
Fragmented Forest Patches in the Indian Himalayas Preserve Unique Components of Biodiversity: Investigation of the Floristic Composition and Phytoclimate of the Unexplored Bani Valley

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
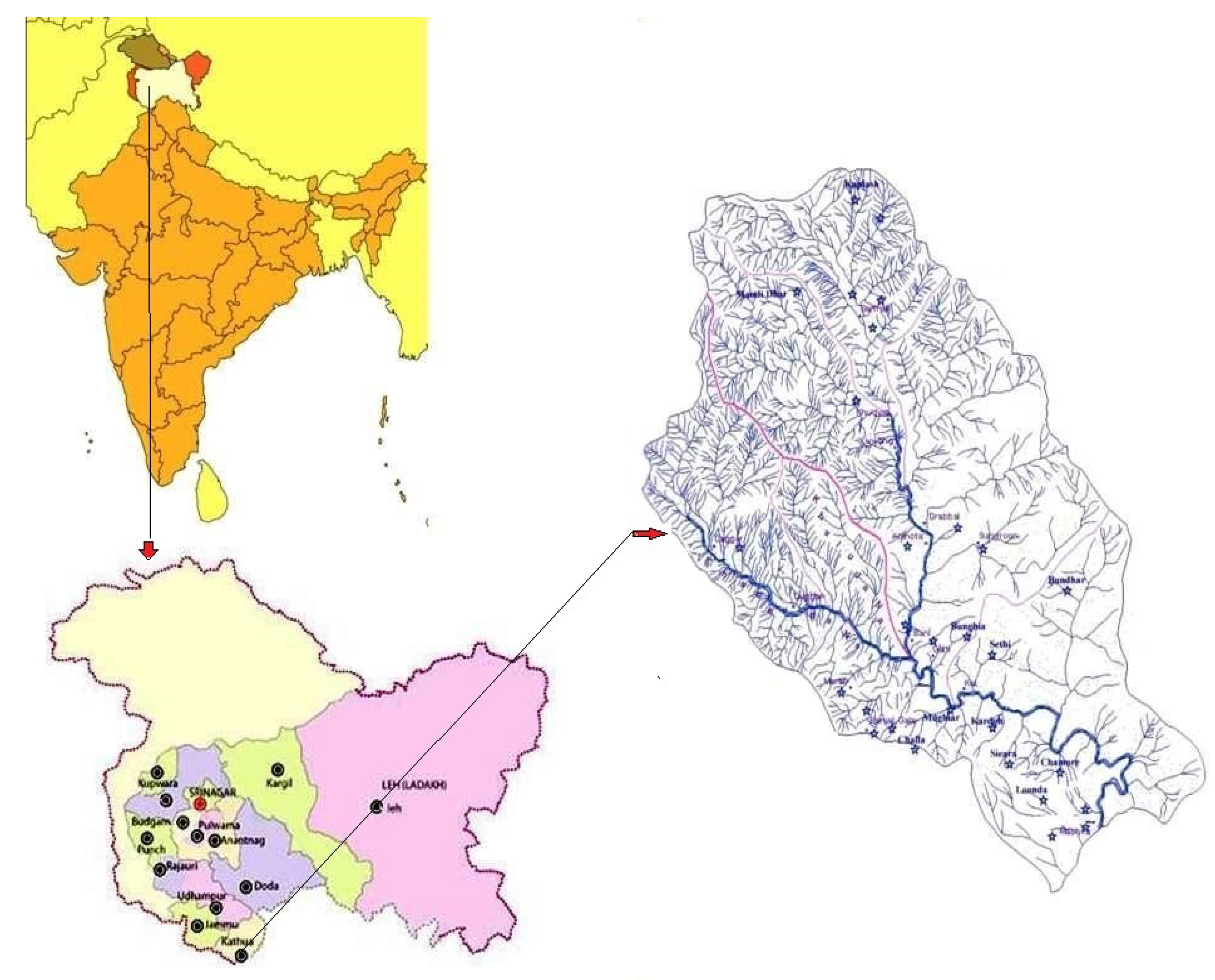
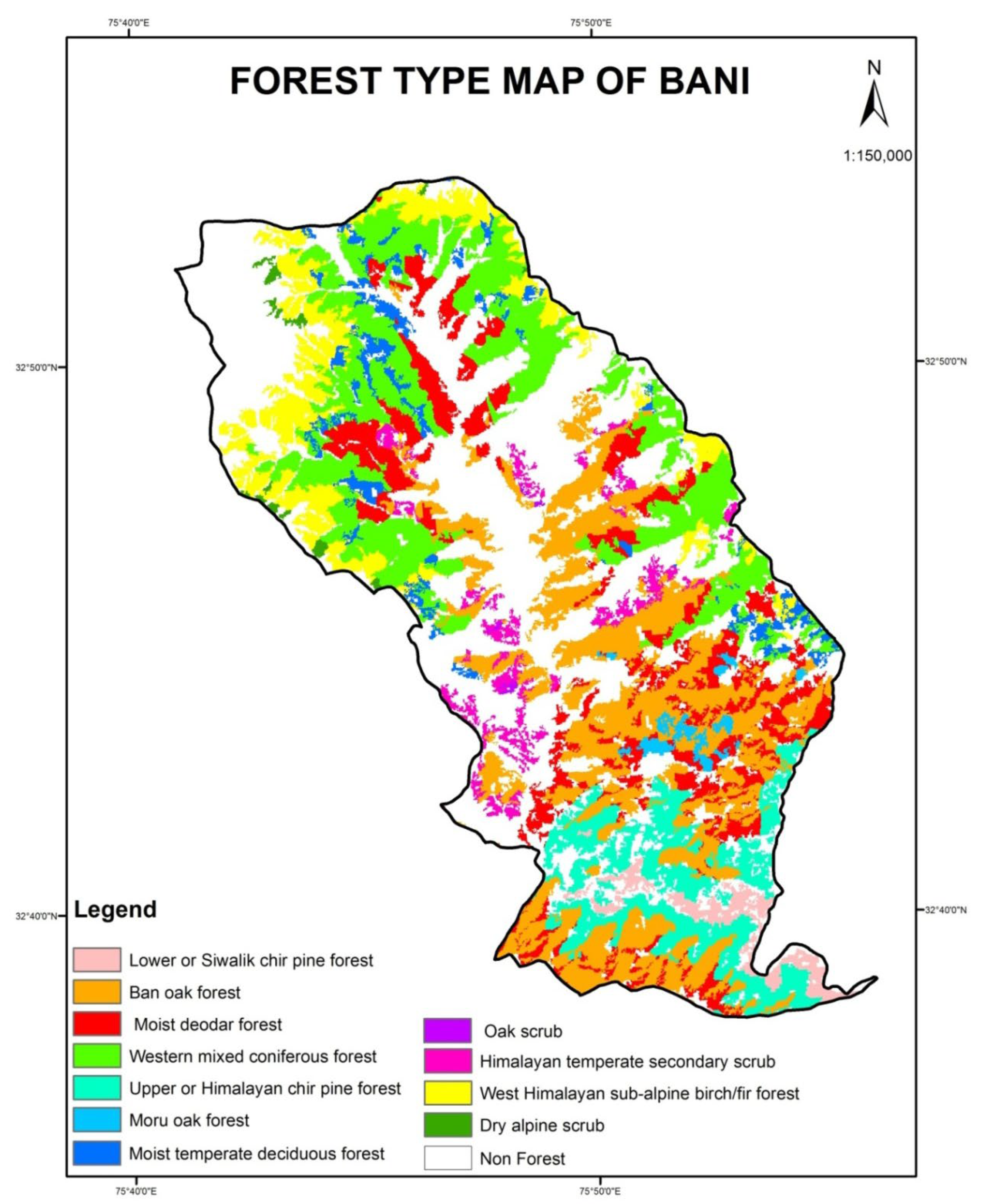
2.1. Study Area
2.2. Data Collection
2.2.1. Field Surveys
2.2.2. Presentation of Data
2.2.3. Literature Sources
3. Results and Discussion
3.1. Diversity of Taxa and Families
3.2. Species Diversity in Different Growth Form
3.3. Life Span
3.4. Life Form and Biological Spectrum
3.5. Leaf Size Spectrum
3.6. Phenological Periods
3.7. Invasive Species
3.8. Conservation Status and IUCN Categories
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Odum, E.P. Ecology and Our Endangered Life-Support Systems; Sinaur Associates, Inc.: Sunderland, MA, USA, 1993. [Google Scholar]
- Tang, Z.Y.; Fang, J.Y. A Review on the Elevational Patterns of Plant Species Diversity. Biodivers. J. 2004, 12, 20–28. [Google Scholar]
- Rahbek, C. The Role of Spatial Scale and the Perception of Large-scale Species-richness Patterns. Ecol. Lett. 2004, 8, 224–239. [Google Scholar] [CrossRef]
- Xu, W.; Xiao, Y.; Zhang, J.; Yang, W.; Zhang, L.; Hull, V.; Wang, Z.; Zheng, H.; Liu, J.; Polasky, S.; et al. Strengthening Protected Areas for Biodiversity and Ecosystem Services in China. Proc. Natl. Acad. Sci. USA 2017, 114, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, J.C.P.; Cano-Ortiz, A.; Musarella, C.M.; Canas, R.Q.; Gomes, C.J.P.; Spampinato, G.; Del Río, S.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus pyrenaica, Acer opalus subsp. granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y. Exploring Altitudinal Patterns of Plant Diversity of China’s Mountains. Biodivers. J. 2004, 12, 1–4. [Google Scholar]
- Sen, U.K.; Bhakat, R.K. Floristic and Phytoclimatic Study of a Sacred Grove Vegetation of West Midnapore District, West Bengal, India. J. Trop. Life Sci. 2019, 9, 119–138. [Google Scholar] [CrossRef]
- Adhikari, D.; Tiwari, R.; Singh, P.P.; Upadhay, K.; Singh, B.; Haridasan, K.E.; Bhatt, B.B.; Chetri, A.; Barik, S.K. Ecological Niche Modeling as a Cumulative Environmental Impact Assessment Tool for Biodiversity Assessment and Conservation Planning a Case Study of Critically Endangered Plant Lagerstroemia minuticarpa in Indian Eastern Himalaya. J. Environ. Manag. 2019, 243, 299–307. [Google Scholar]
- Muller, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; The Blackburn Press: Caldwell, NJ, USA, 1974. [Google Scholar]
- Singh, B.; Singh, B.; Borthakur, S.K.; Phukan, S.J. Contribution to Himalayan Hotspot: Assessment of Forest Types, Floristic Composition and Economic Wealth of Nokrek Biosphere Reserve in Northeast India. Indian Forest. 2018, 144, 734–741. [Google Scholar]
- Warming, E.; Balfour, I.B.; Groom, P.; Vahl, M. Oecology of Plants: An Introduction to the Study of Plant-Communities; Smithsonian Institution: Washington, DC, USA, 1909. [Google Scholar]
- Dansereau, P.; Guinochet, M.; Tomaselli, R. Two New European Commentaries on Phytosociology. Ecology 1957, 38, 364. [Google Scholar] [CrossRef]
- Singh, B.; Singh, B.; Singh, S.; Bhanwaria, R.; Chandra, S. Biological Spectrum and Floral Diversity of Western Himalaya—A Case Study of Nandini Wildlife Sanctuary in J&K State. In Angiopserm Systematics: Recent Trends and Emerging Issues; Agnihotri, P., Khuraijam, J.S., Eds.; Bishen Singh Mahendra Pal Singh Press: Dehradun, India, 2018; pp. 589–605. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Da Costa, R.C.; de Araujo, F.S.; Lima-Verde, L.W. Flora and Life-Form Spectrum in an Area of Deciduous Thorn Woodland (Caatinga) in Northeastern, Brazil. J. Arid. Environ. 2007, 68, 237–247. [Google Scholar] [CrossRef]
- Al-Yemeni, M.; Sher, S. Biological Spectrum with More Other Ecological Attributes of the Flora and Vegetation of the Asir Mountain of South West, Saudi Arabia. Afr. J. Biotech. 2010, 9, 5550–5559. [Google Scholar]
- Batalha, M.A.; Martins, F.R. Floristic, Frequency, and Vegetation Life-Form Spectra of a Cerrado Site. Braz. J. Biol. 2004, 64, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Badshah, L.; Hussain, F.; Sher, Z. Floristic Inventory, Ecological Characteristics and Biological Spectrum of Plants of Parachinar, Kurram Agency, Pakistan. Pak. J. Bot. 2016, 48, 1547–1558. [Google Scholar]
- Box, E.O. Macroclimate and Plant Forms: An Introduction to Predictive Modeling in Phytogeography; Dr. W. Junk: The Hague, The Netherlands, 1981. [Google Scholar]
- Gillison, A.N. Latitudinal Variation in Plant Functional Types. In Geographical Changes in Vegetation and Functional Types; Greller, A.M., Fujiwara, K., Pedrotti, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 21–57. [Google Scholar]
- Cain, S.A.; Castro, G.D.O. Manual of Vegetation Analysis; Harper and Brothers: New York, NY, USA, 1959. [Google Scholar]
- Meher-Homji, V.M. Life Forms and Biological Spectra as Epharmonic Criteria of Aridity and Humidity in Tropics. J. Indian Bot. Soc. 1964, 43, 424–430. [Google Scholar]
- Rana, T.S.; Datt, B.; Rao, R.R. Life Forms and Biological Spectrum of the Flora of Tons Valley, Garhwal Himalaya (Uttaranchal), India. Taiwania 2002, 47, 164–169. [Google Scholar]
- El-Ghani, M.M.; Amer, W.M. Soil–Vegetation Relationships in a Coastal Desert Plain of Southern Sinai, Egypt. J. Arid. Environ. 2003, 55, 607–628. [Google Scholar] [CrossRef]
