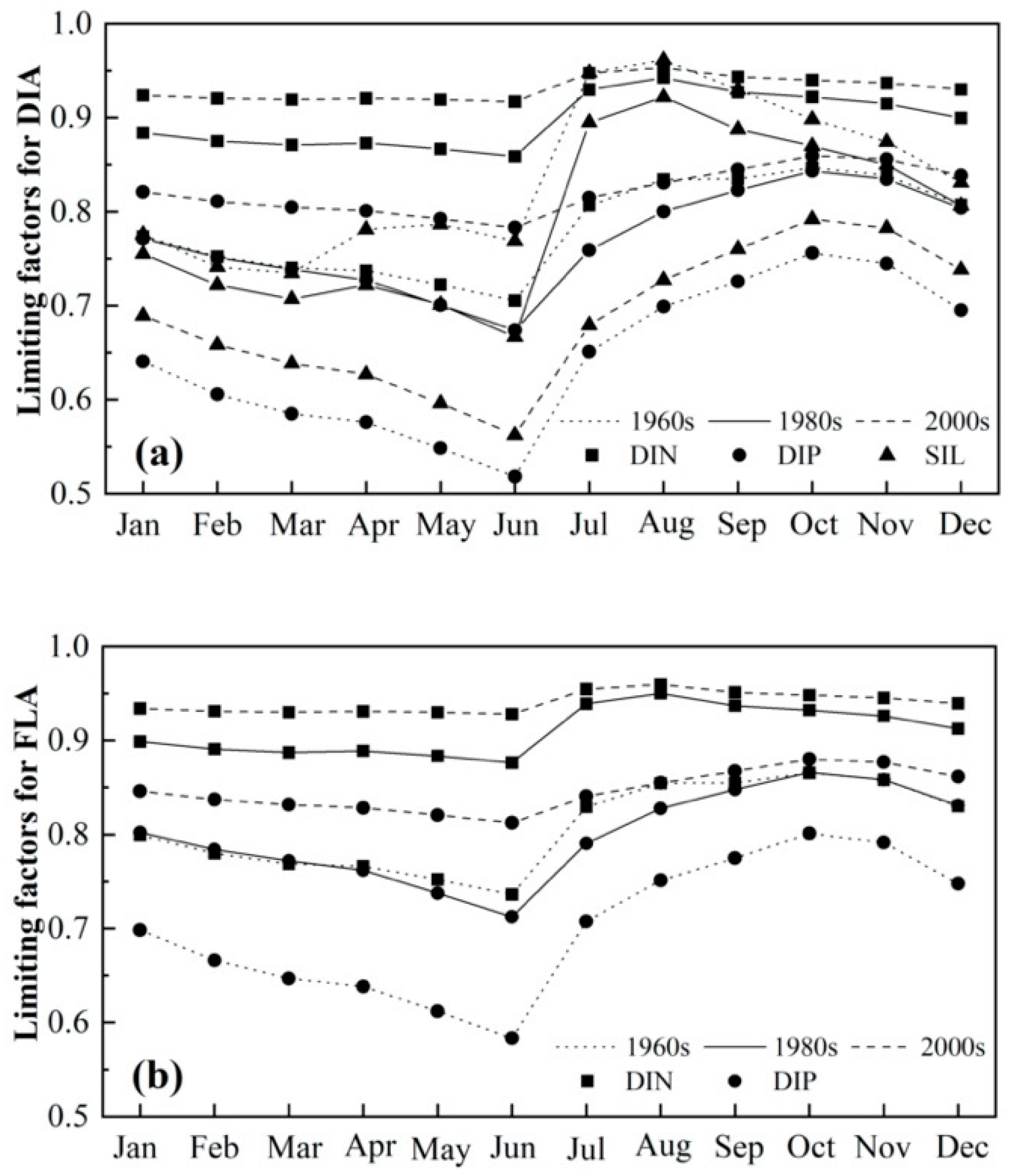
3.3. Seasonal Variations of Nutrient Limitations
3.4. Annual Budgets of Nutrients
3.5. Estimation of the Contributions of Nutrient External Sources on Nutrient Limitations and Phytoplankton Community
3.5.1. The Effects of River Input
3.5.2. The Effects of Aquaculture Activities
3.5.3. The Effects of Wastewater Discharge
3.5.4. The Effects of External Sources on the Cycling Rate of Nutrients in JZB and the Exchange with the YS
3.6. Long-Term Variations of the Nutrient Limitation and Phytoplankton Community
4. Implications and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Gobler, C.J.; Cullison, L.A.; Koch, F.; Harder, T.M.; Krause, J.W. Influence of freshwater flow, ocean exchange, and seasonal cycles on phytoplankton–nutrient dynamics in a temporarily open estuary. Estuar. Coast. Shelf Sci. 2005, 65, 275–288. [Google Scholar] [CrossRef]
- Gong, G.C.; Liu, K.K.; Chiang, K.P.; Hsiung, T.M.; Chang, J.; Chen, C.C.; Hung, C.C.; Chou, W.C.; Chung, C.C.; Chen, H.Y.; et al. Yangtze River floods enhance coastal ocean phytoplankton biomass and potential fish production. Geophys. Res. Lett. 2011, 38, L13603:1–L13603:6. [Google Scholar] [CrossRef] [Green Version]
- Downing, J.A. Marine nitrogen: Phosphorus stoichiometry and the global N:P cycle. Biogeochemistry 1997, 37, 237–252. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman–Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.T.; Chou, T.L. Silicate as the limiting nutrient for phytoplankton in a subtropical eutrophic estuary of Taiwan. Estuar. Coast. Shelf Sci. 2003, 58, 155–162. [Google Scholar] [CrossRef]
- Desmit, X.; Thieu, V.; Billen, G.; Campuzano, F.; Dulière, V.; Garnier, J.; Lassaletta, L.; Ménesguen, A.; Neves, R.; Pinto, L.; et al. Reducing marine eutrophication may require a paradigmatic change. Sci. Total Environ. 2018, 635, 1444–1466. [Google Scholar] [CrossRef]
- Ibáñez, C.; Peñuelas, J. Changing nutrients, changing rivers. Science 2019, 365, 637–638. [Google Scholar] [CrossRef]
- Redfield, A.C.; Ketchum, B.H.; Richards, F.A. The influence of organisms on the composition of seawater. In The Sea; Hill, M.N., Ed.; Interscience Publishers: New York, NY, USA, 1963; Volume 2, pp. 26–77. [Google Scholar]
- Ryther, J.H.; Dunstan, W.M. Nitrogen, phosphorus, and eutrophication in the coastal marine environment. Science 1971, 171, 1008–1031. [Google Scholar] [CrossRef] [Green Version]
- Rhee, G.Y. Effect of N:P atomic ratios and nitrate limitation on algal growth, cell composition and nitrate uptake. Limnol. Oceanogr. 1978, 23, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Tyrerell, T. The relative influences of nitrogen and phosphorus on ocean primary production. Nature 1999, 400, 525–531. [Google Scholar] [CrossRef]
