1. Introduction
Dinoflagellates are ubiquitous protists that can be found in almost every conceivable marine environment [
1]. Recently, they have drawn increasing attention from both the scientific and public communities because of the following reasons: Some dinoflagellates form red tides and/or harmful algal blooms, which often cause large-scale mortality of fish [
2], they play indispensable ecological roles in marine plankton communities, serving as prey for a variety of predators and as a predator on diverse microorganisms [
3], some dinoflagellates are used as various biological resources and they are also treated as one of the most promising bio-resources for new high-value products [
4,
5,
6]. In particular, dinoflagellates are known to be rich in long-chain omega-3 PUFAs, such as EPA (C
20:5 n-3) and DHA (C
22:6 n-3) [
7,
8,
9]. Thus, the amount and composition of omega-3 PUFAs from dinoflagellates in the phytoplankton community are strongly intertwined with higher trophic levels, eventually affecting the nutritional values of higher-order consumers [
7,
10,
11]. Therefore, to better understand the role of dinoflagellates in the marine ecosystems and their potential commercial applications, the establishment of a clonal culture is a critical first step for accurate identification of an isolate of interest.
The family Symbiodiniaceae (Order Suessiales) comprises symbiotic dinoflagellates, most of which are symbiotic with invertebrate and protist hosts [
12,
13,
14,
15,
16,
17], even though some species exist as free-living forms [
18]. These dinoflagellates are vital components of the coral reef ecosystems, and they are promising resources in the production of valuable pigments [
19] and toxin compounds, such as zooxanthella toxins [
20]. Despite their ecological and economic importance in the marine ecosystems and biotechnology, little information is available on their taxonomy. This is due to, in part, the difficulties in culturing and making morphological observations, which must be done by scanning electron and transmission electron microscopy (SEM and TEM). Therefore, their roles as essential components of the marine ecosystems and their potential for biotechnology are often overlooked. Accurate identification and establishment of a clonal culture of these dinoflagellates are essential for further research and commercial application.
The systematics of Symbiodiniaceae was revised, and distinct clades within the family were reassigned into seven genera in 2018 [
21].
Symbiodinium voratum was first described and named by Jeong et al. [
18], it was renamed as
Effrenium voratum by LaJeunesse et al. [
21]. This is the only taxonomically accepted species in the genus
Effrenium [
21].
E. voratum is found in the Pacific and Atlantic Oceans, and it grows at sub-tropical and temperate latitudes [
18,
21,
22,
23,
24]. Jeong et al. [
18] reported the Korean
E. voratum strains that had been isolated from waters off Jeju Island. However, no additional documentation of
E. voratum colonies in Korean water have been reported.
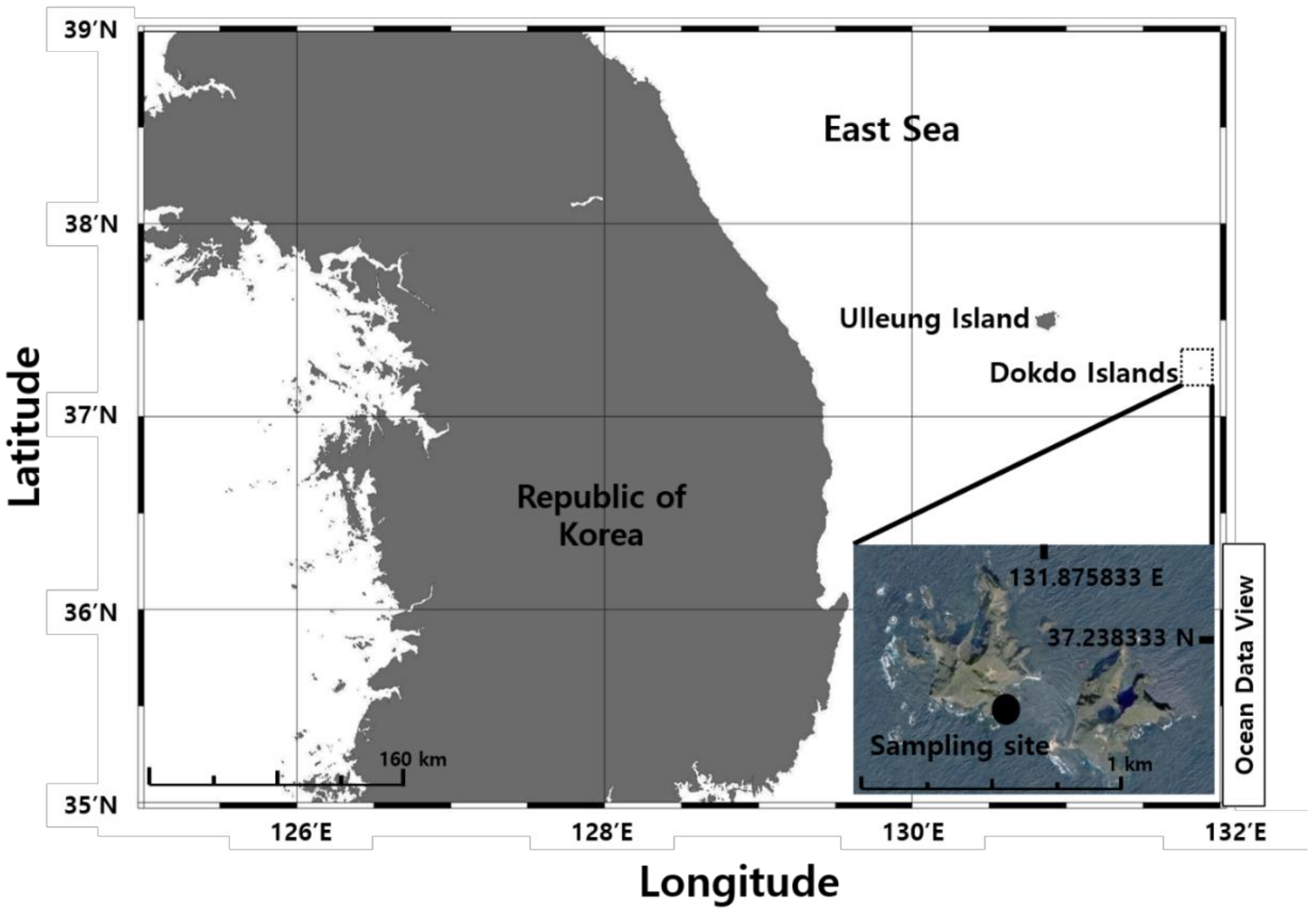
In this study, we isolated and identified a unicellular dinoflagellate E. voratum from seawater off the Dokdo Islands, Dokdo-ri, Ulleung-eup, Ulleung-gun, Gyeongsangbuk-do, Korea. The present report provides information about the morphological, molecular, and chemotaxonomic features on the first record of this species living in the East Sea of Korea.