- Austin, M.P. The Potential Contribution of Vegetation Ecology to Biodiversity Research. Ecography 1999, 22, 465–484. [Google Scholar] [CrossRef]
- Körner, C. Mountain Biodiversity, Its Causes and Function. Ambio 2004, 13, 11–17. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Lovette, I.J. The Roles of Island Area per se and Habitat Diversity in the Species-Area Relationships of Four Lesser Antillean Faunal Groups. J. Anim. Ecol. 1999, 68, 1142–1160. [Google Scholar] [CrossRef] [Green Version]
- Kalmar, A.; Currie, D.J. A Global Model of Island Biogeography. Glob. Ecol. Biogeogr. 2006, 15, 72–81. [Google Scholar] [CrossRef]
- Musarella, C.M.; Brullo, S.; Del Galdo, G.G. Contribution to the Orophilous Cushion-Like Vegetation of Central-Southern and Insular Greece. Plants 2020, 9, 1678. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate Effects on Mountain Plants. Nat. Cell Biol. 1994, 369, 448. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, G.; Gottfried, M.; Pauli, H. Long-term Monitoring of Mountain Peaks in the Alps. In Biomonitoring: General and Applied Aspects on Regional and Global Scales; Burga, C.A., Kratochwil, A., Eds.; Kluwer: Amsterdam, The Netherlands, 2001; pp. 153–177. [Google Scholar]
- Beniston, M. Climatic Change in Mountain Regions: A Review of Possible Impacts. Clim. Chang. 2003, 59, 5–31. [Google Scholar] [CrossRef]
- Boom, B.M. A Forest Inventory in Amazonian Bolivia. Biotropica 1986, 18, 287. [Google Scholar] [CrossRef]
- Rosati, L.; Romano, V.A.; Bartolucci, F.; Bernardo, L.; Bouvet, D.; Cancellieri, L.; Caruso, G.; Conti, F.; Faraoni, F.; Banfi, E.; et al. Contribution to the Floristic Knowledge of the Maddalena Mountains (Basilicata and Campania, Southern Italy). Ital. Bot. 2017, 3, 73–82. [Google Scholar] [CrossRef]
- Pérez-García, F.J.; Medina-Cazorla, J.M.; Martínez-Hernández, F.; Garrido-Becerra, J.A.; Mendoza-Fernández, A.J.; Salmerón Sánchez, E.; Mota, J.F. The Baetic Endemic Flora: Taxonomic Structure, Ecological and Rarity Patterns. Implications for a Conservation Policy. Ann. Bot. Fenn. 2012, 49, 43–54. [Google Scholar] [CrossRef]
- Resource Partner on Biodiversity. ENVIS Center on Biological Diversity. Available online: http://www.bsienvis.nic.in (accessed on 20 August 2020).
- Ashutosh, S.; Pandey, S.; Kaur, T.; Bajpai, R.K. Knowledge Based Remote Sensing and GIS Approach for Forest Type Mapping in Kathua District, Jammu and Kashmir. Trop. Ecol. 2010, 51, 21–29. [Google Scholar]
- Kumari, S.; Batish, D.R.; Singh, H.P.; Negi, K.; Kohli, R.K. An Ethnobotanical Survey of Medicinal Plants Used by Gujjar Community of Trikutta Hills in Jammu and Kashmir, India. J. Med. Plant Res. 2013, 7, 2111–2121. [Google Scholar]
- Thakur, S.; Dutt, H.C.; Singh, B.; Sharma, Y.P.; Tashi, N.; Charak, R.S.; Sharma, G.; Vidyarathi, O.P.; Iqbal, T.; Singh, B.; et al. Plant and Fungi Diversity of Devi Pindiyan Valley in Trikuta Hills of Northwestern Himalaya, India. J. Threat. Taxa 2019, 11, 14827–14844. [Google Scholar] [CrossRef]
- Singh, B. Swertia patnitopiansis, a New Species of Gentianaceae from North-Western Himalaya, India. Phytotaxa 2021, 498, 273–280. [Google Scholar] [CrossRef]
- Singh, B.; Sultan, P.; Bedi, Y.S. Juniperus chinensis L. (Cupressaceae): A New Taxa Record for Himalaya and Extension of Geographic Distribution in South Asia. Natl. Acad. Sci. Lett. 2018, 41, 69–73. [Google Scholar] [CrossRef]
- Singh, B.; Singh, S.; Singh, B. New Distribution Records of the Leopard Plants Ligularia amplexicaulis DC. and Ligularia sibirica (L.) Cass. (Asteraceae) in the Indian Himalaya. J. Threat. Taxa 2018, 10, 12854–12858. [Google Scholar] [CrossRef]
- Malik, A.H.; Khuroo, A.A.; Dar, G.H.; Khan, Z.S. An Annotated Inventory of Arboreal Flora in Jammu and Kashmir State. Bioprospecting 2020, 18, 565–606. [Google Scholar] [CrossRef]
- Singh, B.; Singh, B.; Kishor, A.; Singh, S.; Bhat, M.N.; Surmal, O.; Musarella, C.M. Exploring Plant-Based Ethnomedicine and Quantitative Ethnopharmacology: Medicinal Plants Utilized by the Population of Jasrota Hill in Western Himalaya. Sustainability 2020, 12, 7526. [Google Scholar] [CrossRef]
- Champion, H.G.; Seth, S.K. A Revised Survey of Forest Types of India; The Manager Publication of India: New Delhi, India, 1968. [Google Scholar]
- Whittaker, R.H. Dominance and Diversity in Land Plant Communities: Numerical Relations of Species Express the Importance of Competition in Community Function and Evolution. Science 1965, 147, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.A.; Boom, B.M.; De Carvalho, A.M.; Dos Santos, T.S. Southern Bahian Moist Forests. Bot. Rev. 1983, 49, 155–232. [Google Scholar] [CrossRef]
- Greig-Smith, P. Quantitative Plant Ecology, 3rd ed.; University of California Press: Berkeley, CA, USA, 1983. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Gentry, A.H. Tree Species Richness of Upper Amazonian Forests. Proc. Natl. Acad. Sci. USA 1988, 85, 156–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milliken, W. Structure and Composition of One Hectare of Central Amazonian Terra Firme Forest. Biotropica 1998, 30, 530–537. [Google Scholar] [CrossRef]
- Jain, S.K.; Rao, R.R. A Handbook of Field and Herbarium Methods; Today and Tomorrow’s Printer Publisher: New Delhi, India, 1977. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium 2020. Available online: http://sweetgum.nybg.org/science/ih (accessed on 12 August 2020).
