
Combined Use of Endophytic Bacteria and Pre-Sowing Treatment of Thiamine Mitigates the Adverse Effects of Drought Stress in Wheat (Triticum aestivum L.) Cultivars

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seeds Sterilization
2.2. Seeds Inoculation and Transplantation
2.3. Experimental Design
2.4. Treatment Plan
2.5. Drought Stress
2.6. Harvesting and Data Collection
2.7. Physiological Attributes
2.8. Oxidative Stress Indicators
2.9. Electrolyte Leakage
2.10. Antioxidant Enzymes Activities
2.11. Estimation of Proline, Sugars and Non-Enzymatic Antioxidants
2.12. Statistical Analysis
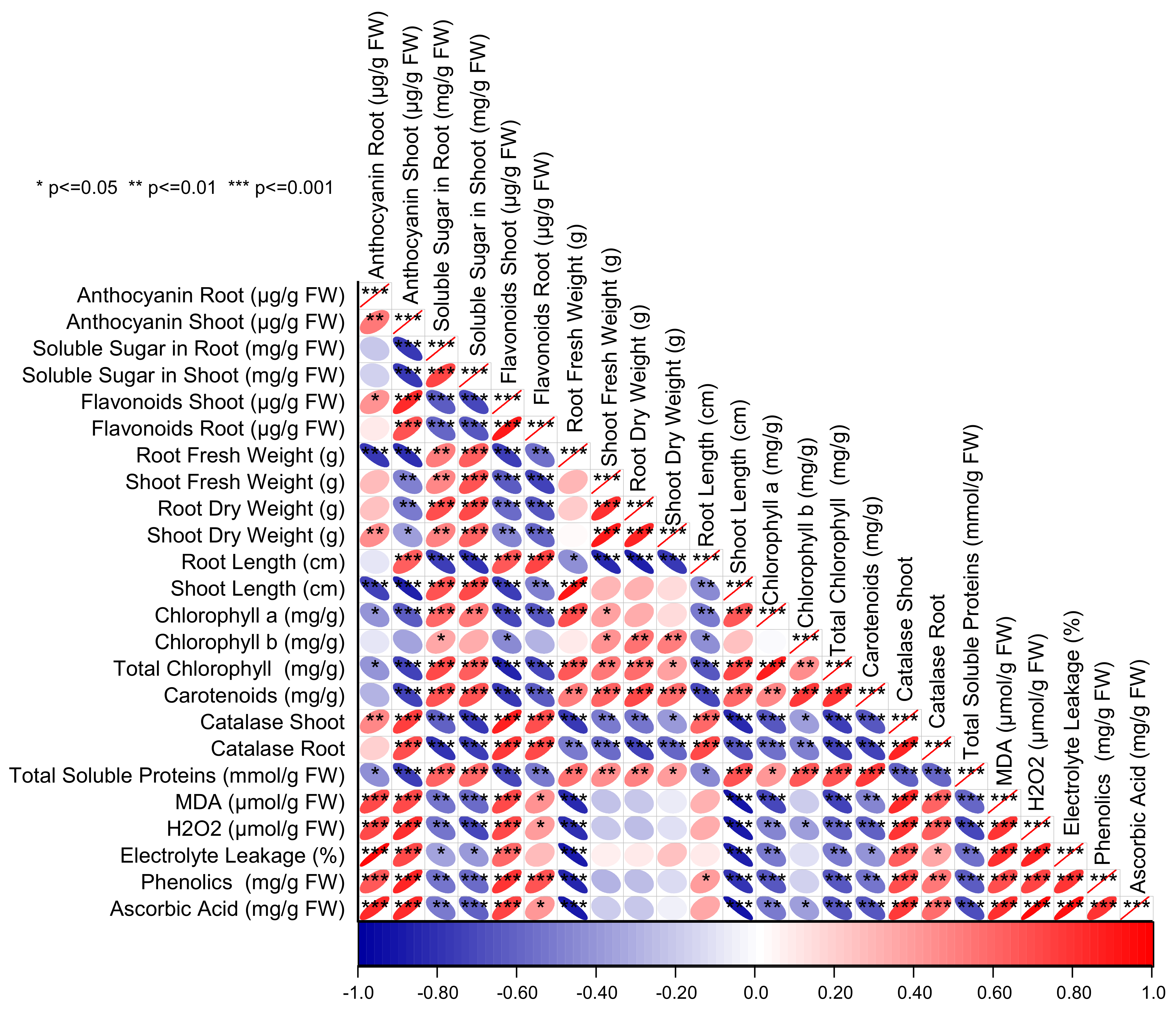
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zafar-ul-hye, M.; Naeem, M.; Danish, S.; Khan, M.J.; Fahad, S.; Datta, R.; Brtnicky, M.; Kintl, A.; Hussain, M.S.; El-esawi, M.A. Effect of cadmium-tolerant rhizobacteria on growth attributes and chlorophyll contents of bitter gourd under cadmium toxicity. Plants 2020, 9, 1386. [Google Scholar] [CrossRef] [PubMed]
- Danish, S.; Tahir, F.A.; Rasheed, M.K.; Ahmad, N.; Ali, M.A.; Kiran, S.; Younis, U.; Irshad, I.; Butt, B. Effect of foliar application of Fe and banana peel waste biochar on growth, chlorophyll content and accessory pigments synthesis in spinach under chromium (IV) toxicity. Open Agric. 2019, 4, 381–390. [Google Scholar] [CrossRef]
- Radziemska, M.; Gusiatin, Z.M.; Cydzik-Kwiatkowska, A.; Cerdà, A.; Pecina, V.; Bęś, A.; Datta, R.; Majewski, G.; Mazur, Z.; Dzięcioł, J.; et al. Insight into metal immobilization and microbial community structure in soil from a steel disposal dump that was phytostabilized with composted, pyrolyzed or gasified wastes. Chemosphere 2021, 272, 129576. [Google Scholar] [CrossRef] [PubMed]
- Zafar-ul-Hye, M.; Tahzeeb-ul-Hassan, M.; Abid, M.; Fahad, S.; Brtnicky, M.; Dokulilova, T.; Datta, R.; Danish, S. Potential role of compost mixed biochar with rhizobacteria in mitigating lead toxicity in spinach. Sci. Rep. 2020, 10, 12159. [Google Scholar] [CrossRef] [PubMed]