- Glibert, P.M.; Maranger, R.; Sobota, D.J.; Bouwman, L. The Haber Bosch–harmful algal bloom (HB–HAB) link. Environ. Res. Lett. 2014, 9, 105001. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Bi, H.; She, J.; Maar, M.; Jonasson, L. Model study on horizontal variability of nutrient N/P ratio in the Baltic Sea and its impacts on primary production, nitrogen fixation and nutrient limitation. Ocean Sci. Discuss. 2012, 9, 385–419. [Google Scholar] [CrossRef] [Green Version]
- Danielsson, Å.; Papush, L.; Rahm, L. Alterations in nutrient limitations–Scenarios of a changing Baltic Sea. J. Mar. Syst. 2008, 73, 263–283. [Google Scholar] [CrossRef]
- Wu, M.L.; Wang, Y.S.; Wang, Y.T.; Yin, J.P.; Dong, J.D.; Jiang, Z.Y.; Sun, F.L. Scenarios of nutrient alterations and responses of phytoplankton in a changing Daya Bay, South China Sea. J. Mar. Syst. 2017, 165, 1–12. [Google Scholar] [CrossRef]
- Yao, P.; Yu, Z.G.; Deng, C.M.; Liu, S.X.; Zhen, Y. Spatial–temporal distribution of phytoplankton pigments in relation to nutrient status in Jiaozhou Bay, China. Estuar. Coast. Shelf Sci. 2010, 89, 234–244. [Google Scholar] [CrossRef]
- Li, C.L.; Zhang, F.; Yang, X.; Yang, B.; Shen, Z.L.; Sun, S. Concentration, distribution and annual fluctuation of chlorophyll–a in the Jiaozhou Bay. Oceanol. Limnol. Sin. 2005, 36, 499–506, (In Chinese with English abstract). [Google Scholar]
- Li, Y.; Li, R.X.; Wang, Z.L.; Zhu, M.Y.; Sun, P.X.; Xia, B. A preliminary study on phytoplankton community structure and its change in the Jiaozhou Bay. Adv. Mar. Sci. 2005, 23, 320–334, (In Chinese with English abstract). [Google Scholar]
- Sun, X.X.; Sun, S.; Zhang, Y.S.; Zhang, F. Long term changes of chlorophyll–a concentration and primary productivity in the Jiaozhou Bay. Oceanol. Limnol. Sin. 2011, 42, 654–661, (In Chinese with English abstract). [Google Scholar]
- Zhao, S.J.; Jiao, N.Z.; Shen, Z.L.; Wu, Y.L. Causes and consequences of changes in Nutrient Structure in the Jiaozhou Bay. J. Integr. Plant Biol. 2005, 47, 396–410. [Google Scholar] [CrossRef]
- Wang, B.D.; Wang, Z.L. Long–term variations in chlorophyll–a and primary productivity in Jiaozhou Bay, China. J. Mar. Biol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.F.; Gao, Z.H.; Sun, P.Y.; Li, M.; Qu, Y.F. Mechanism of nutrient silicon and water temperature influences on phytoplankton growth. Mar. Sci. Bull. 2006, 8, 49–59. [Google Scholar]
- Liu, S.M.; Zhang, J.; Chen, H.T.; Zhang, G.S. Factors influencing nutrient dynamics in the eutrophic Jiaozhou Bay, North China. Prog. Oceanogr. 2005, 66, 66–85. [Google Scholar] [CrossRef]
- Wu, Y.L.; Sun, S.; Zhang, Y.S. Long–term change of environmental and its influence on phytoplankton community structure in Jiaozhou Bay. Oceanol. Limnol. Sin. 2005, 36, 487–498, (In Chinese with English abstract). [Google Scholar]