- The Angiosperm Phylogeny Group. An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Swami, A.; Gupta, R.K. Flora of Udhampur; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1998; pp. 306–312. [Google Scholar]
- Sharma, B.M.; Kachroo, P. Flora of Jammu and Plants of Neighbourhood; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1981. [Google Scholar]
- Kapur, S.K.; Sarin, Y.K. Flora of Trikuta Hills; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1990. [Google Scholar]
- Duke, J.A. Handbook of Medicinal Herbs; CRC Press: London, UK, 2002. [Google Scholar]
- Sharma, B.M. Illustration of Jammu Plants; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2010. [Google Scholar]
- Dar, G.H.; Khuroo, A.A. Floristic Diversity in the Kashmir Himalaya: Progress, Problem and Prospects. Sains Malaysiana 2013, 42, 1377–1386. [Google Scholar]
- Sharma, P.; Rana, J.C.; Devi, U.; Randhawa, S.S.; Kumar, R. Floristic Diversity and Distribution Pattern of Plant Communities along Altitudinal Gradient in Sangla Valley, Northwest Himalaya. Sci. World J. 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chawla, A.; Rajkumar, S.; Singh, K.N.; Lal, B.; Singh, R.D.; Thukral, A.K. Plant Species Diversity along an Altitudinal Gradient of Bhabha Valley in Western Himalaya. J. Mt. Sci. 2008, 5, 157–177. [Google Scholar] [CrossRef]
- Chowdhery, H.J.; Wadhwa, B.M. Flora of Himachal Pradesh; Botanical Survey of India: Calcutta, India, 1984; pp. 1–3.
- Zent, E.L.; Zent, S. Floristic Composition, Structure, and Diversity of Four Forest Plots in the Sierra Maigualida, Venezuelan Guayana. Biodivers. Conserv. 2004, 13, 2453–2483. [Google Scholar] [CrossRef]
- Agrawal, A.K. Floristic Composition and Phenology of Temperate Grasslands of Western Himalaya as Affected by Scraping, Fire and Heavy Grazing. Vegetatio 1990, 88, 177–187. [Google Scholar] [CrossRef]
- Shaheen, H.; Ullah, Z.; Khan, S.M.; Harper, D.M. Species Composition and Community Structure of Western Himalayan Moist Temperate Forests in Kashmir. Forest. Ecol. Manag. 2012, 278, 138–145. [Google Scholar] [CrossRef]
- Haq, S.M.; Malik, A.H.; Khuroo, A.A.; Rashid, I. Floristic Composition and Biological Spectrum of Keran—A Remote Valley of Northwestern Himalaya. Acta Ecol. Sin. 2019, 39, 372–379. [Google Scholar] [CrossRef]
- Dhaliwal, D.S.; Sharma, M. Flora of Kullu District (Himachal Pradesh); Bishen Singh Mahendra Pal Singh Publisher: Dehradun, India, 1999. [Google Scholar]
- Haq, S.M.; Hamid, M.; Lone, F.A.; Singh, B. Himalayan Hotspot with Alien Weeds: A Case Study of Biological Spectrum, Phenology, and Diversity of Weedy Plants of High Altitude Mountains in District Kupwara of J&K Himalaya, India. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2021, 91, 139–152. [Google Scholar] [CrossRef]
- Mir, N.A.; Masood, T.H.; Sofi, P.A.; Husain, M.; Rather, T.A. Life form Spectrum of Vegetation in Betula Dominant Tree Stands along the Available Altitudinal Gradient in North-western Himalayas of Kashmir. Res. Rev. J. Pharmacogn. Phytochem. 2017, 6, 267–272. [Google Scholar]
- Dogra, K.S.; Sood, S.K.; Sharma, R. Distribution, Biology and Ecology of Parthenium hysterophorus L. (Congress Grass) an Invasive Species in the North-western Indian Himalaya (Himachal Pradesh). Acad. J. 2011, 5, 682–687. [Google Scholar]
- Gaston, A.; Garson, P.; Hunter, M. The Status and Conservation of Forest Wildlife in Himachal Pradesh, Western Himalayas. Biol. Conserv. 1983, 27, 291–314. [Google Scholar] [CrossRef]
- Gairola, S.; Sharma, C.M.; Suyal, S.; Ghildiyal, S.K. Composition and Diversity of Five Major Forest Types in Moist Temperate Climate of the Western Himalayas. For. Stud. China 2011, 13, 139–153. [Google Scholar] [CrossRef]
- Subramani, S.P.; Jishtu, V.; Verma, R.K.; Kapoor, K.S. Floristic Composition, Life-Forms and Biological Spectrum of Renuka Wildlife Sanctuary. Indian Forest. 2007, 133, 79–92. [Google Scholar]
- Saha, S. Determination of Life Form Patterns in the Singalila Range of Darjeeling. Environ. Ecol. 2008, 26, 588–590. [Google Scholar]
- Rawat, G.S.; Adhikari, B.S. Floristics and Distribution of Plant Communities across Moisture and Topographic Gradients in Tso Kar Basin, Changthang Plateau, Eastern Ladakh. Arctic Antarct. Alp. Res. 2005, 37, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Namgail Gopal, T.; Rawat, G.S.; Mishra, C.; Van Wieren, S.E.; Prins, H.H.T. Biomass and Diversity of Dry Alpine Plant Communities Along Altitudinal Gradients in the Himalayas. J. Plant Res. 2011, 125, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Klimes, L. Life-Forms and Clonality of Vascular Plants Along an Altitudinal Gradient in Eastern Ladakh (NW Himalayas). Basic Appl. Ecol. 2013, 4, 317–318. [Google Scholar] [CrossRef] [Green Version]
- Pharswan, K.; Mehta, J.P.; Subodh. Floristic Composition and Biological Spectrum of Vegetation in Alpine Meadows of Kedarnath: Garhwal Himalaya. Nat. Sci. 2010, 7, 109–115. [Google Scholar]
- Nautiyal, M.C.; Nautiyal, B.P.; Prakash, V. Phenology and Growth Form Distribution in an Alpine Pasture at Tungnath, Garhwal, Himalaya. Mt. Res. Dev. 2001, 21, 168–174. [Google Scholar] [CrossRef]
- Johnston, M.; Gilman, M. Tree Population Studies in Low Diversity Forests, Guyana. I. Floristic Composition and Stand Structure. Biodivers. Conserv. 1995, 4, 339–362. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Abundance, Growth and Mortality of Very Large Trees in Neotropical Lowland Rain Forest. Forest Ecol. Manag. 1996, 80, 235–244. [Google Scholar] [CrossRef]
- Saxena, A.K.; Pandey, P.; Singh, J.S. Biological Spectrum and Other Structural Functional Attributes of the Vegetation of Kumaun Himalaya. Vegetatio 1982, 49, 111–119. [Google Scholar] [CrossRef]
- Singh, B.; Bedi, Y.S. Eating from Raw Wild Plants in Himalaya: Traditional Knowledge Documentary on Sheena Tribe along LOC Border in Kashmir. Indian J. Nat. Prod. Resour. 2017, 8, 269–275. [Google Scholar]
- Das, D.S.; Rawat, D.S.; Maity, D.; Dash, S.S.; Sinha, B.K. Species Richness Patterns of Different Life-Forms along Altitudinal Gradients in the Great Himalayan National Park, Western Himalaya, India. Taiwania 2020, 65, 154–162. [Google Scholar]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Diaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global Climatic Drivers of Leaf Size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, H.; Khan, S.M.; Harper, D.M.; Ullah, Z.; Qureshi, R.A. Species Diversity, Community Structure, and Distribution Patterns in Western Himalayan Alpine Pastures of Kashmir, Pakistan. Mt. Res. Dev. 2011, 31, 153. [Google Scholar] [CrossRef] [Green Version]
- Prance, G.T. Discovering the Plant World. Taxon 2001, 50, 345–359. [Google Scholar] [CrossRef]
- Kala, C.P. Phenology of Alpine Plants in the Valley of Flowers National Park and Hemkund, Western Himalaya. Indian Forest. 1999, 125, 581–590. [Google Scholar]
- Pangtey, Y.P.S.; Rawal, R.S.; Bankoti, N.S.; Samant, S.S. Phenology of High-altitude Plants of Kumaun in Central Himalaya, India. Int. J. Biometeorol. 1990, 34, 122–127. [Google Scholar] [CrossRef]
- Bisht, V.K.; Kuniyal, C.P.; Bhandari, A.K.; Nautiyal, B.P.; Prasad, P. Phenology of Plants in Relation to Ambient Environment in a Subalpine Forest of Uttarakhand, Western Himalaya. Physiol. Mol. Biol. Plants 2014, 20, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Borthakur, S.K. Phenology and Geographic Extension of Lycophyta and Fern Flora of Nokrek Biosphere Reserve of Eastern Himalaya. Proc. Natl. Acad. Sci. India Sect. B. Biol. Sci. 2015, 85, 291–301. [Google Scholar] [CrossRef]
- Vashistha, R.K.; Rawat, N.; Chaturvedi, A.K.; Nautiyal, B.P.; Prasad, P.; Nautiyal, M.C. An Exploration on the Phenology of Different Growth Forms of an Alpine Expanse of North-West Himalaya, India. N. Y. Sci. J. 2009, 2, 29–40. [Google Scholar]
- Liu, J.; Liang, S.C.; Liu, F.H.; Wang, R.Q.; Dong, M. Invasive Alien Plant Species in China: Regional Distribution Patterns. Divers. Distrib. 2005, 11, 341–347. [Google Scholar] [CrossRef]
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien Species in a Warmer World: Risks and Opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaFace, V.L.A.; Musarella, C.M.; Ortiz, A.C.; Canas, R.Q.; Cannavò, S.; Spampinato, G. Three New Alien Taxa for Europe and a Chorological Update on the Alien Vascular Flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef]
- Reddy, C.S. Catalogue of Invasive Alien Flora of India. Life Sci. J. 2008, 5, 84–89. [Google Scholar]
- Wu, S.H.; Sun, H.T.; Teng, Y.C.; Rejmánek, M.; Chaw, S.M.; Yang, T.Y.A.; Hsieh, C.F. Patterns of Plant Invasions in China: Taxonomic, Biogeographic, Climatic Approaches and Anthropogenic Effects. Biol. Invasions 2010, 12, 2179–2206. [Google Scholar] [CrossRef] [Green Version]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P.; et al. Alien Flora of Europe: Species Diversity, Temporal Trends, Geographical Patterns and Research Needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Khuroo, A.A.; Weber, E.; Malik, A.H.; Dar, G.H.; Reshi, Z.A. Taxonomic and Biogeographic Patterns in the Native and Alien Woody Flora of Kashmir Himalaya, India. Nord. J. Bot. 2010, 28, 685–696. [Google Scholar] [CrossRef]
- Randall, R.P. The Introduced Flora of Australia and Its Weed Status; CRC Press: Adelaide, Australia, 2007. [Google Scholar]
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibáñez, I.; et al. Will Extreme Climatic Events Facilitate Biological Invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Kohli, R.K.; Dogra, K.S.; Batish, D.R.; Singh, H.P. Impact of Invasive Plants on the Structure and Composition of Natural Vegetation of Northwestern Indian Himalayas1. Weed Technol. 2004, 18, 1296–1300. [Google Scholar] [CrossRef]
- Khuroo, A.A.; Reshi, Z.A.; Malik, A.H.; Weber, E.; Rashid, I.; Dar, G.H. Alien Flora of India: Taxonomic Composition, Invasion Status and Biogeographic Affiliations. Biol. Invasions 2011, 14, 99–113. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species, Version 2019-1. Available online: https://www.iucnredlist.org (accessed on 12 August 2020).