- Zafar-Ul-hye, M.; Naeem, M.; Danish, S.; Fahad, S.; Datta, R.; Abbas, M.; Rahi, A.A.; Brtnicky, M.; Holátko, J.; Tarar, Z.H.; et al. Alleviation of cadmium adverse effects by improving nutrients uptake in bitter gourd through cadmium tolerant rhizobacteria. Environments 2020, 7, 54. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Shahjahan, A.; Danish, S.; Abid, M.; Qayyum, M.F. Mitigation of cadmium toxicity induced stress in wheat by ACC-deaminase containing PGPR isolated from cadmium polluted wheat rhizosphere. Pakistan J. Bot. 2018, 50, 1727–1734. [Google Scholar]
- Ahmed, N.; Ahsen, S.; Ali, M.A.; Hussain, M.B.; Hussain, S.B.; Rasheed, M.K.; Butt, B.; Irshad, I.; Danish, S. Rhizobacteria and silicon synergy modulates the growth, nutrition and yield of mungbean under saline soil. Pakistan J. Bot. 2020, 52, 9–15. [Google Scholar] [CrossRef]
- Rafiullah; Jamal Khan, M.; Muhammad, D.; Fahad, S.; Adnan, M.; Wahid, F.; Alamri, S.; Khan, F.; Muhammad Dawar, K.; Irshad, I.; et al. Phosphorus nutrient management through synchronization of application methods and rates in wheat and maize crops. Plants 2020, 9, 1389. [Google Scholar] [CrossRef]
- Wahid, F.; Fahad, S.; Danish, S.; Adnan, M.; Yue, Z.; Saud, S.; Siddiqui, M.H.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Sustainable Management with Mycorrhizae and Phosphate Solubilizing Bacteria for Enhanced Phosphorus Uptake in Calcareous Soils. Agriculture 2020, 10, 334. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M.; Fahad, S.; Saud, S.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Drought stress alleviation by ACC deaminase producing Achromobacter xylosoxidans and Enterobacter cloacae, with and without timber waste biochar in maize. Sustainability 2020, 12, 6286. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Danish, S.; Zafar-Ul-Hye, M.; Hussain, S.; Riaz, M.; Qayyum, M.F. Mitigation of drought stress in maize through inoculation with drought tolerant ACC deaminase containing PGPR under axenic conditions. Pakistan J. Bot. 2020, 52. [Google Scholar] [CrossRef]