3. Results
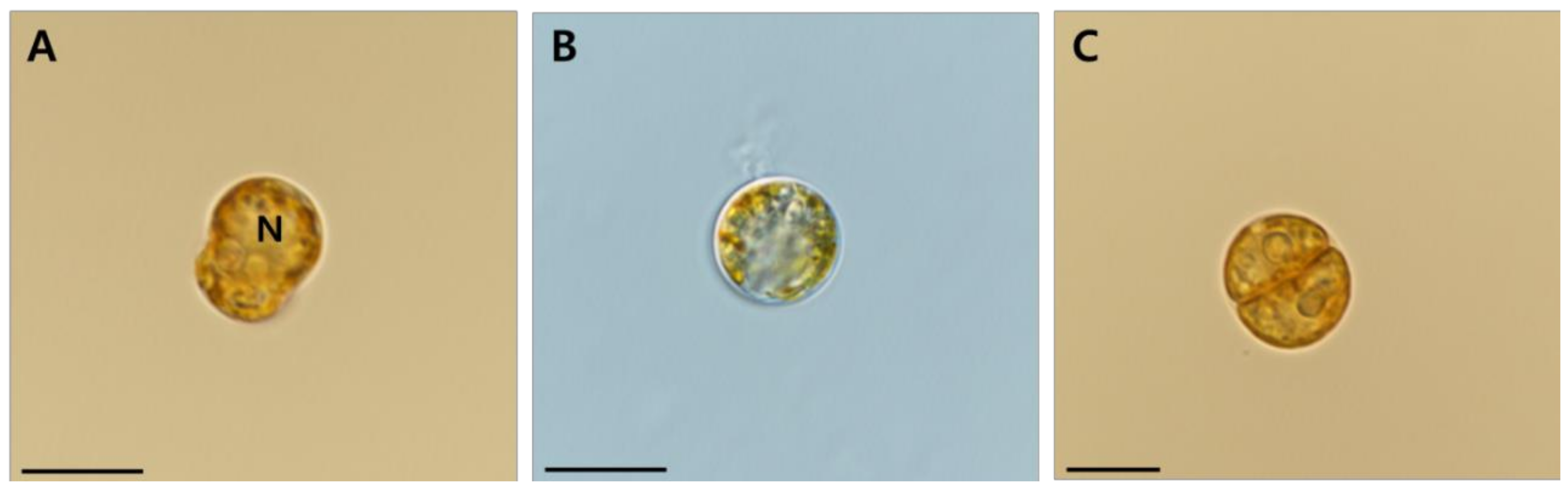
Mastigote, coccoid, and doublet cells occurred in all cultures examined under light microscopy, and they appeared to be like one another (
Figure 2A–C). Motile cells of
E.
voratum were mushroomed-shaped with the hemispherical episome slightly larger than the hemispherical hyposome (
Figure 2A). The nucleus was in the episome (
Figure 2A). The ranges (mean ± standard error, n = 30) of the living cell length and width were 9.42–15.6 μm (12 ± 0.2) and 7.08–11.9 μm (9.7 ± 0.2), respectively (
Table 3). The ratios of the cell length to the width (mean ± standard error, n = 30) ranged from 1.12 to 1.4 (1.2 ± 0.01). When observed under SEM, fixed cells were slightly smaller than unfixed cells. The cell length under SEM had a range of 7.3–12.7 μm (9.6 ± 0.2) and the width had a range of 5.52–11.4 μm (7.7 ± 0.2), respectively (
Table 3). The ratio of the length to the width under SEM had a range of 0.83–1.4 μm (1.2 ± 0.02) (
Table 3).