- Bijalwan, R.; Vats, M.; Joshi, S.P. Plant Phenological Response to Microclimatic Variations in an Alpine Zone of Garhwal Himalaya. J. Appl. Nat. Sci. 2013, 5, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Lanker, U.; Malik, A.R.; Gupta, N.K.; Butola, J.S. Natural Regeneration Status of the Endangered Medicinal Plant, Taxus baccata Hook.f. syn. T. wallichiana, in Northwest Himalaya. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2010, 6, 20–27. [Google Scholar] [CrossRef]
- Paul, A.; Bharali, S.; Khan, M.L.; Tripathi, O.P. Anthropogenic Disturbances Lead to Risk of Extinction of Taxus wallichiana Zuccarini, an Endangered Medicinal Tree in Arunachal Himalaya. Nat. Areas J. 2013, 33, 447–454. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Orders | Families | Genera | Taxa | |||
|---|---|---|---|---|---|---|---|
| Herbs | Shrubs | Trees | Total | ||||
| Dicots | 20 | 52 | 117 | 48 | 34 | 52 | 134 |
| Monocots | 5 | 10 | 37 | 46 | 1 | 0 | 47 |
| Lycophytes and ferns | 1 | 4 | 8 | 10 | 0 | 0 | 10 |
| Gymnosperms | 1 | 2 | 4 | 0 | 0 | 5 | 5 |
| Total | 27 | 68 | 166 | 104 | 35 | 57 | 196 |
| Habit | Number of Taxa | Number of Individuals | Dominance Index | Shannon Index | Evenness Index | |
|---|---|---|---|---|---|---|
| A. Subtropical forests (1201–1600 m) | Trees | 45 | 415 | 0.41 | 3.51 | 0.74 |
| Shrubs | 23 | 480 | 0.08 | 2.75 | 0.68 | |
| Herbs | 42 | 96,000 | 0.06 | 3.01 | 0.76 | |
| B. Temperate forests (1601–2000 m) | Trees | 19 | 400 | 0.11 | 2.53 | 0.66 |
| Shrubs | 20 | 355 | 0.12 | 2.31 | 0.76 | |
| Herbs | 59 | 162,800 | 0.03 | 3.77 | 0.77 |
| Raunkiaer ‘s Life Forms | Total No. of Species | Biological Spectra (%) of the Bani Valley | Raunkiaer’sNormal Spectra (%) | Deviation = (Raunkiaer’s Normal Spectra–Biological Spectra) |
|---|---|---|---|---|
| PH | 96 | 48.98 | 46.00 | 2.98 |
| CHA | 16 | 8.16 | 9.00 | −0.84 |
| GEO | 4 | 2.04 | 6.00 | −3.96 |
| HCP | 30 | 15.31 | 26.00 | −10.69 |
| THP | 50 | 25.51 | 13.00 | 12.51 |
| Total | 196 | 100.00 | 100.00 | 0.00 |
| Raunkiaer’s Life Forms | Leptophyllous | Megaphyllous | Mesophyllous | Microphyllous | Nanophyllous | Total |
|---|---|---|---|---|---|---|
| CHA | 0 | 0 | 6 | 3 | 7 | 16 |
| GEO | 0 | 1 | 2 | 0 | 1 | 4 |
| HCP | 1 | 0 | 10 | 7 | 12 | 30 |
| PH | 5 | 12 | 37 | 20 | 22 | 96 |
| THP | 4 | 1 | 13 | 11 | 21 | 50 |
| Total | 10 | 14 | 68 | 41 | 63 | 196 |
| Plant Groups (APG IV) | Voucher Number | Habit | Life Span | Phenology Period | Habitat | Raunkiaer’s Life Forms | Sub-type of Raunkiaer’s Life Forms | Leaf Spectrum | Leaf Shape | Conservation Status (IUCN) | Specific Distribution | Native (N)/Exotic (E) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I. MONOCOTS | ||||||||||||
| Acorales Martinov | ||||||||||||
| Acoraceae Martinov | ||||||||||||
| Acorus calamus L. | RRLH54665 | H | P | May–June | Streamside | GEO | MES | Ensiform | LC | Native to Asia | N | |
| Alismatales R.Br. ex Bercht. & J. Presl | ||||||||||||
| Potamogetonaceae Bercht. & J.Presl | ||||||||||||
| Potamogeton nodosus Poir. | RRLH55286 | H | P | Jul–Aug | Aquatic | CHA | NP | Lanceolate | LC | Native to North America | E | |
| Araceae Juss. | ||||||||||||
| Arisaema flavum (Forssk.) Schott | RRLH55268 | H | P | July–Aug | Forest slopes | GEO | MES | Oblong to lanceolate | NA | Native to Asia | N | |
| Juncaceae Juss. | ||||||||||||
| Juncus bufonius L. | RRLH55278 | H | A | May–Jun | Streamside | THP | MES | Elliptic | LC | Native to North America | E | |
| Smilacaceae Vent. | ||||||||||||
| Smilax vaginata Decne. | RRLH54673 | S | P | May–Aug | Forest thickets | PH | NPH | LEP | Ovate | NA | Native to Asia and Afghanistan | N |
| Asparagales | ||||||||||||
| Asparagaceae Juss. | ||||||||||||
| Asparagus adscendens Roxb. | RRLH55270 | H | P | Nov–Dec | Forest thickets | PH | NPH | MIC | Spiny | NA | Native to India | N |
| Orchidaceae Juss. | ||||||||||||
| Rhynchostylis retusa (L.) Blume | RRLH55205 | H | A | May–Jun | Epiphytic | THP | LEP | Lorate | NA | Native to Asia | N | |
| Commelinales Mirb. ex Bercht. & J. Presl | ||||||||||||
| Commelinaceae Mirb. | ||||||||||||
| Commelina benghalensis L. | RRLH54667 | H | P | Apr–May | Wet places | CHA | MES | Ovate | LC | Native to Asia | N | |
| Commelina communis L. | RRLH54941 | H | A | Apr–May | Moist places | THP | MES | Lanceolate | NA | Native to Asia | N | |
| Poales Small | ||||||||||||
| Cyperaceae Juss. | ||||||||||||
| Cyperus alulatus J. Kern | RRLH54630 | H | A | Jun–Jul | Forest slopes | THP | NP | Elliptic | LC | Native to the Indian subcontinent | N | |
| Carex brunnea Thunb. | RRLH55270 | H | P | Jun–Jul | Mountain slopes | HCP | NP | Lanceolate | NA | Native to Africa | E | |
| Carex muricata L. | RRLH55271 | H | P | May–Jun | Hill slopes | HCP | MIC | Lanceolate | NA | Native to North America | E | |
| Cyperus niveus Retz. | RRLH54667 | H | A | Sep–Oct | Stream margins | THP | MIC | Elliptic | NA | Native to Asia | N | |
| Cyperus paniceus (Rottb.) Boeckeler | RRLH55274 | H | P | Apr–May | Grasslands | HCP | NP | Oblong | LC | Native to Africa | E | |
| Cyperus rotundus L. | RRLH52669 | H | A | May–Jun | Grasslands | THP | MIC | Elliptic | LC | Native to Africa and Eurasia | E | |
| Eleocharis palustris (L.) Roem. & Schult. | RRLH55276 | H | P | Jun–Jul | Stream margins | HCP | MIC | Linear | LC | Native to North America | E | |
| Eriophorum comosum (Wall.) Nees | RRLH54904 | H | P | May–Jun | Rock crevices | THP | NP | Linear | LC | Native to Asia | N | |
| Schoenoplectus lacustris (L.) Roem. & Schult. | RRLH55288 | H | P | Jun–Jul | Swampy areas | HCP | NP | Linear | LC | Native to Europe | E | |
| Poaceae Barnhart | ||||||||||||
| Alopecurus arundinaceus Poir. | RRLH54668 | H | P | Jul–Aug | Grasslands | CHA | NP | Linear | LC | Native to Eurasia | E | |
| Arthraxon lancifolius (Trin.) Hochst. | RRLH52617 | H | A | Sep–Oct | Rocky surfaces | THP | NP | Elliptic | LC | Native to Eurasia | E | |
| Arundinella pumila (Hochst.) Steud. | RRLH55269 | H | P | Aug–Sep | Grasslands | HCP | MES | Linear to lanceolate | NA | Native to Asia | N | |
| Brachiaria ramosa (L.) Stapf | RRLH52615 | H | A | May–Jun | Grasslands | THP | MIC | Lanceolate | LC | Native to tropical Africa | E | |
| Cenchrus ciliaris L. | RRLH54626 | H | A | May–Jun | Wastelands | HCP | MES | Linear | LC | Native to tropical Africa | E | |
| Chrysopogon fulvus (Spreng.) Choiv. | RRLH54960 | H | A | Jun–Jul | Moist places | THP | MES | Linear | NA | Native to Asia | N | |
| Chrysopogon gryllus (L.) Trin. | RRLH55273 | H | P | Aug–Sep | Moist places | HCP | NP | Linear | NA | Native to Eurasia | E | |
| Cymbopogon distans (Nees ex Steud.) W.Watson | RRLH54669 | H | P | Jun–Jul | Open, grassy places | HCP | MES | Linear to filiform | NA | Native to India and China | N | |
| Cymbopogon jwarancusa (Jones) Schult. | RRLH54670 | H | P | Mar–May | Mountain slopes | HCP | MIC | Linear | NA | Native of Africa | E | |