- Gilani, M.; Danish, S.; Ahmed, N.; Rahi, A.A.; Akrem, A.; Younis, U.; Irshad, I.; Iqbal, R.K. Mitigation of drought stress in spinach using individual and combined applications of salicylic acid and potassium. Pakistan J. Bot. 2020, 52, 1505–1513. [Google Scholar] [CrossRef]
- Janjua, P.Z.; Samad, G.; Khan, N. Climate change and wheat production in Pakistan: An autoregressive distributed lag approach. NJAS Wageningen J. Life Sci. 2014, 68, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Izhar Shafi, M.; Adnan, M.; Fahad, S.; Wahid, F.; Khan, A.; Yue, Z.; Danish, S.; Zafar-ul-Hye, M.; Brtnicky, M.; Datta, R. Application of Single Superphosphate with Humic Acid Improves the Growth, Yield and Phosphorus Uptake of Wheat (Triticum aestivum L.) in Calcareous Soil. Agronomy 2020, 10, 1224. [Google Scholar] [CrossRef]
- Camilios-Neto, D.; Bonato, P.; Wassem, R.; Tadra-Sfeir, M.Z.; Brusamarello-Santos, L.C.C.; Valdameri, G.; Donatti, L.; Faoro, H.; Weiss, V.A.; Chubatsu, L.S.; et al. Dual RNA-seq transcriptional analysis of wheat roots colonized by Azospirillum brasilense reveals up-regulation of nutrient acquisition and cell cycle genes. BMC Genom. 2014, 15, 378. [Google Scholar] [CrossRef] [Green Version]
- Creus, C.M.; Sueldo, R.J.; Barassi, C.A. Water relations and yield in Azospirillum-inoculated wheat exposed to drought in the field. Can. J. Bot. 2004, 82, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jiménez, M.; Tallón, C.I.; Pérez-Tornero, O. Inducing mutations in Citrus spp.: Sensitivity of different sources of plant material to gamma radiation. Appl. Radiat. Isot. 2020, 157, 109030. [Google Scholar] [CrossRef]