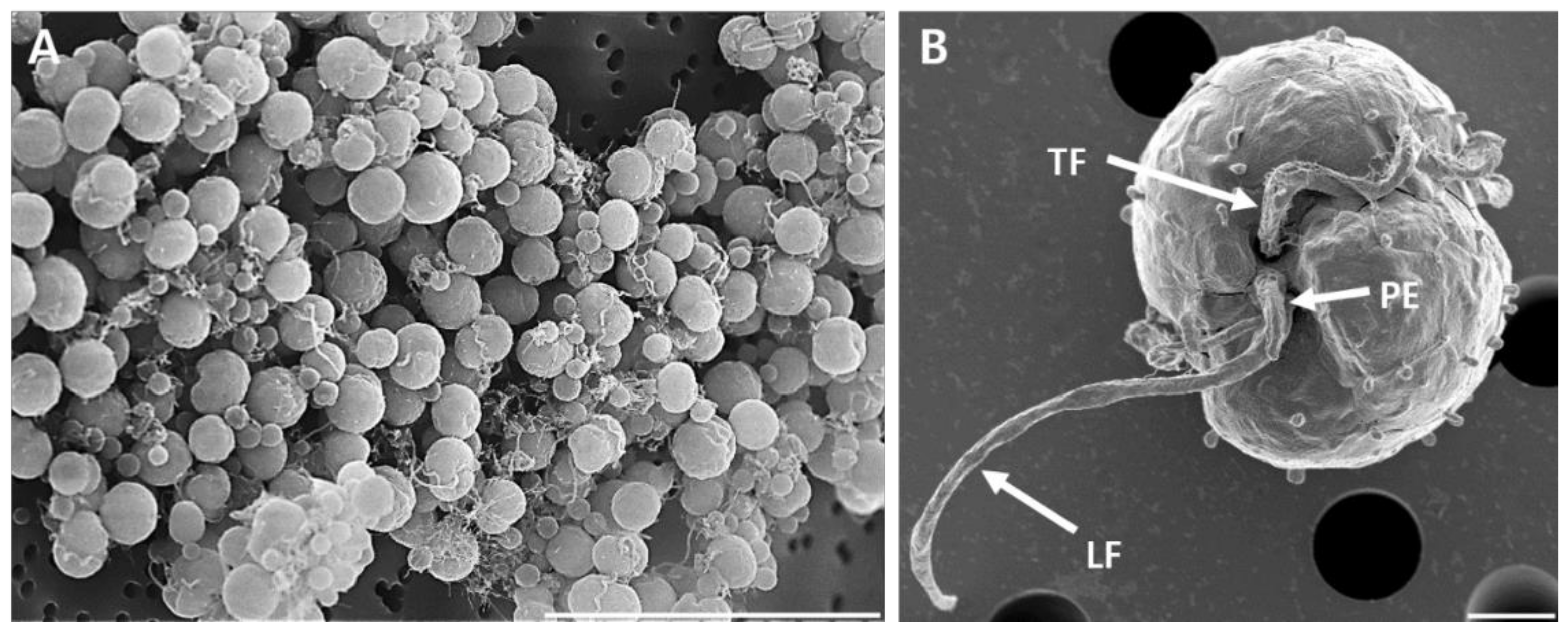
Mastigote, coccoid, and doublet cells were present in all cultures under SEM observation (
Figure 3 and
Figure 4). Mastigotes possessed a well-formed peduncle (PE) extending near the base of the longitudinal (LF) and transverse flagella (TF) (
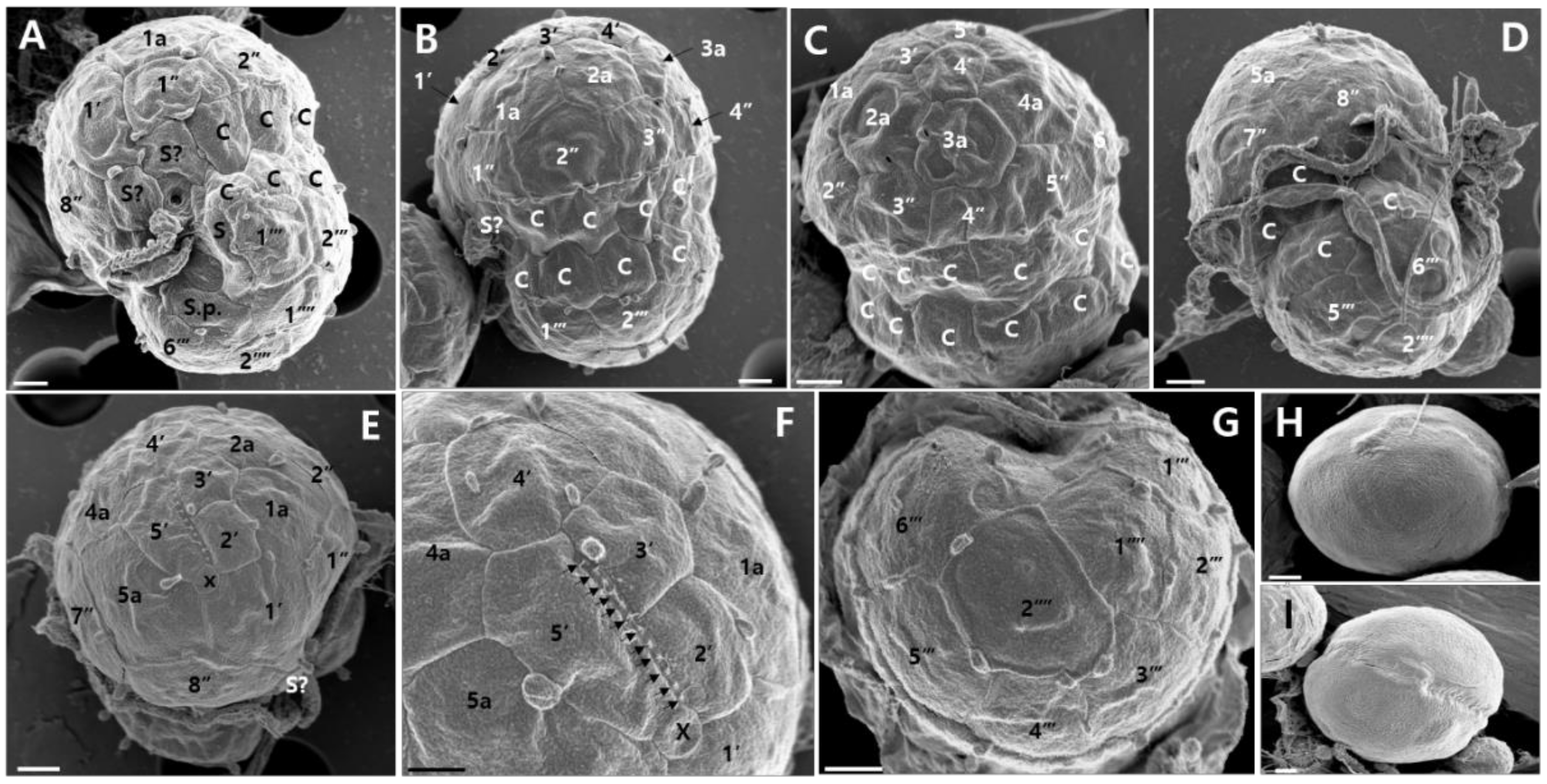
Figure 3B). The Kofoidian plate formula of
E.
voratum cells was x, elongated amphiesmal vesicle (EAV), 5′, 5a, 8′′, 9s, two cingulum rows 17-20c, 6′′′, and 2′′′′ (
Table 3,
Figure 4). At the cell’s apex, the EAV possessed 11–12 aligned knobs, and the length had a range of 1.71–2.72 μm (2.25 ± 0.11) and the width had a range of 0.15–0.26 μm (0.21 ± 0.01) (
Table 3). This structure was bordered ventrally, by the x plate and surrounded by four apical amphiesmal plates (2′, 3′, 4′, and 5′ plates;
Figure 4E,F). The rhomboid-shaped 1′ plate was relatively large and the quadrangular 2′ plate was relatively small (
Figure 4E). The pentagonal 3′ plate touched the 2′, 4′, 1a, and 2a plates and the pentagonal 4′ plate touched the 3′, 5′, 2a, 3a, and 4a (
Figure 4C,E,F). The pentagonal 5′ plate touched the x, 4′, 4a, and 5a plates (
Figure 4C,E,F). Eight pre-cingular plates were present (
Figure 4A–E). The 1′′, 4′′, and 6′′ plates were quadrangular, while the 2′′, 3′′, 5′′, 7′′, and 8′′ were pentagonal. In addition, five intercalary plates were observed (
Figure 4A–F). Cells of
E. voratum had a wide cingulum comprising of two rows of pentagonal amphiesmal plates (
Figure 4A–D). Cells contained 17–20 cingular plates (
Table 3). The cingulum of
E. voratum was displaced by ~0.1–0.3 times the cell length and by ~0.5–0.8 times the cingular width (
Table 3,
Figure 4A). The cells of
E. voratum contained six post-cingular plates (
Figure 4G). Except for the pentagonal 3″′ plate, all post-cingular plates were quadrangular (
Figure 4G). Two antapical plates were present in the cells of
E. voratum (
Figure 4G). The 2′′′′ plates in most cells were hexagonal, and they touched the 3′′′, 4′′′, 5′′′, 6′′′, and 1′′′′ plates and the posterior sulcal plate (S.p.) (
Figure 4A,G).