- Liu, Z.; Wei, H.; Liu, G.L.; Zhang, J. Simulation of water exchange in Jiaozhou Bay by average residence time approach. Estuar. Coast. Shelf Sci. 2004, 61, 5–35. [Google Scholar] [CrossRef]
- Song, X.X.; Yu, Z.M. Nutrient effect on phytoplankton in typical mariculture waters in summer in the northeast of the Jiaozhou Bay. Oceanol. Limnol. Sin. 2007, 38, 446–452, (In Chinese with English abstract). [Google Scholar]
- Liu, S.M.; Li, X.N.; Zhang, J.; Wei, H.; Ren, J.L.; Zhang, G.L.; Zhang, G.L. Nutrient dynamics in Jiaozhou Bay. Water Air Soil Pollut. 2007, 7, 625–643. [Google Scholar] [CrossRef]
- Liu, Z.; Wei, H.; Bai, J.; Zhang, J.; Liu, D.Y.; Liu, S.M. Nutrients seasonal variation and budget in Jiaozhou Bay, China: A 3–dimensional physical–biological coupled model study. Water Air Soil Pollut. 2007, 7, 607–623. [Google Scholar] [CrossRef]
- Luo, X.; Sun, X.X.; Zheng, S.; Zhao, Y.F. On phytoplankton community structure and causative factors in the Jiaozhou Bay in 2011. Oceanol. Limnol. Sin. 2016, 47, 915–923. [Google Scholar] [CrossRef]
- Liu, H.; Yin, B.S. Numerical investigation of nutrient limitations in the Bohai Sea. Mar. Environ. Res. 2010, 70, 308–317. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.L. Historical changes in nutrient structure and its influences on phytoplankton composition in Jiaozhou Bay. Estuar. Coast. Shelf Sci. 2001, 52, 211–224. [Google Scholar] [CrossRef]
- Liu, D.; Sun, J.; Zhang, J.; Liu, G. Response of the diatom flora in Jiaozhou Bay, China to environmental changes during the last century. Mar. Micropaleontol. 2008, 66, 279–290. [Google Scholar] [CrossRef]
- Liu, D.; Sun, J.; Zou, J.; Zhang, J. Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Mar. Pollut. Bull. 2005, 50, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Ji, R.B.; Zheng, L.Y.; Zhu, M.Y.; Rawson, M. Influences of physical processes on the ecosystem in Jiaozhou Bay: A coupled physical and biological model experiment. J. Geophys. Res.-Oceans. 1999, 104, 29925–29949. [Google Scholar] [CrossRef]
- Shi, H.H.; Shen, C.C.; Li, F.; Wang, Y.Z. Parameter sensitivity analysis of a coupled biological-physical model in Jiaozhou Bay. Acta Ecologica Sin. 2014, 34, 41–49, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Skogen, M.D.; Svendsen, E.; Berntsen, J.; Aksenes, D.; Ulvestad, K.B. Modeling the primary production in the North Sea using a coupled three–dimensional physical–chemical–biological ocean model. Estuar. Coast. Shelf Sci. 1995, 41, 545–565. [Google Scholar] [CrossRef]