- Amir, K.; Hussain, S.; Shuaib, M.; Hussain, F.; Urooj, Z.; Khan, W.M.; Zeb, U.; Ali, K.; Zeb, M.A.; Hussain, F. Effect of gamma irradiation on OKRA (Abelmoschus esculentus L.). Shengtai Xuebao Acta Ecol. Sin. 2018, 38, 368–373. [Google Scholar] [CrossRef]
- Carvalho, T.L.G.; Ballesteros, H.G.F.; Thiebaut, F.; Ferreira, P.C.G.; Hemerly, A.S. Nice to meet you: Genetic, epigenetic and metabolic controls of plant perception of beneficial associative and endophytic diazotrophic bacteria in non-leguminous plants. Plant Mol. Biol. 2016, 90, 561–574. [Google Scholar] [CrossRef]
- Jabeen, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Thiamin stimulates growth and secondary metabolites in turnip (Brassica rapa L.) leaf and root under drought stress. Physiol. Plant. 2020. [Google Scholar] [CrossRef]
- Ors, S.; Suarez, D.L. Spinach biomass yield and physiological response to interactive salinity and water stress. Agric. Water Manag. 2017, 190, 31–41. [Google Scholar] [CrossRef]
- Hamada, A.; Al-Hakimi, A. Exogenous ascorbic acid or thiamine increases the resistance of sunflower and maize plants to salt stress. Acta Agron. Hungarica 2009, 57, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.; Shrewry, P.R. Wheat: Chemistry and Technology, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2009; ISBN 9780128104545. [Google Scholar]
- Arshad, M.; Amjath-Babu, T.S.; Aravindakshan, S.; Krupnik, T.J.; Toussaint, V.; Kächele, H.; Müller, K. Climatic variability and thermal stress in Pakistan’s rice and wheat systems: A stochastic frontier and quantile regression analysis of economic efficiency. Ecol. Indic. 2018, 89, 496–506. [Google Scholar] [CrossRef]