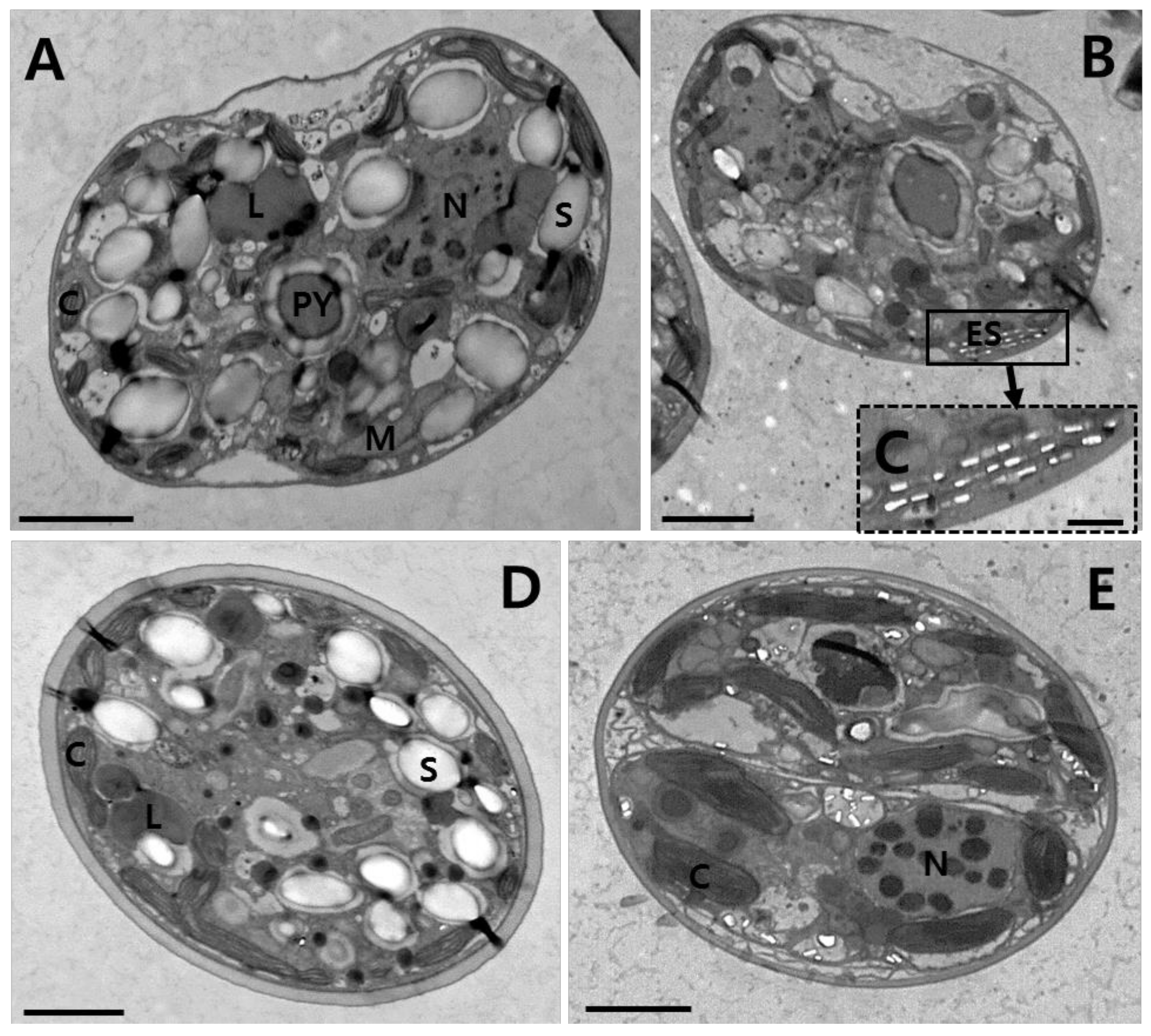
Mastigote, coccoid, and doublet cells were observed in all cultures under TEM (
Figure 5). Thin sections for TEM showed the main features of the cell, including the chloroplasts (C), eyespot (ES), lipids (L), mitochondria (M), nucleus (N), pyrenoids (PY), and starch (S) (
Figure 5). Many chloroplasts were observed along the cell periphery (
Figure 5A,B). A single pyrenoid located toward the central part of each cell was connected by two stalks to the adjacent chloroplast, and surrounded by a distinct polysaccharide cap (
Figure 5A,B). Eyespots composed of several flattened vesicles or cisternae containing electron translucent brick-shaped structures were observed near the surface of the cell (
Figure 5B,C). An eyespot (type E), composed of multiple layers of rectangular electron-translucent vesicles or crystalline deposits, was observed in sectioned mastigote cells (
Figure 5B,C).
When properly aligned, the sequence of LSU rDNA of
E.
voratum MABIKLP88 (NCBI GenBank accession number MN904916, Dokdo Island, Korea), was identical to
E.
voratum strains SVFL 1 (Jeju Island, Korea), TSP-C2-Sy (Tsushima Island, Japan), and CCMP 421 (Cook Strait, New Zealand); while
E.
voratum strains RCC 1521 (Blanes, Spain) and rt-383 (Santa Barbara, CA, USA) have a single-base substitution in LSU compared to strain MABIKLP88 (
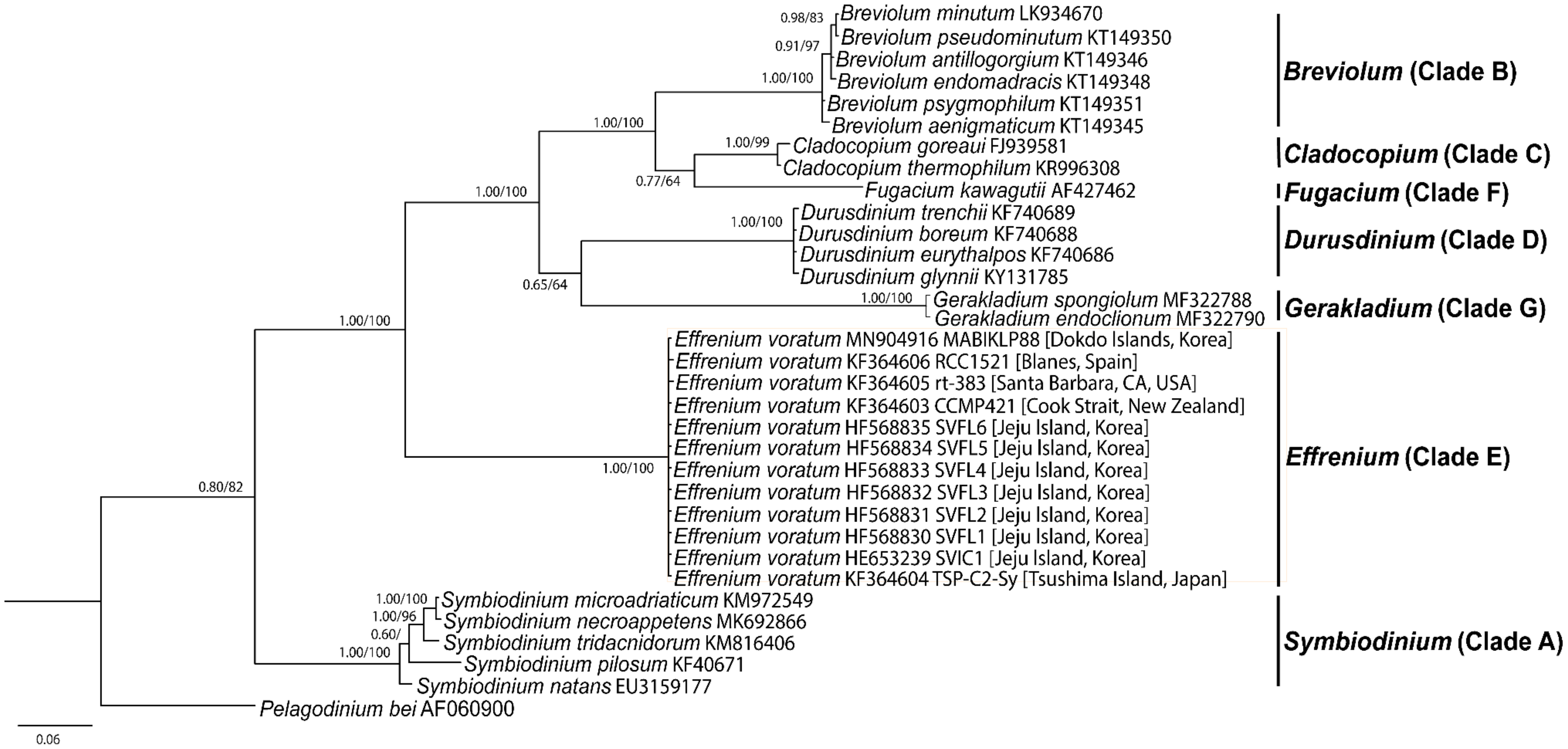
Table 4). In the phylogenetic tree based on the LSU rDNA sequences, the phylogenetic diversity was presently divided into seven distinct monophyletic groups (clades A-G). The
Effrenium clade E was phylogenetically basal of clade B, C, D, F, and G (
Figure 6). In addition,
E.
voratum strain MABIKLP88 formed a big clade (i.e.,
Effrenium clade E) with strains SVIC1, SVFL1-6, TSP-C2-Sy, CCMP 421, RCC 1521, and rt-383 (
Figure 6).
The major FAME profile of the isolate was C
16:0 (22.1%), C
18:4 n-3 (15.2%), C
20:5 n-3 (10.9%), and C
22:6 n-3 (25.4%) (
Table 5). Trace amounts of saturated fatty acids (SFAs, C
12:0, 0.6%; C
14:0, 3.5%; C
15:0, 0.3%; C
18:0, 0.7%), unsaturated fatty acids (C
16:1 n-7, 9.3%; C
16:2 n-4, 0.4%; C
18:1 n-9, 3.3%; C
18:2 n-6, 0.6%; C
18:3 n-6, 0.9%; C
18:3 n-3, 0.3%), and unidentified fatty acids (6.5%), were detected (
Table 5).
4. Discussion
The key for differentiating families in the order Suessiales [
36] is the morphology of the apical furrow: Species in the family Tovelliaceae have an apical line of narrow plates (ALP), species in the family Borghiellaceae have a pair of elongated anterior vesicles (PEV), and species in the family Symbiodinaceae have a single EAV [
