
Alleviation of Salt Stress in Wheat Seedlings via Multifunctional Bacillus aryabhattai PM34: An In-Vitro Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Isolation and Fungal Strain Acquisition
2.2. Plant Growth-Promoting Traits of Strain PM34
2.3. Extracellular Enzymes, ACC Deaminase, and Exopolysaccharide (EPS) Production by Strain PM34
2.4. Molecular Profiling (Amplification of 16S rRNA, nifH, ituC, and Acds Gene
2.5. Abiotic and Biotic Stress Tolerance Assay
2.6. In Vitro Experiment
2.7. Statistical Analysis
3. Results
3.1. Biochemical and Molecular Characterization of the Strain PM34
3.2. Abiotic and Biotic Stress Tolerance Assay
3.3. Wheat Germination and Growth Responses under Salt Stress
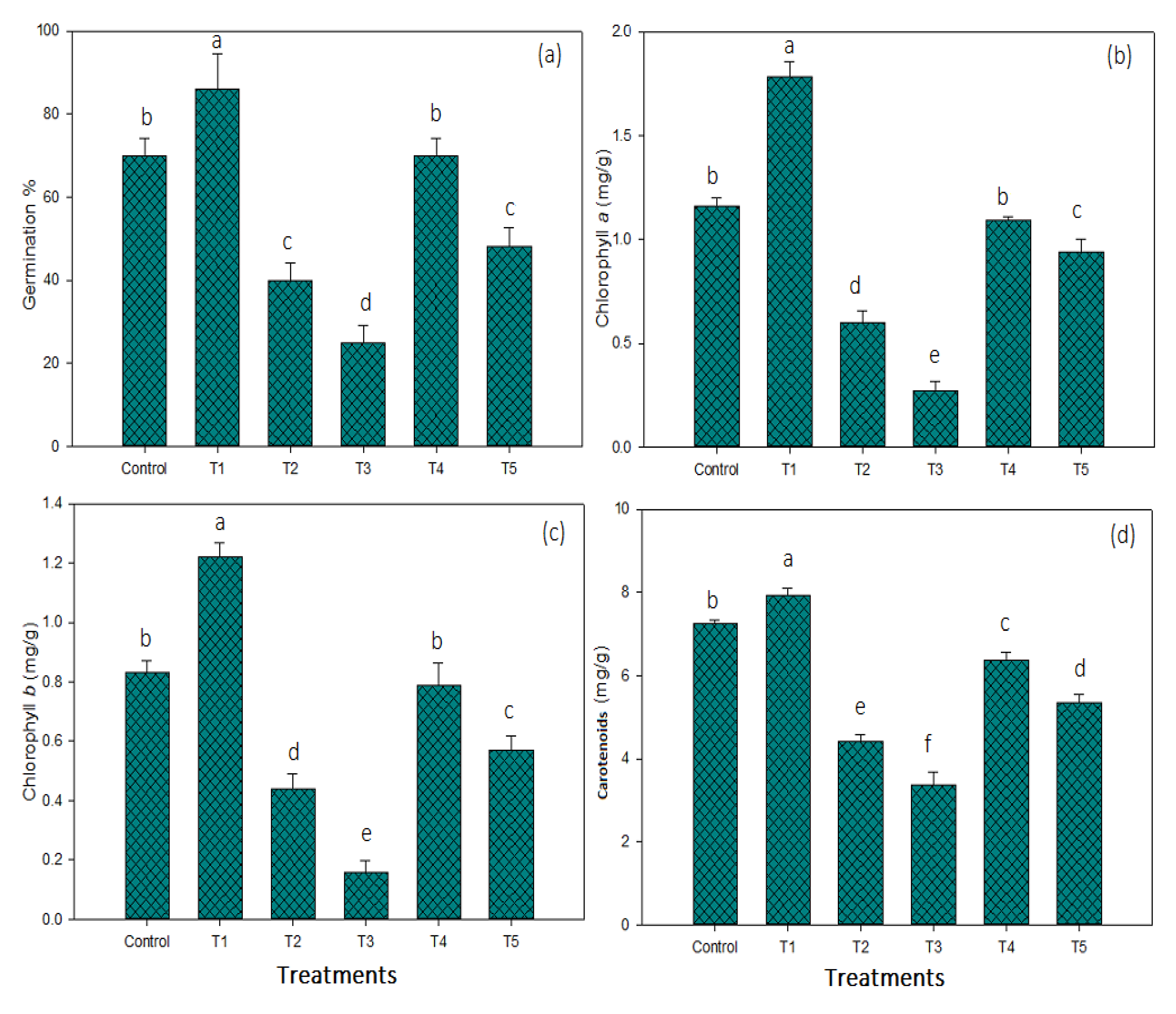
3.4. Chlorophyll Contents
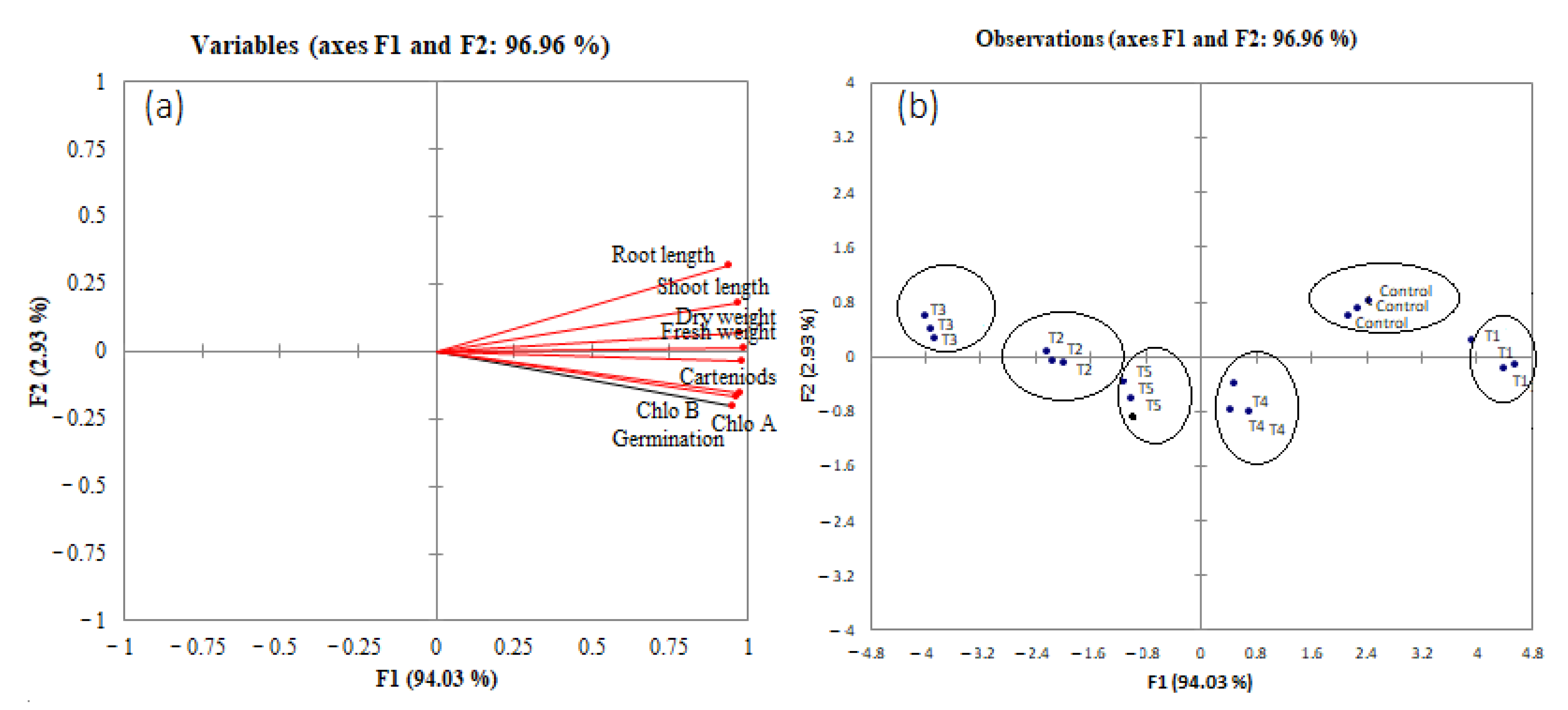
3.5. Principal Component and Pearson Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jasrotia, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kumar, S.; Singh, G.P. Scope and applications of nanotechnology for wheat production: A review of recent advances. Wheat Barley Res. 2018, 10, 1–14. [Google Scholar]
- Tsvetanov, T.; Qi, L.; Mukherjee, D.; Shah, F.; Bravo-Ureta, B. Climate change and land use in Southeastern US: Did the “dumb farmer” get it wrong? Clim. Chang. Econ. 2016, 7, 1650005. [Google Scholar] [CrossRef]
- Tiwari, P.; Indoliya, Y.; Singh, P.K.; Singh, P.C.; Chauhan, P.S.; Pande, V.; Chakrabarty, D. Role of dehydrin-FK506-binding protein complex in enhancing drought tolerance through the ABA-mediated signaling pathway. Environ. Exp. Bot. 2019, 158, 136–149. [Google Scholar] [CrossRef]
- Afridi, M.S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Chaudhary, H.J. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; A.l-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruinsma, J. (Ed.) World Agriculture: Towards 2015/2030: An FAO Perspective; Earthscan: London, UK, 2003. [Google Scholar]
- Panta, S.; Flowers, T.J.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases—A field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Ben-Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2021, 152, 104444. [Google Scholar] [CrossRef]
- Mejdoub-Trabelsi, B.; Touihri, S.; Ammar, N.; Riahi, A.; Daami-Remadi, M. Effect of chitosan for the control of potato diseases caused by Fusarium species. J. Phytopathol. 2020, 168, 18–27. [Google Scholar] [CrossRef]