- Zhao, L.; Guo, X.Y. Influence of cross–shelf water transport on nutrients and phytoplankton in the East China Sea: A model study. Ocean Sci. 2011, 7, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Pu, X.; Shanghai Nuclear Engineering Research & Design Institute Co, LTD., Shanghai, China. Personal communication, 2011.
- Yang, W.H.; Gao, H.W. Research on numerical model and environmental impact of scallop cultivation in Jiaozhou Bay. Mar. Environ. Sci. 2010, 29, 559–563, (In Chinese with English abstract). [Google Scholar]
- Ren, L.; Zhang, M.P.; Sun, J.; Li, T.; Zhu, C.J. Analysis of some key ecological parameters in the seawater of Jiaozhou Bay. J. Ocean Univ. 2003, 33, 557–564, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.T. Overviews on the nutrients of Jiaozhou Bay. Ocean Sci. 2009, 33, 90–94, (In Chinese with English abstract). [Google Scholar]
- Ren, L.; Zhang, M.P.; Brockmann, U.H.; Feng, S.Z. Pelagic nitrogen cycling in Jiaozhou Bay, a model study Ⅰ: The conceptual model. Chin. J. Oceanol. Limnol. 2003, 21, 358–367. [Google Scholar]
- Editorial Board of Annals of Bays in China. Annals of Bays in China (Series 4); Wang, J.L., Ed.; China Ocean Press: Beijing, China, 1993; pp. 157–169. ISBN 7-5027-2563-6. (In Chinese) [Google Scholar]
- Zhang, J. Watershed nutrient loss and eutrophication of marine recipients: A case study of the Jiaozhou Bay. Water Air Soil Pollut. 2007, 7, 583–592. [Google Scholar] [CrossRef]
- Shi, J.H.; Li, G.X.; Wang, P. Anthropogenic influence on the tidal prism and water exchanges in Jiaozhou Bay, Qingdao, China. J. Coast. Res. 2011, 27, 57–72. [Google Scholar] [CrossRef]
- Yao, P.; Yu, Z.G.; Deng, C.M.; Liu, S.X.; Zhen, Y. Functional types of phytoplankton in Jiaozhou Bay using diagnostic pigment analysis (DPA). Oceanol. Limnol. Sin. 2013, 44, 866–876, (in Chinese with English abstract). [Google Scholar]
- Du, S.; Huang, S.; Zang, C.; Wu, M.; Gao, F.; Lin, C.; Guo, Y.; Luo, Y. Correlation research between the indicators of phytoplankton standing stock Ⅱ: Chlorophyll a and algal density. J. Water Resour. Water Eng. 2011, 22, 44–49, (In Chinese with English abstract). [Google Scholar]
- Sun, S.; Liu, G.M.; Zhang, Y.S. Community composition and distribution character of phytoplankton in the Jiaozhou Bay in the 1990s. Oceanol. Limnol. Sin. 2002, 33, 37–44, (in Chinese with English abstract). [Google Scholar]
- Wu, Y.L.; Sun, S.; Zhang, Y.S.; Zhang, F. Quantitative study on long–term variation of phytoplankton in Jiaozhou Bay. Oceanol. Limnol. Sin. 2004, 35, 518–523, (In Chinese with English abstract). [Google Scholar]
- Zhao, S.J.; Jiao, N.Z.; Wu, C.W.; Liang, B.; Zhang, S.Y. Evolution of nutrient structure and phytoplankton composition in the Jiaozhou Bay ecosystem. J. Environ. Sci. 2005, 17, 95–102. [Google Scholar]