- Miedaner, T.; Juroszek, P. Climate change will influence disease resistance breeding in wheat in Northwestern Europe. Theor. Appl. Genet. 2021. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat. Bot. 1982, 12, 345–354. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Sakharov, I.Y.; Ardila, G.B. Variations of peroxidase activity in cocoa (Theobroma cacao L.) beans during their ripening, fermentation and drying. Food Chem. 1999, 65, 51–54. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Lewis, C.E.; Walker, J.R.L.; Lancaster, J.E.; Sutton, K.H. Determination of anthocyanins, flavonoids and phenolic acids in potatoes. I: Coloured cultivars of Solanum tuberosum L. J. Sci. Food Agric. 1998, 77, 45–57. [Google Scholar] [CrossRef]
- Bray, H.G.; Thorpe, W.V. Analysis of phenolic compounds of interest in metabolism. Methods Biochem. Anal. 1954, 1, 27–52. [Google Scholar]
- Pękal, A.; Pyrzynska, K. Evaluation of Aluminium Complexation Reaction for Flavonoid Content Assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Azuma, K.; Nakayama, M.; Koshioka, M.; Ippoushi, K.; Yamaguchi, Y.; Kohata, K.; Yamauchi, Y.; Ito, H.; Higashio, H. Phenolic antioxidants from the leaves of Corchorus olitorius L. J. Agric. Food Chem. 1999, 47, 3963–3966. [Google Scholar] [CrossRef] [PubMed]
- Steel, R.G.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; McGraw Hill Book International Co.: Singapore, 1997. [Google Scholar]
- OriginLab Corporation. OriginPro; OriginLab: Northampton, MA, USA, 2021. [Google Scholar]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeiger, E.; Taiz, L. Plant Physiology; Sinauer Associates Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Paul, S.; Aggarwal, C.; Manjunatha, B.; Rathi, M.S. Characterization of osmotolerant rhizobacteria for plant growth promoting activities in vitro and during plant-microbe association under osmotic stress. Indian J. Exp. Biol. 2018, 56, 582–589. [Google Scholar]