37]. All Symbiodinaceae contain a type E eyespot next to the sulcus groove of the motile cell. The motile stage (mastigote) has only seven latitudinal series of amphiesmal vesicles [
21]. Strain MABIKLP88 had the EAV, type E eyespot, and seven latitudinal series of amphiesmal vesicles, these morphologies fit the criteria for the family Symbiodinaceae.
The morphological features of this isolate resembled those of the strain SvFL 1 and CCMP421 because it had a type E eyespot, a peduncle, two-stalked pyrenoid, peripheral chloroplasts, and a Kofoidian series of the small plate (x), the EAV, 5′, 5a, 8′′, 9s, 17-20c, 6′′′, and 2′′′′ (
Table 3). The length and the width of living
E.
voratum MABIKLP88 cells were 9.42–15.6 μm and 7.08–11.9 μm, respectively. The sequence of the D1/D2 region of LSU rDNA was also identical to those of strains SvFL 1 and CCMP421 (
Table 4). Although these
E.
voratum strains were isolated from distinct sites (the Dokdo Islands in Korea, Jeju Island in Korea, and Cook Strait in New Zealand), they all share the typical morphologies of
E.
voratum and identical LSU rDNA sequences. Thus, this newly isolated Korean strain could serve as a good example of a cosmopolite species found in different oceans. The length and width of living cells of the
E.
voratum MABIKLP88 (9.42-15.6 μm and 7.08-11.9 μm, respectively) were comparable to those of other strains of
E.
voratum reported at 10.1–17.1 μm and 7.8–14.4 μm, respectively (
Table 3). In SEM micrographs of cells, the width and EAV length of the
E.
voratum MABIKLP88 (5.52–11.4 μm and 1.71–2.72 μm, respectively) were also similar to those of other strains of
E.
voratum (5.8–10.9 μm and 1.75–3.19 μm, respectively) (
Table 3). The ratio of cell length to the width of the
E.
voratum MABIKLP88 (0.83–1.4) was comparable to those of other strains of
E.
voratum (1.2–1.4) (
Table 3). The ratio of cingulum displacement to cell length of the
E.
voratum MABIKLP88 (0.11–0.26) was similar to those of other strains of
E.
voratum (0.1–0.21) (
Table 3). Therefore, our report provides new information on the extended ranges of cell length, cell width, EAV length, the ratio of cell length to cell width, and the ratio of cingulum displacement to cell length in
E.
voratum. These results showed more variability on the measured morphological traits of the species than previously reported
E.
voratum strains.