- De-Oliveira, D.F.; de Sousa Lopes, L.; Gomes-Filho, E. Metabolic changes associated with differential salt tolerance in sorghum genotypes. Planta 2020, 252, 1–18. [Google Scholar] [CrossRef]
- Zainab, N.; Din, B.U.; Javed, M.T.; Afridi, M.S.; Mukhtar, T.; Kamran, M.A.; Chaudhary, H.J. Deciphering metal toxicity responses of flax (Linum usitatissimum L.) with exopolysaccharide and ACC-deaminase producing bacteria in industrially contaminated soils. Plant Physiol. Biochem. 2020, 152, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Amin, I.; Rasool, S.; Mir, M.A.; Wani, W.; Masoodi, K.Z.; Ahmad, P. Ion homeostasis for salinity tolerance in plants: A molecular approach. Physiol. Plant. 2021, 171, 578–594. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd_Allah, E.F. Impact of soil salinity on the plant-growth–promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-tolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Akhtar, M.J.; Mehmood, S.; Akhter, K.; Tahir, M.; Saeed, M.F.; Hussain, S. Combined application of compost and Bacillus sp. CIK-512 ameliorated the lead toxicity in radish by regulating the homeostasis of antioxidants and lead. Ecotoxicol. Environ. Saf. 2018, 148, 805–812. [Google Scholar] [CrossRef]
- Amna, S.S.; Xia, Y.; Kamran, M.A.; Javed, M.T.; Sultan, T.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of wheat inoculated with exopolysaccharide and ACC-deaminase producing Bacillus strains under induced salinity stress. Ecotoxicol. Environ. Saf. 2019, 183, 109466. [Google Scholar] [CrossRef] [PubMed]
- Amna, X.Y.; Farooq, M.A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Chaudhary, H.J. Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, V.K.; Tripathi, V.; Singh, P.P.; Singh, A.K. Plant growth-promoting rhizobacteria (PGPR): Perspective in agriculture under biotic and abiotic stress. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 333–342. [Google Scholar]
- Marzan, L.W.; Hossain, M.; Mina, S.A.; Akter, Y.; Chowdhury, A.M.A. Isolation and biochemical characterization of heavy-metal resistant bacteria from tannery effluent in Chittagong city, Bangladesh: Bioremediation viewpoint. Egypt. J. Aquat. Res. 2017, 43, 65–74. [Google Scholar] [CrossRef]
- Mejdoub-Trabelsi, B.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Effect of Fusarium species and temperature of storage on the susceptibility ranking of potato cultivars to tuber dry rot. Pest. Technol. 2012, 6, 41–46. [Google Scholar]