- Li, M.; Xu, K.; Watanabe, M.; Chen, Z. Long–term variations in dissolved silicate, nitrogen, and phosphorus flux from the Yangtze river into the east China sea and impacts on estuarine ecosystem. Estuar. Coast. Shelf Sci. 2007, 71, 3–12. [Google Scholar] [CrossRef]
- Liu, M.; Zhi, C.Y.; Li, K. Mechanism and ecological effect of river–transported silicon flux reduction. Ecol. Sci. 2010, 29, 171–175, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Domingues, R.B.; Barbosa, A.B.; Galvão, H.M. River damming leads to decreased phytoplankton biomass and disappearance of cyanobacteria blooms. Estuar. Coast. Shelf Sci. 2014, 136, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Conley, D.; Stålnacke, P.; Pitkänen, H.; Wilander, A. The transport and retention of dissolved silicate by rivers in Sweden and Finland. Limnol. Oceanogr. 2000, 45, 1850–1853. [Google Scholar] [CrossRef]
- Wu, G.J.; Cao, W.Z.; Wang, F.F.; Su, X.L.; Yan, Y.Y.; Guan, Q.S. Riverine nutrient fluxes and environmental effects on China’s estuaries. Sci. Total Environ. 2019, 661, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Garnier, J.; Beusen, A.; Thieu, V.; Billen, G.; Bouwman, L. N:P:Si nutrient export ratios and ecological consequences in coastal seas evaluated by the icep approach. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Ma, J.; Zhou, B.B.; Duan, D.D.; Wei, Y.; Pan, K. Silicon limitation reduced the adsorption of cadmium in marine diatoms. Aquat. Toxicol. 2018, 202, 136–144. [Google Scholar] [CrossRef]
- Wang, W.X.; Dei, R.C.H. Metal uptake in a coastal diatom influenced by major nutrients (N, P, and Si). Water Res. 2001, 35, 315–321. [Google Scholar] [CrossRef]
- Burson, A.; Stomp, M.; Akil, L.; Brussaard, C.P.D.; Huisman, J. Unbalanced reduction of nutrient loads has created an offshore gradient from phosphorus to nitrogen limitation in the North Sea. Limnol. Oceanogr. 2016, 61, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, G.T.; Wan, A.Y.; Zhao, Z.X.; Wang, S.W.; Liu, Q. Nutrient–limitation induced diatom–dinoflagellate shift of spring phytoplankton community in an offshore shellfish farming area. Mar. Pollut. Bull. 2019, 141, 1–8. [Google Scholar] [CrossRef]
- Sugiura, S.H. Phosphorus, Aquaculture, and the Environment. Rev. Fish. Sci. Aquac. 2018, 26, 515–521. [Google Scholar] [CrossRef]
- Kumararaja, P.; Suvana, S.; Saraswathy, R.; Lalitha, N.; Muralidhar, M. Mitigation of eutrophication through phosphate removal by aluminium T pillared bentonite from aquaculture discharge water. Ocean Coast. Manage. 2019, 182, 104951. [Google Scholar] [CrossRef]
- Fisher, T.R.; Hagy, J.D.; Boynton, W.R.; Williams, M.R. Cultural eutrophication in the Choptank and Patuxent estuaries of Chesapeak Bay. Limnol. Oceanogr. 2006, 5, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Saeck, E.A.; O’Brien, K.R.; Weber, T.R.; Burford, M.A. Changes to chronic nitrogen loading from sewage discharges modify standing stocks of coastal phytoplankton. Mar. Pollut. Bull. 2013, 71, 159–167. [Google Scholar] [CrossRef]