- Glick, B.; Penrose, D.; Li, J. A Model For the Lowering of Plant Ethylene Concentrations by Plant Growth-promoting Bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Matile, P.; Schellenberg, M.; Vicentini, F. Planta Localization of chlorophyllase in the chloroplast envelope. Planta 1997, 201, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Wang, H.; Xie, J. Thiamin (vitamin B1) biosynthesis and regulation: A rich source of antimicrobial drug targets? Int. J. Biol. Sci. 2011, 7, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.; Shabbir, G.; Ahmed, M.; Shah, K.N. Phenotyping for drought resistance in bread wheat using physiological and biochemical traits. Sci. Total Environ. 2020, 729, 139082. [Google Scholar] [CrossRef] [PubMed]
- Kishor, P.B.K.; Rajesh, K.; Reddy, P.S.; Seiler, C.; Sreenivasulu, N. Drought stress tolerance mechanisms in barley and its relevance to cereals. In Biotechnology in Agriculture and Forestry; Kumlehn, J., Stein, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 69, pp. 161–179. [Google Scholar]
- Shan, C.; Zhang, S.; Ou, X. The roles of H2S and H2O2 in regulating AsA-GSH cycle in the leaves of wheat seedlings under drought stress. Protoplasma 2018, 255, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Kleinenkuhnen, N.; Büchel, F.; Gerlich, S.C.; Kopriva, S.; Metzger, S. A novel method for identification and quantification of sulfated flavonoids in plants by neutral loss scan mass spectrometry. Front. Plant Sci. 2019, 10, 885. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
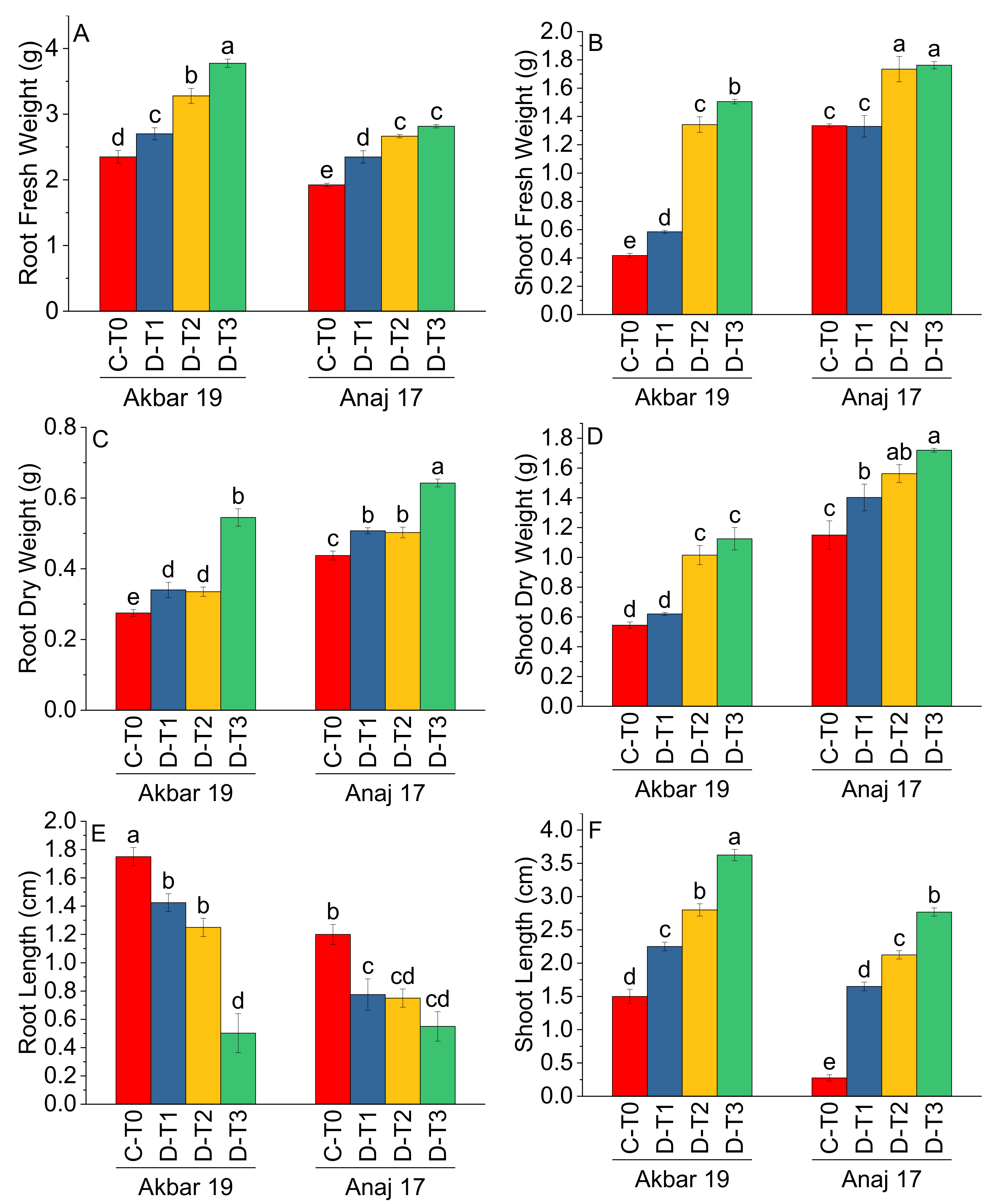
| Treatment | Main Effect of Treatments | |||||
|---|---|---|---|---|---|---|
| RFW (g) | SFW (g) | RDW (g) | SDW (g) | RL (cm) | SL (cm) | |
| C-T0 | 2.14 d | 0.88 b | 0.35 c | 0.85 d | 1.48 a | 3.20 a |
| D-T1 | 2.52 c | 0.96 b | 0.42 b | 1.01 c | 1.10 b | 2.46 b |
| D-T2 | 2.97 b | 1.54 a | 0.42 b | 1.29 b | 1.00 b | 1.95 c |
| D-T3 | 3.30 a | 1.63 a | 0.59 a | 1.42 a | 0.53 c | 0.89 d |
| Variety | Main Effect of Variety | |||||
| Akbar 19 | 3.03 a | 0.96 b | 0.37 b | 0.83 b | 1.23 a | 2.54 a |