The sequence of the D1/D2 region of LSU rDNA was identical to the of the
E.
voratum strains located in the waters off the Jeju Island of Korea, Tsushima Island of Japan, and Cook Strait of New Zealand. Our phylogenetic analysis confirmed that the strain MABIKLP88 belonged to
E.
voratum (
Table 4,
Figure 6). Molecular phylogeny results from the LSU rDNA sequences supported the morphological identification by forming a well-supported clade, including the sequences from the original description.
Prior to the present study,
E.
voratum had been reported to live in waters in the temperate latitudes in the western North Pacific, the southwest Western Pacific, the eastern North Pacific, the eastern Atlantic, and the Mediterranean Sea [
18,
21,
22,
23,
24,
38,
39,
40].
E.
voratum appears to be distributed across high sub-tropical and low temperate latitudes.
E.
voratum is cold-water-adapted and may tolerate temperatures as low as 10–12 °C. This is supported by physiological measurements taken on culture rt-383, which showed that this strain grew optimally at 15–20 °C, but also grew at 12 °C and 28 °C [
41]. This wide temperature range tolerance may enable
E.
voratum to survive in a wide variety of oceanic environments around the world (
Figure 7). Additional physiological tests on rt-383 and other
E.
voratum strains are needed to elucidate their thermal tolerance. In addition, the maximal growth of strain MABIKLP88 was obtained at 22–28 °C and 300–700 μmol photons m
−2 s
−1 (unpublished data). Moreover,
E.
voratum has an ability to feed on bacteria and other macroalgae, proving a possible survival strategy for
Effrenium to persist in nutrient-poor conditions [
39].
It seems that this wide temperature range tolerance of the species, coupled with tolerance to high light intensities and mixotrophic ability, attributed to E. voratum’s survival and presence in a wide variety of oceanic environments around the world. Thus, this newly isolated Korean strain could serve as a good example of a candidate for cosmopolite species in high sub-tropical and low temperate latitudes. Exploration of this dinoflagellate’s distribution in greater detail would be worthwhile.
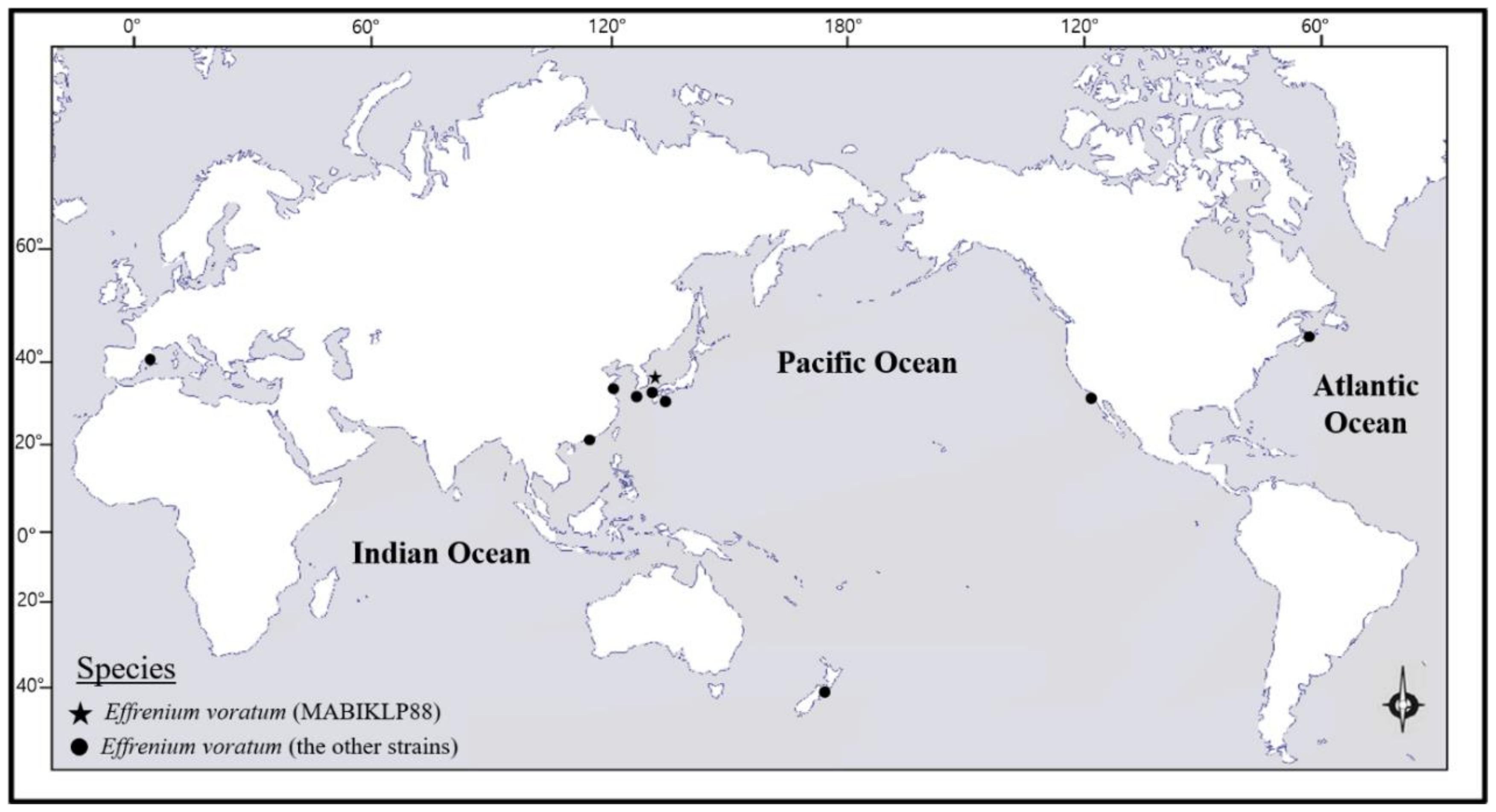
As shown in
Figure 7,
E.