| Cynodon dactylon (L.) Pers. | RRLH54671 | H | P | Jan–Dec | Roadsides | HCP | NP | Linear | NA | Native to Africa | E | |
| Echinochloa stagnina (Retz.) P.Beauv. | RRLH55275 | H | P | May–Jun | Moist places | HCP | NP | Linear | LC | Native to Africa | E | |
| Isachne himalaica Hook.f. | RRLH54971 | H | P | May–Jun | Swampy places | HCP | NP | Linear | NA | Native to the Himalayas | N | |
| Leersia hexandra Sw. | RRLH55279 | H | P | May–June | Streamside | HCP | NP | Linear | LC | Native to America | E | |
| Melinis minutiflora P.Beauv. | RRLH54627 | H | P | Jul–Aug | Field margins | HCP | MES | Linear | NA | Native to Africa | E | |
| Microstegium nudum (Trin.) A.Camus | RRLH55280 | H | A | Aug–Sep | Field margins | THP | NP | Linear | NA | Native to Africa | E | |
| Miscanthus nepalensis (Trin.) Hack. | RRLH55281 | H | P | Aug–Sep | Mountain slopes | HCP | MES | Linear | NA | Native to the Himalayas | N | |
| Oplismenus burmanii (Retz.) P.Beauv | RRLH55282 | H | P | Sep–Oct | Moist places | HCP | NP | Lanceolate | NA | Native to Africa | E | |
| Panicum virgatum L. | RRLH54628 | H | P | Jul–Aug | Cultivated | THP | NP | Linear | LC | Native to North America | E | |
| Paspalum vaginatum Sw. | RRLH54986 | H | P | Jun–Jul | Swampy areas | HCP | MIC | Linear | LC | Native to North America | E | |
| Pennisetum flaccidum Griseb. | RRLH55283 | H | P | Jul–Aug | Grasslands | HCP | MIC | Linear | LC | Native to the Himalayas | N | |
| Poa annua L. | RRLH55284 | H | A | Apr–May | Moist places | THP | MES | Linear | LC | Native to America | E | |
| Polypogon fugax Nees ex Steud. | RRLH55285 | H | A | Jun–Aug | Moist places | THP | NP | Linear | NA | Native to South America | E | |
| Saccharum filifolium Steud. | RRLH55287 | H | P | Apr–May | Grasslands | HCP | MIC | Linear | NA | Native to the Himalayas | N | |
| Saccharum spontaneum L. | RRLH54953 | H | P | Jul–Aug | Mountain slopes | HCP | NP | Linear | LC | Native to the Indian subcontinent | N | |
| Setaria flavida (Retz.) Veldkamp | RRLH55290 | H | P | Jul–Aug | Roadsides | HCP | MES | Linear to lanceolate | NA | Native to Africa | E | |
| Setaria italica (L.) P.Beauv | RRLH55291 | H | A | May–Jul | Cultivated | THP | NP | Linear | NA | Native to Eurasia | E | |
| Setaria viridis (L.) P.Beauv | RRLH55292 | H | A | May–Jun | Roadsides | THP | LEP | Linear | NA | Native to Asia | N | |
| Thysanolaena latifolia (Roxb. ex Hornem) Honda | RRLH54952 | H | P | May–Dec | Hillsides | HCP | NP | Lanceolate | NA | Native to Asia | N | |
| Tragus racemosus (L.) All. | RRLH55294 | H | P | May–Jun | Roadsides | THP | NP | Linear | NA | Native to Eurasia and Africa | E | |
| II. EUDICOTS | ||||||||||||
| Ranunculales Juss. ex Bercht. & J.Presl | ||||||||||||
| Ranunculaceae Juss. | ||||||||||||
| Clematis grata Wall. | RRLH55388 | S | P | Jul–Aug | Forest margins | PH | NPH | NP | Ovate | NA | Native to the Himalayas | N |
| Clematis graveolens Lindl. | RRLH55336 | S | P | Apr–May | Forest thickets | PH | NPH | NP | Ovate | NA | Native to Afghanistan and India | N |
| Thalictrum foliolosum DC. | RRLH55367 | S | P | May–Jun | Montane forests | PH | NPH | MIC | Elliptic to ovate | NA | Native to the Indian subcontinent | N |
| Thalictrum virgatum Hook.f. & Thomson | RRLH55245 | S | P | Jun–Jul | Forest margins | PH | NPH | MIC | Rhombic | NA | Native to the Himalayas | N |
| Berberidaceae Juss. | ||||||||||||
| Berberis lycium Royle | RRLH55359 | S | P | Apr–May | Forest margins | PH | NPH | NP | Oblanceolate | NA | Native to the Himalayas | N |
| Platanaceae T.Lestib. | ||||||||||||
| Platanus orientalis L. | RRLH55215 | T | P | Mar–May | Cultivated | PH | MPH | NP | Ovate | DD | Native to Europe | E |
| Buxales Takht. ex Reveal | ||||||||||||
| Buxaceae Dumort. | ||||||||||||
| Sarcococca saligna (D.Don) Müll.Arg. | RRLH55268 | S | P | May–Jun | Evergreen forests | PH | NPH | MES | Lanceolate | NA | Native to India and Pakistan | N |
| Papaveraceae Juss. | ||||||||||||
| Argemone mexicana L. | RRLH54671 | H | A | Mar–Apr | Streamside | THP | MIC | Oblanceolate | NA | Native to North America | E | |
| Menispermaceae Juss. | ||||||||||||
| Cissampelos pareira L. | RRLH54662 | S | P | Mar–Apr | Forest margins | PH | NPH | NP | Ovate | NA | Native to Asia | N |
| Tinospora cordifolia (Willd.) Hook.f. & Thomson | RRLH54670 | S | P | May–Jun | Forest thickets | PH | NPH | MES | Cordate | NA | Native to Asia | N |
| Piperales Bercht. & J.Presl | ||||||||||||
| Piperaceae Giseke | ||||||||||||
| Peperomia tetraphylla (G.Forst.) Hook. & Arn. | RRLH55254 | H | P | Feb–Mar | Epiphytes | GEO | NP | Elliptic | NA | Native to Asia | N | |
| Laurales Juss. | ||||||||||||
| Lauraceae Juss. | ||||||||||||
| Neolitsea umbrosa (Nees) Gamble | RRLH54992 | T | P | Mar–May | Forest margins | PH | MPH | MES | Oblong | NA | Native to the Himalayas | N |
| III. CORE EUDICOTS | ||||||||||||
| SUPERROSIDS | ||||||||||||
| Saxifragales Bercht. & J. Presl | ||||||||||||
| Crassulaceae J.St.–Hil. | ||||||||||||
| Rosularia adenotricha (Wall. ex Edgew.) C.-A.Jansson | RRLH54904 | H | A | May–Jun | Rock crevices | THP | MIC | Obovate | NA | Native to the Himalayas | N | |
| Fabales Bromhead | ||||||||||||
| Fabaceae Lindl. | ||||||||||||
| Acacia concinna (Willd.) DC. | RRLH54925 | S | P | April–Jun | Forest thickets | PH | NPH | MIC | Ovate | NA | Native to Asia | N |
| Albizia chinensis (Osbeck) Merr. | RRLH54675 | T | P | Mar–May | Open areas | PH | MPH | MIC | Oblong | NA | Native to Southeast Asia | N |
| Argyrolobium roseum (Cambess.) Jaub. & Spach | RRLH55372 | H | A | May–Jun | Forest margins | THP | MIC | Obovate | NA | Native to India and Pakistan | N | |
| Bauhinia purpurea L. | RRLH55373 | T | P | May–Jun | Cultivated | PH | MPH | MIC | Suborbicular | LC | Native to the Indian subcontinent | N |
| Biancaea decapetela (Roth.) O.Deg. | RRLH55374 | S | P | Apr–May | Roadsides | PH | NPH | MIC | Ovate | NA | Native to Asia | N |
| Butea monosperma (Lam.) Kuntze | RRLH54676 | T | P | Mar–Apr | Forest margins | PH | MPH | NP | Obovate | LC | Native to Asia | N |
| Cassia fistula L. | RRLH55375 | T | P | Aug–Sep | Roadsides | PH | MGP | MES | Ovate | LC | Native to the Indian subcontinent | N |
| Chamaecrista mimosoides (L.) Greene | RRLH55611 | S | P | Apr–May | Wastelands | PH | NPH | MEG | Linear | LC | Native to Africa | E |
| Dalbergia sissoo Roxb. ex DC. | RRLH54931 | T | P | Mar–Apr | Cultivated | PH | MPH | MEG | Ovate | LC | Native to India | N |
| Delonix regia (Bojer ex Hook.) Raf. | RRLH55375 | T | P | Jun–Jul | Cultivated | PH | MPH | NP | Oblong | LC | Native to Madagascar | E |
| Erythrina indica Lam. | RRLH55376 | T | P | Apr–May | Field margins | PH | MPH | MES | Lanceolate | LC | Native to Africa and Asia | N |
| Grona triflora (L.) H.Ohashi & K.Ohashi | RRLH54905 | S | P | May–Jun | Forest thickets | PH | NPH | MIC | Obovate | NA | Native to America | E |