| Anaj 17 | 2.44 b | 1.54 a | 0.52 a | 1.46 a | 0.82 b | 1.70 b |
| ANOVA | ||||||
| Grand Mean | 2.73 | 1.25 | 0.44 | 1.14 | 1.03 | 2.12 |
| CV | 5.47 | 7.65 | 6.89 | 10.94 | 17.39 | 7.12 |
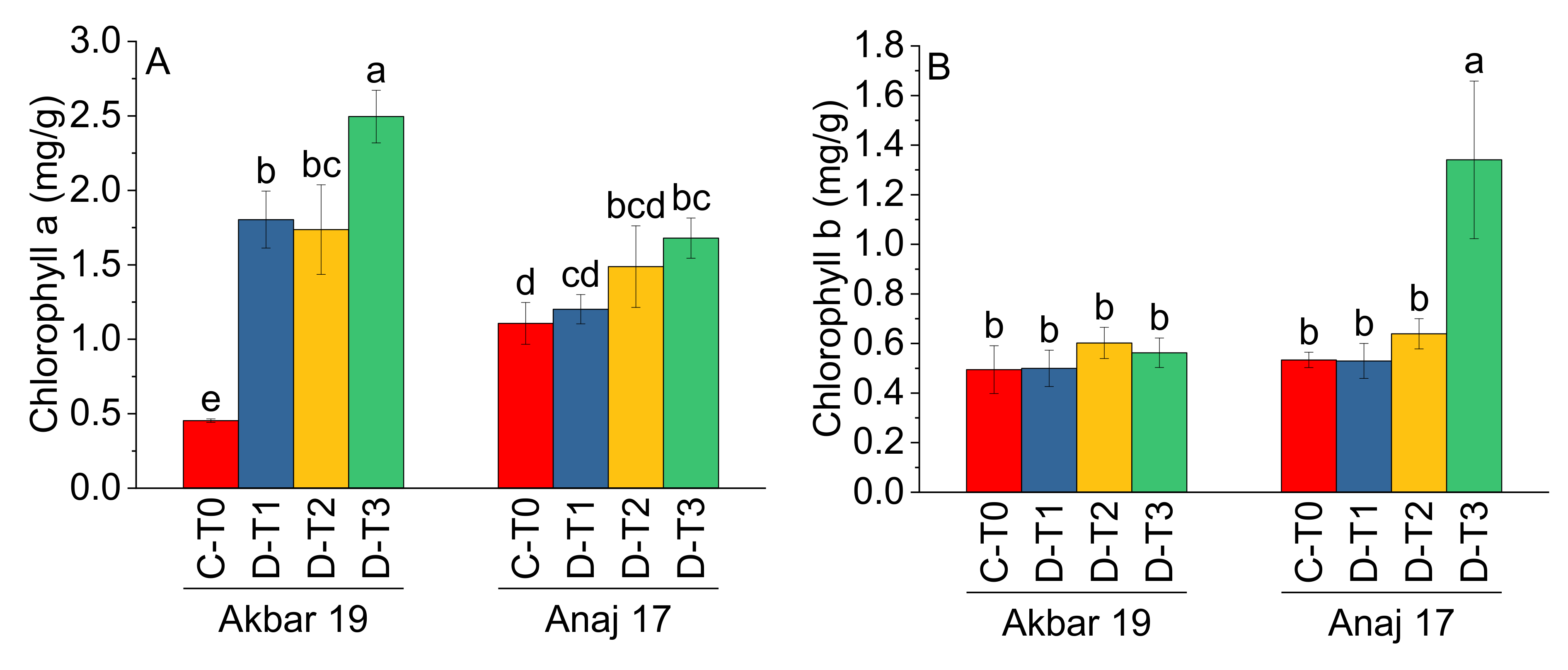
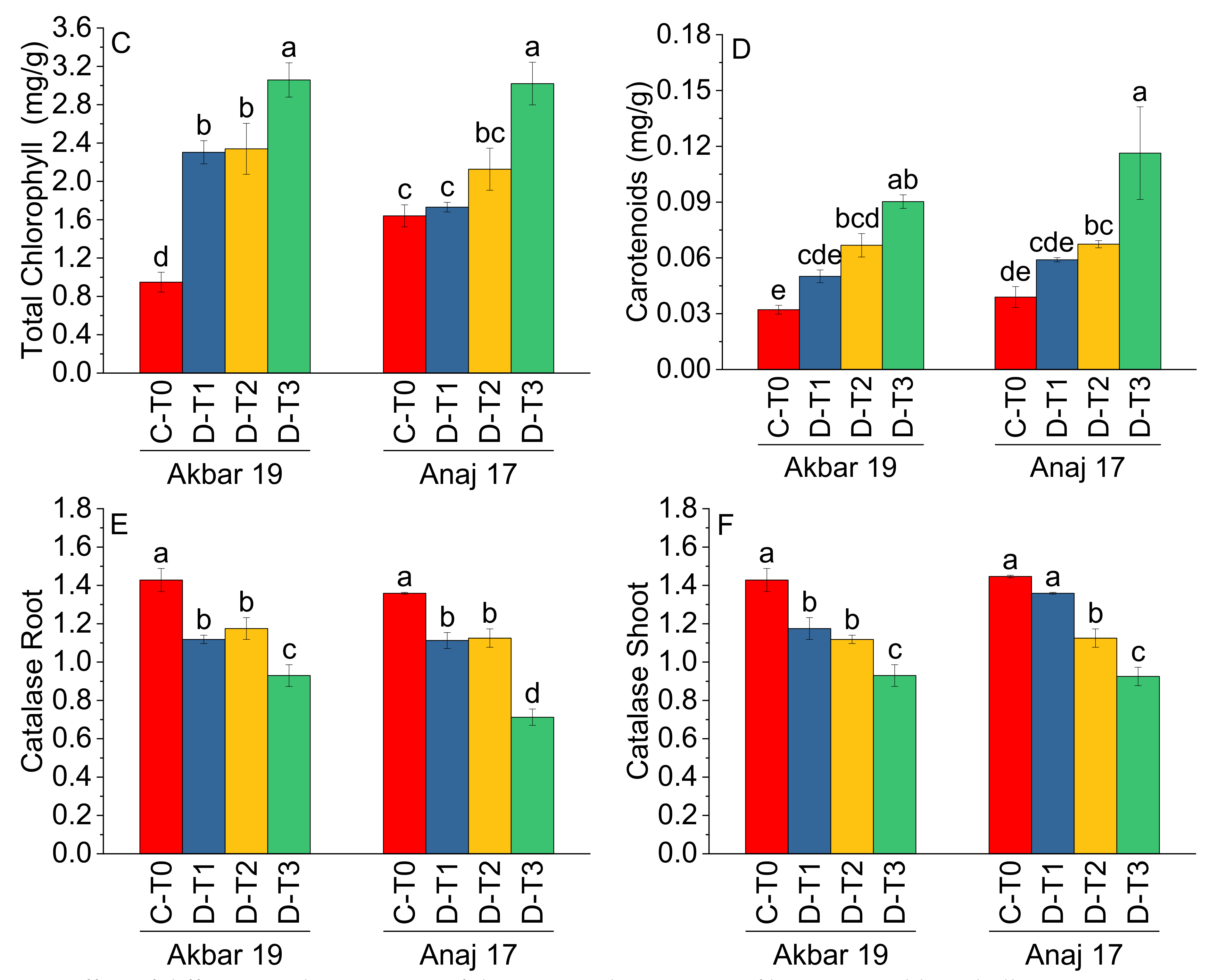
| Treatment | Main Effect of Treatments | |||||
|---|---|---|---|---|---|---|
| Chla (mg/g) | Chlb (mg/g) | TChl (mg/g) | Ca (mg/g) | CATR | CATS | |
| C-T0 | 0.78 c | 0.51 b | 1.29 c | 0.036 c | 1.39 a | 1.43 a |
| D-T1 | 1.50 b | 0.51 b | 2.02 b | 0.055 bc | 1.50 b | 1.27 b |
| D-T2 | 1.61 b | 0.62 b | 2.23 b | 0.067 b | 1.11 b | 1.12 c |
| D-T3 | 2.08 a | 0.95 a | 3.04 a | 0.103 a | 0.82 c | 0.93 d |
| Variety | Main Effect of Variety | |||||
| Akbar 19 | 1.62 a | 0.54 b | 2.16 a | 0.070 a | 1.16 a | 1.62 a |
| Anaj 17 | 1.37 a | 0.76 a | 2.12 a | 0.060 a | 1.07 b | 1.21 a |
| ANOVA | ||||||
| Grand Mean | 1.50 | 0.65 | 2.15 | 0.07 | 1.12 | 1.19 |
| CV | 25.05 | 39.65 | 16.18 | 29.29 | 8.09 | 7.33 |
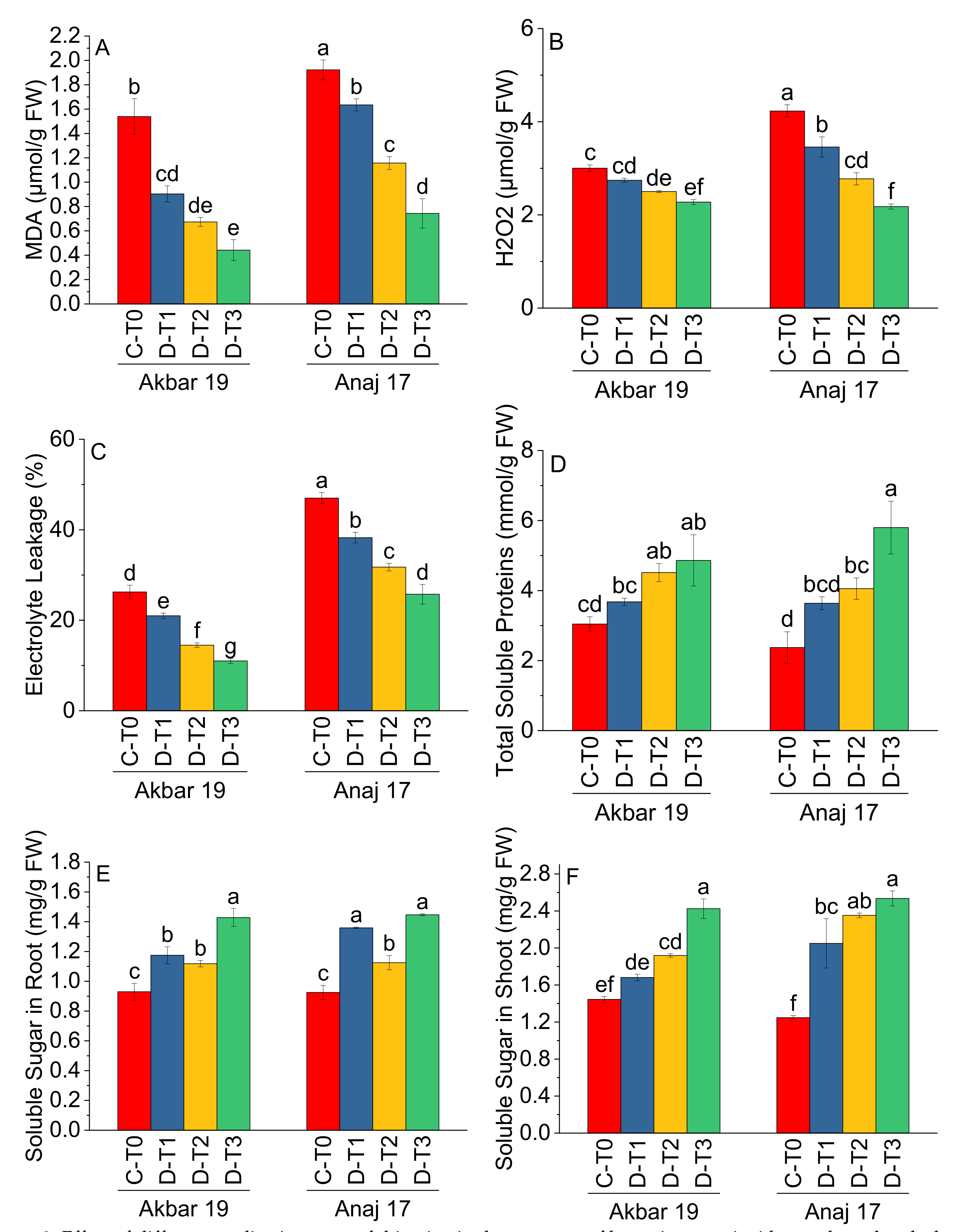
| Treatment | Main Effect of Treatments | |||||
|---|---|---|---|---|---|---|
| MDA (μmol/g FW) | H2O2 (μmol/g FW) | EL (%) | TSP (mmol/g FW) | SSR (mg/g FW) | SSS (mg/g FW) | |
| C-T0 | 1.73 a | 3.61 a | 36.62 a | 2.71 c | 0.93 d | 1.34 d |
| D-T1 | 1.27 b | 3.10 b | 29.62 b | 3.66 b | 1.12 c | 1.87 c |
| D-T2 | 0.91 c | 2.64 c | 23.13 c | 4.29 b | 1.27 b | 2.14 b |
| D-T3 | 0.59 d | 2.23 d | 18.38 d | 5.32 a | 1.44 a | 2.48 a |
| Variety | Main Effect of Variety | |||||
| Akbar 19 | 0.89 b | 2.63 b | 18.18 b | 4.02 a | 1.16 a | 1.87 b |
| Anaj 17 | 1.36 a | 3.16 a | 35.68 a | 3.97 a | 1.21 a | 2.05 a |
| ANOVA | ||||||
| Grand Mean | 1.13 | 2.89 | 26.99 | 4.00 | 1.19 | 1.96 |
| CV | 15.48 | 7.50 | 8.90 | 22.03 | 7.33 | 11.00 |
| Treatment | Main Effect of Treatments | |||||
|---|---|---|---|---|---|---|
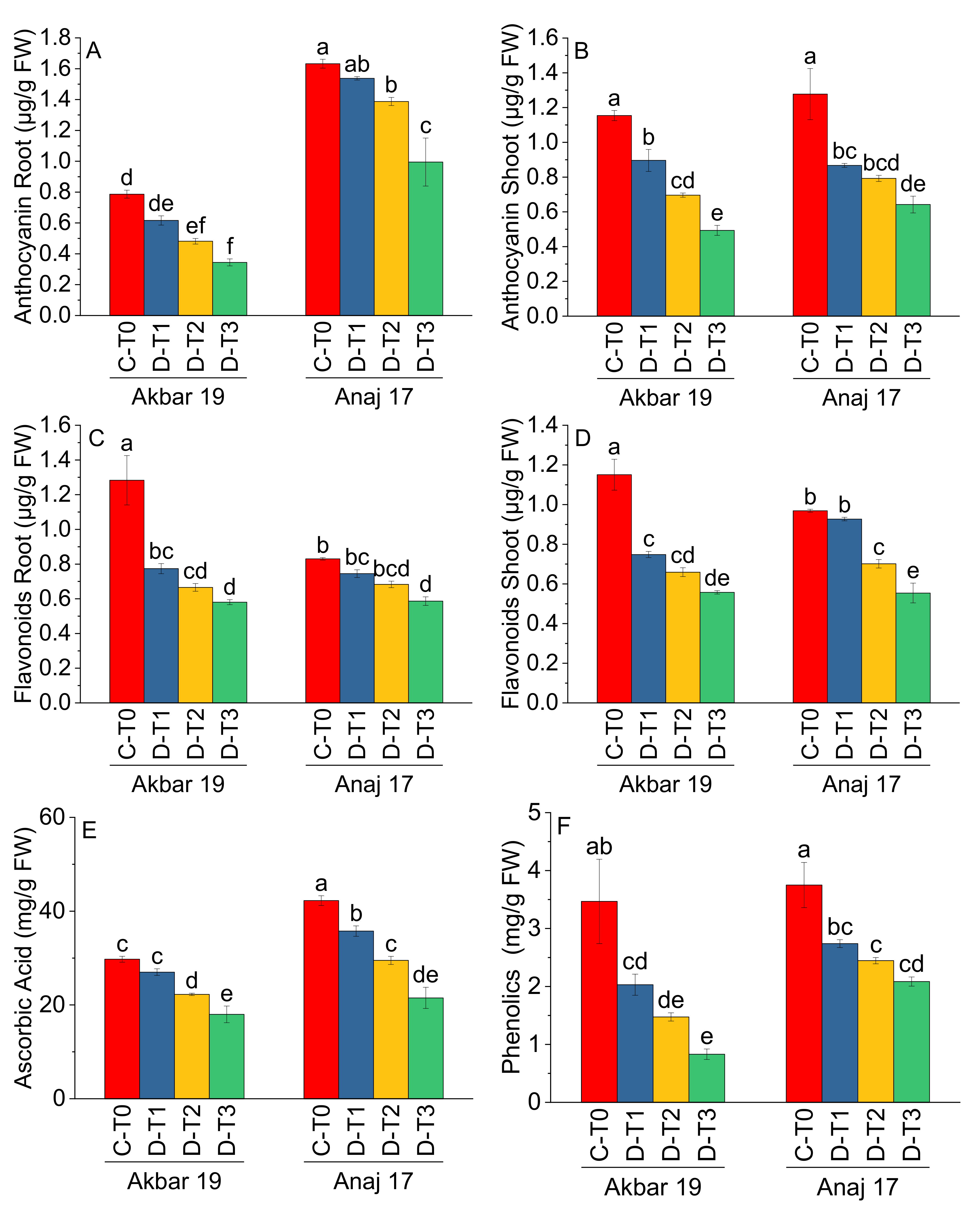