voratum strains are present in many locations, and the current report further describes the occurrence of
E.
voratum in the western North Pacific Ocean (
Table 6,
Figure 7). The latitude of the Dokdo Islands, where strain MABIKLP88 was isolated, is 37.2 N. The locations where additional stains of the western North Pacific have been reported range from 22.5 N to 36.0 N (
Table 6). To date, the waters around the Dokdo Islands are the highest latitude in the western North Pacific, where
E.
voratum has been found. The findings in this report expand upon the understanding of the geographic distribution of
E.
voratum in the western North Pacific. It may also show that climate changes caused by global warming have taken place in the oceans around the Korean Peninsula since the Dokdo Islands are located at the front between the warm Kuroshio and the cold Kuril currents.
Analysis of the cellular fatty acid composition of the strain MABIKLP88 revealed that it was rich in C
16:0 (22.1%), SFA and C
18:4 n-3 (15.2%), C
20:5 n-3 (10.9%), and C
22:6 n-3 (25.4%) PUFAs. The fatty acid profile of different dinoflagellate species has been extensively studied in order to find valuable strains with high potentials for commercial applications [
7,
8,
42,
43,
44,
45,
46]. The profile has also been used as a general chemotaxonomic guide to defining a variety of taxonomic groups since different microalgal groups show distinct fatty acid distributions [
44,
47,
48]. In particular, the C
18 fatty acids and > C
20 PUFAs are generally regarded as the signature fatty acids for dinoflagellates [
8,
48]. In this study,
E.
voratum MABIKLP88 was found to be rich in the C
18 fatty acid, EPA, and DHA. It has also been reported that dinoflagellates typically have higher DHA than EPA concentrations [
7,
8,
9], and the isolate exhibited much higher DHA (25.4%) content than EPA (10.9%). Several previous studies have shown that the essential omega-3 PUFAs have a variety of beneficial health effects [
49]. Importantly, EPA and DHA, which are known essential omega-3 fatty acids, have been reported to be beneficial to human health [
50,
51]. Most of the omega-3 PUFAs come from marine sources, such as fish oils and a variety of commercial products that are available worldwide. Using marine fish as a sustainable and safe resource of omega-3 is in question because of global climate change, overfishing issues, and the increasing levels of environmental pollutants, such as heavy metals and radioactive materials found in the ocean [
52,
53]. Some marine protists have high contents of EPA and/or DHA [
6,
54,
55,
56,
57]. Therefore, this marine microalga may have the potential to be a clean and sustainable omega-3 source alternative to fish-based oil. On the other hand, an excessive intake of omega-6 causes negative health outcomes [
58,
59], even though both omega-3 and omega-6 PUFAs are essential, and they are required for many biological processes. Due to the ever-growing demands for food, the fish farming industry has rapidly expanded over the last few decades and aquaculture now accounts for over 50% of the fish consumed worldwide [
60]. This has caused further shortage of wild fish as feed at fish farms, and thus, fish oil in the aquaculture feed has been increasingly replaced by terrestrial vegetable oils that normally lack EPA and DHA, but often contain high levels of omega-6 PUFAs [
61]. The imbalance between omega-3 and omega-6 levels has deteriorated the nutritional quality of farmed fish such as Atlantic salmon that contains less EPA and DHA and more omega-6 PUFAs than before [
62,
63]. Thus, strain MABIKLP88 could be used for the production of designed aquafeeds for balancing the dietary omega-6 and omega-3 ratios. Furthermore, several authors reported that the amount of omega-3 PUFAs in the phytoplankton community is reflected in the nutritional quality of predatory fish [
7,
10,
11]. It should also be noted that the availability of omega-3 PUFAs in aquatic ecosystems is closely related to egg production and hatching success of marine copepods [
64,
65] and the fish larvae survival [
66]. Hence, the presence and abundance of this Korean dinoflagellate may have a potentially positive impact on the sustainability of both capture fisheries and aquaculture in Korea.
In this study, we report the first record of E. voratum from the Dokdo Islands, in the East Sea of Korea. This marine dinoflagellate would serve as potential biological resources to produce aquaculture feeds and biochemicals of commercial interests. Importantly, a clonal culture was established and deposited in a national culture collection to allow for further research.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}