| Lespedeza juncea (L.f.) Pers. | RRLH54636 | H | P | Jul–Aug | Forest thickets | HCP | NP | Oblanceolate | LC | Native to Asia | N | |
| Ototropis multiflora (DC.) H. Ohashi & K. Ohashi | RRLH55363 | S | P | Jul–Aug | Mountain slopes | PH | NPH | MIC | Elliptic | NA | Native to the Indian subcontinent | N |
| Phyllodium elegans (Lour.) Desv. | RRLH54987 | S | P | May–Jun | Forest thickets | PH | NPH | NP | Ovate | LC | Native to the Himalayas | N |
| Robinia pseudoacacia L. | RRLH54602 | T | P | May–Jun | Cultivated | PH | MPH | MES | Oblong | LC | Native to North America | E |
| Senegalia modesta (Wall.) P.J.H.Hurter | RRLH54674 | T | P | Mar–May | Cultivated | PH | MPH | MES | Ovate | NA | Native to the Indian subcontinent | N |
| Rosales Bercht. & J.Presl | ||||||||||||
| Cannabaceae Martinov | ||||||||||||
| Cannabis sativa L. | RRLH54915 | H | A | May–Jun | Wastelands | THP | NP | Lanceolate | NA | Native to Asia | N | |
| Celtis australis L. | RRLH55360 | T | P | Mar–May | Field margins | PH | MPH | NP | Ovate | LC | Native to the Mediterranean region and West Asia | E |
| Elaeagnaceae Juss. | ||||||||||||
| Elaeagnus umbellata Thunb. | RRLH55354 | T | P | Apr–May | Forest thickets | PH | MPH | MIC | Obovate | LC | Native to Asia | N |
| Rosaceae Juss. | ||||||||||||
| Prinsepia utilis Royle | RRLH55224 | S | P | Apr–May | Wastelands | PH | NPH | MIC | Ovate to lanceolate | NA | Native to the Western Himalayas | N |
| Prunus domestica L. | RRLH55390 | T | P | May–Jun | Cultivated | PH | MPH | NP | Elliptic | DD | Native to North America | E |
| Prunus persica (L.) Batsch. | RRLH55391 | T | P | Apr–May | Cultivated | PH | MPH | MIC | Elliptic | LC | Native to Asia | N |
| Pyrus pashia Buch.-Ham. ex D.Don | RRLH54901 | T | P | Mar–May | Forest thickets | PH | MPH | MES | Ovate | LC | Native to the Himalayas | N |
| Pyrus pyrifolia (Burm.f.) Nakai | RRLH55392 | T | P | May–Jun | Field margins | PH | MPH | MES | Ovate toelliptic | NA | Native to Southeast Asia | N |
| Rosa moschata Herrm. | RRLH55393 | S | P | Jun–Jul | Forest thickets | PH | NPH | MES | Oblong to lanceolate | NA | Native to Iran and Afghanistan | E |
| Rubus ellipticus Sm. | RRLH54951 | S | P | Mar–Apr | Mountain slopes | PH | NPH | MES | Ovate | LC | Native to Asia | N |
| Rubus idaeus L. | RRLH55210 | S | P | May–Jun | Forest thickets | PH | NPH | NP | Ovate to lanceolate | NA | Native to Eurasia | E |
| Rubus paniculatus Sm. | RRLH55292 | S | P | Jun–Jul | Forest thickets | PH | NPH | NP | Ovate to lanceolate | NA | Native to the Himalayas | N |
| Rhamnaceae Juss. | ||||||||||||
| Ziziphus mauritiana Lam. | RRLH55389 | T | P | Aug–Sep | Forest thickets | PH | MPH | MES | Ovate | LC | Native to India | N |
| Ulmaceae Mirb. | ||||||||||||
| Trema orientalis (L.) Blume | RRLH55370 | T | P | Mar–Apr | Mountain slopes | PH | MPH | NP | Lanceolate to ovate | LC | Native to Asia and Africa | N |
| Ulmus wallichiana Planch. | RRLH55400 | T | P | Apr–May | Field margins | PH | MPH | MES | Ovate | VU | Native to the Himalayas | N |
| Moraceae Gaudich. | ||||||||||||
| Ficus auriculata Lour. | RRLH55383 | T | P | Aug–Sep | Forest margins | PH | MPH | MES | Ovate to cordate | LC | Native to Asia | N |
| Ficus hispida L.f. | RRLH55383 | T | P | Jun–Jul | Forest margins | PH | MPH | MES | Ovate to oblong | LC | Native to Asia | N |
| Ficus palmata Forssk. | RRLH55224 | T | P | May–Jun | Roadsides | PH | MPH | MES | Ovate | NA | Native to the Indian subcontinent | N |
| Morus alba L. | RRLH55385 | T | P | Apr–May | Cultivated | PH | MPH | MES | Ovate | LC | Native to North America | E |
| Urticaceae Juss. | ||||||||||||
| Debregeasia saeneb (Forssk.) Hepper & J.R.I.Wood | RRLH54948 | S | P | Mar–Apr | Shady places | PH | MES | Lanceolate | NA | Native to the Himalayas | N | |
| Elatostema sessile J.R.Forst. & G.Forst. | RRLH54662 | H | P | May–Jun | Forest margins | CHA | NP | Linear | NA | Native to the Indian subcontinent | N | |
| Fleurya interrupta (L.) Gaudich | RRLH54663 | H | A | Jul–Aug | Moist places | THP | MES | Ovate | NA | Native to Asia and Australia | N | |
| Pilea scripta (Buch.- Ham. ex D.Don) Wedd. | RRLH54664 | H | P | Jun–Jul | Shady places | PH | MIC | Elliptic | NA | Native to the Himalayas | N | |
| Pilea umbrosa Wedd. ex Blume | RRLH55664 | H | P | Jul–Aug | Shady places | CHA | MES | Ovate | NA | Native to the Himalayas | N | |
| Urtica dioica L. | RRLH54991 | H | P | Jun–Jul | Forest thickets | HCP | MES | Ovate | LC | Native to Eurasia | E | |
| Fagales Engl. | ||||||||||||
| Fagaceae Dumort. | ||||||||||||
| Castanea sativa Mill. | RRLH55272 | T | P | Apr–May | Forest margins | PH | MPH | MES | Elliptic | LC | Native to Europe | E |
| Lithocarpus henryi (Seemen) Rehder & E.H.Wilson | RRLH55377 | T | P | Aug–Sep | Mixed forests | PH | MPH | MES | Oblong | LC | Native to the Himalayas | N |
| Quercus oblongata D.Don | RRLH54684 | T | P | May–Jun | Mixed forests | PH | MPH | MES | Oblong | NT | Native to Asia | N |
| Betulaceae Gray | ||||||||||||
| Alnus nitida (Spach) Endl. | RRLH55302 | T | P | Mar–Apr | Forest margins | PH | MPH | MIC | Elliptic | LC | Native to the Himalayas | N |
| Juglandaceae DC. ex Perleb | ||||||||||||
| Juglans regia L. | RRLH54857 | T | P | Apr–May | Cultivated | PH | MPH | MES | Elliptic | LC | Native to Eurasia | E |
| Cucurbitales Juss. ex Bercht. & J.Presl | ||||||||||||
| Cucurbitaceae Juss. | ||||||||||||
| Solena amplexicaulis (Lam.) Gandhi | RRLH54604 | S | P | Apr–May | Forest thickets | PH | NPH | MIC | Ovate | NA | Native to Asia | N |
| Combretaceae R.Br. | ||||||||||||
| Terminalia bellirica (Gaertn.) Roxb. | RRLH55361 | T | P | Mar–Apr | Field margins | PH | MPH | MEG | Obovate | NA | Native to Asia | N |
| Terminalia chebula Retz. | RRLH54361 | T | P | May–Jun | Forest margins | PH | MPH | MEG | Elliptic | LC | Native to Asia | N |
| Lythraceae J.St.-Hil. | ||||||||||||
| Punica granatum L. | RRLH55379 | S | P | May–Jun | Cultivated | PH | NPH | MEG | Elliptic to oblanceolate | LC | Native to Iran and India | N |
| Woodfordia fruticosa (L.) Kurz | RRLH54914 | T | P | Jan–Feb | Forest slopes | PH | MPH | NP | Lanceolate | LC | Native to Asia | N |
| Malpighales Juss. ex Bercht. & J.Presl | ||||||||||||
| Hypericaceae Juss. | ||||||||||||
| Hypericum perforatum L. | RRLH54665 | H | P | Jun–Jul | Grassland slopes | PH | NP | Oblong | NA | Native to Eurasia | E | |
| Violaceae Batsch. | ||||||||||||
| Viola odorata L. | RRLH55218 | H | P | Apr–May | Forest Slopes | THP | NP | Ovate | NA | Native to Asia | N | |
| Euphorbiaceae Juss. | ||||||||||||
| Euphorbia hirsuta L. | RRLH54944 | H | A | Jun–Jul | Roadsides | HCP | MIC | Lanceolate to ovate | NA | Native to India | N | |
| Euphorbia thymifolia L. | RRLH54920 | H | A | Jun–Jul | Roadsides | THP | NP | Lanceolate | NA | Native to America | E | |
| Phyllanthus emblica L. | RRLH52672 | T | P | Apr–May | Open areas | PH | MPH | NP | Oblong | LC | Native to India | N |