- Gypens, N.; Delhez, E.; Brunier, A.V.; Vanhoutte–Brunier, A.; Burton, S.; Thieu, V.; Passy, P.; Liu, Y.; Callens, J.; Rousseau, V.; et al. Modelling phytoplankton succession and nutrient transfer along the Scheldt estuary (Belgium, The Netherlands). J. Mar. Syst. 2012, 128, 89–105. [Google Scholar] [CrossRef]
- Taylor, D.I.; Candace, C.A.; Giblin, A.E.; Tucker, J.; Diaz, R.J.; Keay, K. Wastewater input reductions reverse historic hypereutrophication of Boston Harbor, USA. Ambio 2020, 49, 187–196. [Google Scholar] [CrossRef]
- Oviatt, C.; Smith, L.; Krumholz, J.; Coupland, C.; Stoffel, H.; Keller, A.; McManus, M.C.; Reed, L. Managed nutrient reduction impacts on nutrient concentrations, water clarity, primary production, and hypoxia in a north temperate estuary. Estuar. Coast. Shelf Sci. 2017, 199, 25–34. [Google Scholar] [CrossRef]
- Zhang, G.S.; Zhang, J.; Liu, S.M. Characterization of nutrients in the atmospheric wet and dry deposition observed at the two monitoring sites over yellow sea and East China Sea. J. Atmos. Chem. 2007, 57, 41–57. [Google Scholar] [CrossRef]
- Duce, R.A.; Laroche, J.; Altieri, K.; Arrigo, K.R.; Baker, A.R.; Cornell, S.; Dentener, F.; Galloway, J.; Ganeshram, R.S.; Geider, R.J.; et al. Impacts of Atmospheric Anthropogenic Nitrogen on the Open Ocean. Science 2008, 320, 893–897. [Google Scholar] [CrossRef] [Green Version]
- Martino, M.; Hamilton, D.; Baker, A.R.; Jickells, T.D.; Bromley, T.; Nojiri, Y.; Quack, B.; Boyd, P.W. Western pacific atmospheric nutrient deposition fluxes, their impact on surface ocean productivity. Glob. Biogeochem. Cycles 2014, 28, 712–728. [Google Scholar] [CrossRef]
- Guo, W.; Shi, J.; Zhu, J.Y.; Zhao, L. Modelling the Effects of the Atmospheric Nitrogen Deposition on the Primary Production Process in the Southern Yellow Sea. J. Ocean Univ. 2020, 50, 115–125, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Qi, J.H.; Liu, X.H.; Yao, X.H.; Zhang, R.F.; Chen, X.J.; Lin, X.H.; Gao, H.W.; Liu, R.H. The concentration, source and deposition flux of ammonium and nitrate in atmospheric particles during dust events at a coastal site in northern China. Atmos. Chem. Phys. 2018, 18, 571–586. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.W.; Song, J.M.; Yuan, H.M.; Li, X.G.; Li, N.; Duan, L.Q.; Kang, X.M.; Wang, Q.D. Fluxes, seasonal patterns and sources of various nutrient species (nitrogen, phosphorus and silicon) in atmospheric wet deposition and their ecological effects on Jiaozhou Bay, North China. Sci. Total Environ. 2017, 576, 617–627. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, X.H.; Wu, Q.S.; Huang, D.J. The Kuroshio nutrient stream and its temporal variation in the East China Sea. J. Geophys. Res.Oceans. 2012, 117, C01026:1–C01026:17. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhu, X.H.; Yu, L.; Huang, D.J. Spatial variations in the Kuroshio nutrient transport from the East China Sea to south of Japan. Biogeosciences 2013, 10, 6403–6417. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Unit | Description of Parameter |
|---|---|---|---|
| 1.3 | Maximum growth rate of DIA | ||
| 0.3 | Maximum growth rate of FLA | ||
| 0.138 | - | Percentage of basic respiration | |
| 0.05 | - | Percentage of photorespiration | |
| 0.12 | Phytoplankton mortality rate | ||
| 0.055 | Temperature-dependent growth rate | ||
| 0.054 | Temperature-dependent respiration rate | ||
| 120 | Optimum light intensity of DIA | ||
| 70 | Optimum light intensity of FLA | ||
| 1.4 | DIN half saturation constant for DIA | ||
| 0.12 | DIP half saturation constant for DIA | ||
| 2.4 | SIL half saturation constant for DIA | ||
| 1.2 | DIN half saturation constant for FLA | ||
| 0.1 | DIP half saturation constant for FLA | ||
| 0.1 | Mineralization rate of DIA detritus | ||
| 0.15 | Mineralization rate of FLA detritus | ||
| N/P/Si | 16/1/22 | - | Atomic ratio of N, P, and Si |
| 1960s | 1980s | 2000s | Unit | Reference | |
|---|---|---|---|---|---|
| Dagu River runoff (F) | 7.5 | 5.0 | 3.0 | 1011 L y−1 | [38,41] |
| Riverine DIN concentration () | 60 | 260 | 400 | [23,41] | |
| Riverine DIP concentration () | 3 | 5.3 | 7.0 | [23,41] | |
| Riverine SIL concentration () | 600 | 420 | 150 | [23,41] | |
| DIN from wastewater () | 0 | 80 | 200 | 106 | [23,26] |
| DIP from wastewater () | 0 | 0.8 | 2.0 | 106 | [23,26] |
| SIL from wastewater () | 0 | 3.2 | 8.0 | 106 | [23,26] |
| DIN from aquaculture () | 0 | 1.8 | 54 | 106 | [24,39] |
| DIP from aquaculture () | 0 | 0.12 | 3.8 | 106 | [24,40] |
| DIN from atmospheric deposition () | 56 | 56 | 56 | 106 | [27] |
| DIP from atmospheric deposition () | 0.42 | 0.42 | 0.42 | 106 | [27] |
| SIL from atmospheric deposition () | 0.93 | 0.93 | 0.93 | 106 | [27] |
| DIN concentration of YS () | 2.2 | 5.0 | 6.0 | [18,40] | |
| DIP concentration of YS () | 0.17 | 0.3 | 0.3 | [18,40] | |
| SIL concentration of YS () | 6.0 | 5.5 | 4.0 | [18,40] |
| Parameter | Variation of Parameter (%) | Sensitivities of Annual DIA Biomass | Sensitivities of Annual FLA Biomass |
|---|---|---|---|
| +50/−50 | 0.71/0.93 | 0.13/0.12 | |
| +50/−50 | 0/0 | 0.99/0.99 | |
| +50/−50 | 0.34/0.34 | 0.22/0.22 | |
| +50/−50 | 0.06/0.06 | 0.06/0.06 | |
| +50/−50 | 0.62/1.66 | 0.65/1.91 | |
| +50/−50 | 0.54/0.63 | 0.11/0.12 | |
| +50/−50 | 0/0 | 0.31/0.17 | |
| +50/−50 | 0/0 | 0/0 | |