- Parmar, P.; Sindhu, S.S. Potassium solubilization by rhizosphere bacteria: Influence of nutritional and environmental conditions. J. Microbiol. Res. 2013, 3, 25–31. [Google Scholar]
- Gontia-Mishra, I.; Sapre, S.; Tiwari, S. Zinc solubilizing bacteria from the rhizosphere of rice as prospective modulator of zinc biofortification in rice. Rhizosphere 2017, 3, 185–190. [Google Scholar] [CrossRef]
- Tahir, M.; Khalid, U.; Khan, M.B.; Shahid, M.; Ahmad, I.; Akram, M.; Ahmad, N. Auxin and 1-Aminocyclopropane-1-carboxylate deaminase activity exhibiting rhizobacteria improved maize quality and productivity under drought conditions. Int. J. Agric. Biol. 2019, 21, 943–954. [Google Scholar]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, S.; Amna, A.; Mehmood, S.; Suhaib, M.; Sultan, T.; Munis, M.F.H. Interaction Of Acc Deaminase And Antioxidant Enzymes To Induce Drought Tolerance In Enterobacter Cloacae 2wc2 Inoculated Maize Genotypes. Pak. J. Bot. 2021, 53, 3. [Google Scholar] [CrossRef]
- Hjort, K.; Presti, I.; Elväng, M.F.; Sjöling, S. Bacterial chitinase with phytopathogen control capacity from suppressive soil revealed by functional metagenomics. Appl. Microbiol. Biotechnol. 2014, 98, 2819–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Bano, A. Role of plant growth promoting rhizobacteria and Ag-nano particle in the bioremediation of heavy metals and maize growth under municipal wastewater irrigation. Int. J. Phytoremediation 2016, 18, 211–221. [Google Scholar] [CrossRef]
- Mukhtar, T.; Smith, D.; Sultan, T.; Seleiman, M.F.; Alsadon, A.A.; Ali, S.; Saad, M.A. Mitigation of heat stress in Solanum lycopersicum L. by ACC-deaminase and exopolysaccharide producing Bacillus cereus: Effects on biochemical profiling. Sustainability 2020, 12, 2159. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; McREYNOLDS, L.A. Use of degenerate oligonucleotides for amplification of the nifH gene from the marine cyanobacterium Trichodesmium thiebautii. Appl. Environ. Microbiol. 1989, 55, 2522–2526. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.M.; Soares, H.M.; Soares, E.V. Promising bacterial genera for agricultural practices: An insight on plant growth-promoting properties and microbial safety aspects. Sci. Total Environ. 2019, 682, 779–799. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.Z.; Sandhya, V.; Grover, M.; Kishore, N.; Rao, L.V.; Venkateswarlu, B. Pseudomonas sp. strain AKM-P6 enhances tolerance of sorghum seedlings to elevated temperatures. Biol. Fertil. Soils 2009, 46, 45–55. [Google Scholar] [CrossRef]