| Triadica sebifera (L.) Small | RRLH55371 | T | P | May–Jun | Forest slopes | PH | MPH | NP | Rhomboid to ovate | LC | Native to China and Taiwan | E |
| Salicaceae Mirb. | ||||||||||||
| Flacourtia indica (Burm.f.) Merr. | RRLH55395 | S | P | Jan–Feb | Mixed forests | PH | NPH | MEG | Oblong | LC | Native to Africa and Asia | N |
| Populus ciliata Wall. ex Royle | RRLH55396 | T | P | May–Jun | Roadsides | PH | MPH | MES | Ovate | LC | Native to the Himalayas | N |
| Xylosma longifolia Clos | RRLH55397 | T | P | Apr–May | Mountain forests | PH | MPH | MES | Elliptic | LC | Native to the Indian subcontinent | N |
| Sapindales Juss. | ||||||||||||
| Sapindaceae Juss. | ||||||||||||
| Aesculus indica (Wall. ex Cambess.) Hook. | RRLH55368 | T | P | Apr–May | Field margins | PH | MPH | MEG | Oblong | LC | Native to the Himalayas | N |
| Acer caesium Wall. ex Brandis | RRLH42643 | T | P | May–Jun | Forest margins | PH | MPH | MEG | Obovate | LC | Native to the Himalayas | N |
| Rutaceae Juss. | ||||||||||||
| Murraya koenigii (L.) Spreng. | RRLH52649 | S | P | Mar–Apr | Mixed forests | PH | NPH | MES | Ovate | NA | Native to India and Sri Lanka | N |
| Aegle marmelos (L.) Corrêa | RRLH52641 | T | P | Jul–Aug | Mixed forests | PH | MPH | MES | Ovate | NT | Native to the Indian subcontinent | N |
| Zanthoxylum armatum DC. | RRLH55226 | T | P | Apr–May | Forest margins | PH | MPH | MIC | Lanceolate | LC | Native to the Himalayas | N |
| Anacardiaceae R.Br. | ||||||||||||
| Cotinus coggygria Scop. | RRLH54912 | T | P | Feb–mar | Forest thickets | PH | MPH | MIC | Elliptic | LC | Native to Eurasia | E |
| Simaroubaceae DC. | ||||||||||||
| Ailanthus altissima (Mill.) Swingle | RRLH55398 | T | P | Apr–May | Mixed forests | PH | MPH | MEG | Ovate | EN | Native to Asia | N |
| Meliaceae Juss. | ||||||||||||
| Melia azedarach L. | RRLH55382 | T | P | Mar–Apr | Field margins | PH | MPH | MIC | Ovate | LC | Native to Asia | N |
| Toona sinensis (Juss.) M.Roem. | RRLH55381 | T | P | May–Jun | Forest margins | PH | MPH | MES | Lanceolate | LC | Native to the Indian subcontinent | N |
| Malvaceae Juss. | ||||||||||||
| Abutilon indicum (L.) Sweet. | RRLH54666 | H | P | July–Oct | Wastelands | CHA | MIC | Ovate | NA | Native to Asia | N | |
| Bombax ceiba L. | RRLH55364 | T | P | Mar–Apr | Field margins | PH | MPH | MES | Oblong | LC | Native to Asia | N |
| Grewia optiva J.R.Drumm. ex Burret | RRLH55380 | T | P | Jun–Jul | Field margins | PH | MPH | MES | Elliptic | NA | Native to the Indian subcontinent | N |
| Malvastrum coromandelianum (L.) Garcke. | RRLH54667 | H | P | May–Jun | Wastelands | GEO | MEG | Ovate | NA | Native to North America | E | |
| Thymelaeaceae Juss. | ||||||||||||
| Daphne papyracea Wall. ex G.Don | RRLH55215 | S | P | Nov–Dec | Forest thickets | PH | NPH | MES | Ovate | NA | Native to Asia | N |
| Brassicales Bromhead | ||||||||||||
| Capparaceae Juss. | ||||||||||||
| Crateva adansonii DC. | RRLH54671 | T | P | Jun–Jul | Roadsides | PH | MPH | NP | Elliptic | LC | Native to Asia | N |
| Caryophyllales Juss. ex Bercht. & J. Presl | ||||||||||||
| Amaranthaceae Juss. | ||||||||||||
| Achyranthes aspera L. | RRLH52653 | H | P | Jun–Aug | Wastelands | CHA | NP | Obovate | NA | Native to South America | E | |
| Aerva sanguinolenta (L.) Blume | RRLH52658 | H | P | April–Jun | Forest margin | CHA | NP | Ovate toelliptic | NA | Native to Asia | N | |
| Dysphania ambrosioides (L.) Mosyakin & Clemants | RRLH54988 | H | A | Mar–Apr | Field margins | THP | MIC | Oblong | NA | Native to South America | E | |
| Polygonaceae Juss. | ||||||||||||
| Persicaria capitata (Buch.-Ham ex D.Don) H.Gross | RRLH54910 | H | P | May–Jun | Forest slopes | CHA | NP | Ovate | NA | Native to Asia | N | |
| Persicaria maculosa Gray | RRLH54928 | H | A | Jun–Jul | Streamside | THP | MES | Lanceolate | LC | Native Asia | N | |
| Rumex dentatus L. | RRLH54700 | H | A | May–Jun | Mountain slopes | THP | MES | Oblong | NA | Native to Asia | N | |
| Rumex hastatus D.Don | RRLH54975 | H | P | Apr–May | Rocky crevices | CHA | MIC | Hastate | NA | Native to the Himalayas | N | |
| IV. ASTERIDS | ||||||||||||
| Santalales | ||||||||||||
| Santalaceae R.Br. | ||||||||||||
| Viscum album L. | RRLH55295 | S | P | Nov–Dec | Parasitic | PH | NPH | MIC | Obovate | NA | Native to Eurasia | E |
| Ericales | ||||||||||||
| Ericaceae Durande | ||||||||||||
| Lyonia ovalifolia (Wall.) Drude. | RRLH55355 | T | P | May–Jun | Forest thickets | PH | MPH | MES | Ovate | LC | Native to the Himalayas | N |
| Rhododendron arboreum Sm. | RRLH55204 | T | P | May–Jun | Forest slopes | PH | MPH | MES | Oblong | LC | Native to Asia | N |
| Primulaceae Batsch ex Borkh. | ||||||||||||
| Lysimachia arvensis (L.) U.Manns & Anderb. | RRLH55387 | H | A | May–Jun | Wastelands | THP | MEG | Ovate | NA | Native to Asia | N | |
| Gentianales Juss. ex Bercht. & J.Presl | ||||||||||||
| Rubiaceae Juss. | ||||||||||||
| Galium asperuloides Edgew. | RRLH55213 | H | P | Apr–May | Mountain slopes | CHA | MIC | Elliptic | NA | Native to the Indian subcontinent | N | |
| Galium aparine L. | RRLH55394 | H | A | Mar–Apr | Forest thickets | THP | MIC | Linear | NA | Native to South America and Eurasia | E | |
| Rubia cordifolia L. | RRLH54625 | H | A | Aug–Sep | Forest margins | THP | NP | Lanceolate | NA | Native to India | N | |
| Apocynaceae Juss. | ||||||||||||
| Holarrhena antidysenterica Wall. | RRLH52631 | T | P | Apr–Jul | Mixed forests | PH | MPH | MEG | Ovate | LC | Native to India | N |
| Boraginaceae Juss. | ||||||||||||
| Cordia dichotoma G. Forst. | RRLH52610 | T | P | Feb–Mar | Field margins | PH | MPH | MEG | Ovate | LC | Native to India | N |
| Solanales Juss. ex Bercht. & J.Presl | ||||||||||||
| Convolvulaceae Juss. | ||||||||||||
| Argyreia nervosa (Burm.f.) Bojer | RRLH55363 | S | P | Mar–Apr | Mixed forests | PH | NPH | MES | Cordate | NA | Native to the Indian subcontinent | N |
| Solanaceae Juss. | ||||||||||||
| Solanum americanum Mill. | RRLH54619 | H | A | Nov–Dec | Wastelands | THP | MES | Ovate | NA | Native to North America | E | |
| Solanum virginianum L. | RRLH54927 | H | A | Oct–Dec | Moist places | THP | MES | Ovate to oblong | NA | Native to the Indian subcontinent | N | |
| Solanum xanthocarpum Schrad. | RRLH52367 | H | P | Nov–Dec | Moist places | THP | MES | Ovate | LC | Native to the Indian subcontinent | N | |
| Lamiales Bromhead | ||||||||||||
| Oleaceae Hoffmanns. & Link | ||||||||||||
| Syringa emodi Wall. ex Royle | RRLH55386 | T | P | May–Jun | Mixed forests | PH | MPH | MES | Ovate | NA | Native to the Himalayas | N |
| Plantaginaceae Juss. | ||||||||||||
| Plantago lanceolata L. | RRLH55365 | H | P | May–Jun | Wastelands | HCP | MES | Lanceolate | VU | Native to Eurasia to Asia | N | |
| Acanthaceae Juss. | ||||||||||||
| Barleria cristata L. | RRLH52605 | H | P | May–Jun | Forest slopes | CHA | MES | Elliptic | NA | Native to Asia | N | |