| +50/−50 | 0/0 | 0/0 | |
| +50/−50 | 0.31/0.39 | 0/0.01 | |
| +50/−50 | 0/0 | 0/0 | |
| +50/−50 | 0/0 | 0.17/0.20 | |
| +50/−50 | 0.05/0.14 | 0/0.02 | |
| +50/−50 | 0/0 | 0/0 |
| DIA Photosynthesis | DIA Respiration | FLA Photosynthesis | FLA Respiration | DSi Mineralization | DNSi Mineralization | Exchange with YS | ||
|---|---|---|---|---|---|---|---|---|
| CONTROL | DIN | −395.83 | +117.75 | −49.91 | +11.10 | +278.37 | +38.82 | −432.09 |
| DIP | −24.74 | +7.36 | −3.12 | +0.69 | +17.40 | +2.43 | −8.36 | |
| SIL | −544.27 | +161.90 | 0 | 0 | +382.76 | 0 | −54.70 | |
| NORI | DIN | −266.29 | +91.83 | −46.50 | +10.61 | +174.62 | +35.90 | −309.99 |
| DIP | −16.64 | +5.74 | −2.90 | +0.66 | +10.92 | +2.24 | −6.22 | |
| SIL | −366.15 | +126.25 | 0 | 0 | +240.10 | 0 | −8.92 | |
| NOWA | DIN | −384.07 | +115.40 | −47.81 | +10.79 | +268.96 | +37.03 | −232.09 |
| DIP | −24.00 | +7.21 | −2.98 | +0.67 | +16.81 | +2.31 | −6.36 | |
| SIL | −528.10 | +158.69 | 0 | 0 | +369.82 | 0 | −46.70 | |
| NOAQ | DIN | −395.83 | +117.75 | −45.40 | +10.44 | +278.37 | +35.00 | −378.09 |
| DIP | −24.74 | +7.36 | −2.84 | +0.65 | +17.40 | +2.19 | −4.56 | |
| SIL | −544.27 | +161.90 | 0 | 0 | +382.76 | 0 | −54.70 | |
| NOAT | DIN | −394.49 | +117.48 | −49.49 | +11.04 | +277.29 | +38.47 | −376.09 |
| DIP | −24.66 | +7.34 | −3.09 | +0.69 | +17.33 | +2.40 | −7.94 | |
| SIL | −542.42 | +161.54 | 0 | 0 | +381.29 | 0 | −53.77 |
| 1960s | 1980s | 2000s | ||||
|---|---|---|---|---|---|---|
| Simulation | Observation | Simulation | Observation | Simulation | Observation | |
| DIN | 0.98–5.43 | 1.00–4.00 | 12.05–23.65 | 2.55–19.50 | 17.24–37.74 | 3.60–44.21 |
| DIP | 0.18–0.46 | 0.10–0.22 | 0.08–0.52 | 0.20–0.72 | 0.09–1.37 | 0.09–2.00 |
| SIL | 8.38–62.83 | - | 5.59–27.66 | 0.20–11.50 | 2.44–12.51 | 1.10–20.39 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Leng, Q.; Zhu, J.; Gao, H.; Guo, X.; Mao, X. Influences of Nutrient Sources on the Alternation of Nutrient Limitations and Phytoplankton Community in Jiaozhou Bay, Southern Yellow Sea of China. Sustainability 2020, 12, 2224. https://0-doi-org.brum.beds.ac.uk/10.3390/su12062224
Shi J, Leng Q, Zhu J, Gao H, Guo X, Mao X. Influences of Nutrient Sources on the Alternation of Nutrient Limitations and Phytoplankton Community in Jiaozhou Bay, Southern Yellow Sea of China. Sustainability. 2020; 12(6):2224. https://0-doi-org.brum.beds.ac.uk/10.3390/su12062224
Chicago/Turabian StyleShi, Jie, Qian Leng, Junying Zhu, Huiwang Gao, Xinyu Guo, and Xinyan Mao. 2020. "Influences of Nutrient Sources on the Alternation of Nutrient Limitations and Phytoplankton Community in Jiaozhou Bay, Southern Yellow Sea of China" Sustainability 12, no. 6: 2224. https://0-doi-org.brum.beds.ac.uk/10.3390/su12062224