| AnthR (μg/g FW) | AnthS (μg/g FW) | FR (μg/g FW) | FS (μg/g FW) | AsA (mg/g FW) | Phe (mg/g FW) | |
| C-T0 | 1.21 a | 1.22 a | 1.06 a | 1.06 a | 36.00 a | 3.61 a |
| D-T1 | 1.08 b | 0.88 b | 0.76 b | 0.84 b | 31.38 b | 2.38 b |
| D-T2 | 0.93 c | 0.74 c | 0.67 bc | 0.68 c | 25.88 c | 1.96 bc |
| D-T3 | 0.67 d | 0.57 d | 0.58 c | 0.56 d | 19.75 d | 1.45 c |
| Variety | Main Effect of Variety | |||||
| Akbar 19 | 0.56 b | 0.81 a | 0.83 a | 0.78 a | 24.25 b | 1.95 b |
| Anaj 17 | 1.39 a | 0.89 a | 0.71 b | 0.79 a | 32.25 a | 2.76 a |
| ANOVA | ||||||
| Grand Mean | 0.97 | 0.85 | 0.77 | 0.78 | 28.25 | 2.35 |
| CV | 12.23 | 14.43 | 14.03 | 9.00 | 8.76 | 25.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amjad, S.F.; Mansoora, N.; Yaseen, S.; Kamal, A.; Butt, B.; Matloob, H.; Alamri, S.A.M.; Alrumman, S.A.; Eid, E.M.; Shahbaz, M. Combined Use of Endophytic Bacteria and Pre-Sowing Treatment of Thiamine Mitigates the Adverse Effects of Drought Stress in Wheat (Triticum aestivum L.) Cultivars. Sustainability 2021, 13, 6582. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126582
Amjad SF, Mansoora N, Yaseen S, Kamal A, Butt B, Matloob H, Alamri SAM, Alrumman SA, Eid EM, Shahbaz M. Combined Use of Endophytic Bacteria and Pre-Sowing Treatment of Thiamine Mitigates the Adverse Effects of Drought Stress in Wheat (Triticum aestivum L.) Cultivars. Sustainability. 2021; 13(12):6582. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126582
Chicago/Turabian StyleAmjad, Syeda Fasiha, Nida Mansoora, Samia Yaseen, Afifa Kamal, Beenish Butt, Humera Matloob, Saad A. M. Alamri, Sulaiman A. Alrumman, Ebrahem M. Eid, and Muhammad Shahbaz. 2021. "Combined Use of Endophytic Bacteria and Pre-Sowing Treatment of Thiamine Mitigates the Adverse Effects of Drought Stress in Wheat (Triticum aestivum L.) Cultivars" Sustainability 13, no. 12: 6582. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126582