| Dicliptera bupleuroides Nees | RRLH52611 | H | A | Jun–Jul | Roadsides | THP | NP | Ovate | NA | Native to Asia | N | |
| Justicia adhatoda L. | RRLH52653 | H | P | Jan–Feb | Roadsides | PH | MES | Ovate toelliptic | NA | Native to the Indian subcontinent | N | |
| Lepidagathis cuspidata Nees | RRLH52619 | H | P | Mar–May | Forest thickets | CHA | MES | Elliptic | NA | Native to India | N | |
| Rungia pectinata (L.) Nees | RRLH54924 | H | P | Nov–Dec | Wastelands | THP | MES | Oblong | NA | Native to Asia | N | |
| Strobilanthes wallichii Nees | RRLH54997 | S | P | Jun–Jul | Open forests | PH | NPH | NP | Elliptic | NA | Native to the Himalayas | N |
| Lamiaceae Martinov | ||||||||||||
| Colebrookea oppositifolia Sm. | RRLH54908 | S | P | Jan–Mar | Forest thickets | PH | NPH | NP | Oblong | NA | Native to the Indian subcontinent | N |
| Isodon japonicus (Burm.f.) H.Hara | RRLH55224 | S | P | Jul–Aug | Forest thickets | PH | NPH | NP | Ovate | NA | Native to Asia | N |
| Isodon rugosus (Wall. ex Benth.) Codd | RRLH55225 | S | P | Jul–Aug | Forest thickets | PH | NPH | MEG | Ovate | NA | Native to Asia | N |
| Leucas ciliata Benth. | RRLH54903 | H | P | Jul–Oct | Roadsides | THP | NP | Lanceolate | NA | Native to Asia | N | |
| Scutellaria discolor Wall. ex Benth. | RRLH55289 | H | P | Jun–Jul | Forest margins | CHA | MES | Elliptic to ovate | NA | Native to the Himalayas | N | |
| Vitex negundo L. | RRLH55378 | S | P | Apr–May | Forest thickets | PH | NPH | NP | Lanceolate | LC | Native to Asia and Africa | N |
| Scrophulariaceae Juss. | ||||||||||||
| Buddleja paniculata Wall. | RRLH55369 | S | P | Mar–Apr | Forest thickets | PH | NPH | MES | Elliptic | NA | Native to Asia | N |
| Asterales Link | ||||||||||||
| Asteraceae Bercht. & J.Presl | ||||||||||||
| Ageratum conyzoides L. | RRLH55357 | H | A | Jan–Dec | Field margins | THP | NP | Ovate | NA | Native to South America | E | |
| Bidens biternata (Lour.) Merr. & Sherff | RRLH54650 | H | A | Sep–Oct | Roadsides | THP | NP | Ovate | NA | Native to the tropical and subtropical Old World | E | |
| Elephantopus scaber L. | RRLH55258 | H | A | Jul–Aug | Forest margins | THP | MES | Oblanceolate | NA | Native to the Indian subcontinent | N | |
| Galinsoga parviflora Cav. | RRLH54630 | H | A | Jul–Aug | Field margins | THP | MIC | Elliptic | NA | Native to South America | E | |
| Gynura angulosa (Wall.) DC. | RRLH54609 | H | P | Sep–Oct | Forest slopes | CHA | NP | Obovate | NA | Native to Asia | N | |
| Jacobaea nudicaulis (Buch.-Ham. ex D.Don) B.Nord. | RRLH55277 | H | A | Mar–Apr | Grassy slopes | THP | NP | Oblong | NA | Native to Asia | N | |
| Oreoseris gossypina (Royle) X.D.Xu & V.A.Funk | RRLH55385 | H | P | May–Jun | Rocky slopes | CHA | MES | Oblanceolate | NA | Native to the foothills of the Himalayas | N | |
| Parthenium hysterophorus L. | RRLH54947 | H | A | Apr–May | Wastelands | THP | NP | Ovate toelliptic | NA | Native to South America | E | |
| Blainvillea acmella (L.) Philipson | RRLH52646 | H | A | Mar–May | Moist places | THP | MES | Elliptic | NA | Native to Brazil | E | |
| Apiales Nakai | ||||||||||||
| Araliaceae Juss. | ||||||||||||
| Hedera nepalensis K.Koch | RRLH55356 | S | P | Oct–Nov | Mixed forests | PH | NPH | MIC | Lanceolate | NA | Native to Asia | N |
| GYMNOSPERMS | ||||||||||||
| Pinales Gorozh. | ||||||||||||
| Pinaceae Spreng. ex F.Rudolphi | ||||||||||||
| Cedrus deodara (Roxb. ex D.Don) G.Don | RRLH55222 | T | P | May–Jun | Coniferous forests | PH | MPH | MES | Linear | LC | Native to the Himalayas | N |
| Juniperus recurva Buch.-Ham. ex D.Don | RRLH55225 | T | P | Apr–May | Coniferous forests | PH | MPH | LEP | Linear | LC | Native to the Himalayas | N |
| Pinus roxburghii Sarg. | RRLH55223 | T | P | Sep–Oct | Coniferous forests | PH | MPH | LEP | Linear | LC | Native to the Himalayas | N |
| Pinus wallichiana A.B.Jacks. | RRLH55224 | T | P | May–Jun | Coniferous forests | PH | MPH | LEP | Linear | LC | Native to the Himalayas | N |
| Taxaceae Gray | ||||||||||||
| Taxus baccata L. | RRLH55293 | T | P | Aug-Dec | Coniferous forests | PH | NPH | LEP | Linear | LC | Native to the Himalayas | N |
| LYCOPHYTES AND FERNS Polypodiales | ||||||||||||
| Dennstaedtiaceae Pic.Serm. | ||||||||||||
| Microlepia nepalensis (Spreng.) Fraser-Jenk., Kandel & Pariyar | RRLH54681 | H | A | May–Jun | Forest slopes | THP | MIC | Ovate | NA | Native to Asia | N | |
| Pteridaceae E.D.M. Kirchn. | ||||||||||||
| Adiantum capillus-veneris L. | RRLH54676 | H | A | May–Jun | Shady places | THP | NP | Elliptic | LC | Native to the Indian subcontinent | N | |
| Cheilanthes subvillosa Hook. | RRLH54679 | H | A | Mar–Apr | Moist places | THP | NP | Lanceolate | NA | Native to the Indian subcontinent | N | |
| Onychium japonicum (Thunb.) Kunze | RRLH54682 | H | A | May–Jun | Moist places | THP | MIC | Ovate | NA | Native to Asia | N | |
| Polystichum polyblepharum (Roem. ex Kunze) C.Presl | RRLH54683 | H | P | Mar–Apr | Shady places | HCP | LEP | Lanceolate | NA | Native to North and South America | E | |
| Pteris cretica L. | RRLH54684 | H | P | Mar–Apr | Forest margins | HCP | MES | Obovate | NA | Native to Africa and Eurasia | E | |
| Pteris vittata L. | RRLH54685 | H | P | Mar–Apr | Forest margins | HCP | MES | Elliptic | LC | Native to Asia | N | |
| Aspleniaceae Newman | ||||||||||||
| Asplenium adiantum–nigrum L. | RRLH54677 | H | A | May–Jun | Rocky crevices | THP | LEP | Ovate | NA | Native to Eurasia | E | |
| Asplenium dalhousiae Hook. | RRLH54678 | H | A | May–Jun | Rocky surfaces | THP | NP | Ovate | NA | Native to India | N | |
| Selaginellaceae Willk. | ||||||||||||
| Selaginella eurynota A.Br. | RRLH54686 | H | A | Mar–Apr | Shady places | THP | LEP | Elliptic to ovate | NA | Native to Asia | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Singh, B.; Surmal, O.; Bhat, M.N.; Singh, B.; Musarella, C.M. Fragmented Forest Patches in the Indian Himalayas Preserve Unique Components of Biodiversity: Investigation of the Floristic Composition and Phytoclimate of the Unexplored Bani Valley. Sustainability 2021, 13, 6063. https://0-doi-org.brum.beds.ac.uk/10.3390/su13116063
Singh S, Singh B, Surmal O, Bhat MN, Singh B, Musarella CM. Fragmented Forest Patches in the Indian Himalayas Preserve Unique Components of Biodiversity: Investigation of the Floristic Composition and Phytoclimate of the Unexplored Bani Valley. Sustainability. 2021; 13(11):6063. https://0-doi-org.brum.beds.ac.uk/10.3390/su13116063
Chicago/Turabian StyleSingh, Sumit, Bikarma Singh, Opender Surmal, Mudasir Nazir Bhat, Bishander Singh, and Carmelo Maria Musarella. 2021. "Fragmented Forest Patches in the Indian Himalayas Preserve Unique Components of Biodiversity: Investigation of the Floristic Composition and Phytoclimate of the Unexplored Bani Valley" Sustainability 13, no. 11: 6063. https://0-doi-org.brum.beds.ac.uk/10.3390/su13116063