- Din, B.U.; Rafique, M.; Javed, M.T.; Kamran, M.A.; Mehmood, S.; Khan, M.; Chaudhary, H.J. Assisted phytoremediation of chromium spiked soils by Sesbania Sesban in association with Bacillus xiamenensis PM14: A biochemical analysis. Plant Physiol. Biochem. 2020, 146, 249–258. [Google Scholar] [CrossRef]
- Muneer, M.A.; Wang, P.; Zhang, J.; Li, Y.; Munir, M.Z.; Ji, B. Formation of common mycorrhizal networks significantly affect plant biomass and soil properties of the neighboring plants under various nitrogen levels. Microorganisms 2020, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Muneer, M.A.; Wang, P.; Lin, C.; Ji, B. Potential role of common mycorrhizal networks in improving plant growth and soil physicochemical properties under varying nitrogen levels in a grassland ecosystem. Glob. Ecol. Conserv. 2020, 24, e01352. [Google Scholar] [CrossRef]
- Natasha, S.M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Farooq, A.B.U.; Rabbani, F.; Khalid, S.; Dumat, C. Risk assessment and biophysiochemical responses of spinach to foliar application of lead oxide nanoparticles: A multivariate analysis. Chemosphere 2020, 245, 125605. [Google Scholar]
- Duan, J.; Müller, K.M.; Charles, T.C.; Vesely, S.; Glick, B.R. 1-aminocyclopropane-1-carboxylate (ACC) deaminase genes in rhizobia from southern Saskatchewan. Microb. Ecol. 2009, 57, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of exogenous salicylic acid and nitric oxide on growth, photosynthesis, and ascorbate-glutathione cycle in salt stressed Vigna angularis. Biomolecules 2020, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Ali, J.; Ali, F.; Ahmad, I.; Rafique, M.; Munis, M.F.H.; Hassan, S.W.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of Sesbania sesban seedlings with Bacillus anthracis PM21 under heavy metals stress: An in vitro study. Ecotoxicol. Environ. Saf. 2021, 208, 111769. [Google Scholar] [CrossRef]
- Yoo, S.J.; Weon, H.Y.; Song, J.; Sang, M.K. Induced tolerance to salinity stress by halotolerant bacteria Bacillus aryabhattai H19-1 and B. mesonae H20-5 in tomato plants. J. Microbiol. Biotechnol. 2019, 29, 1124–1136. [Google Scholar] [CrossRef]
- Alam, P.; Albalawi, T.H.; Altalayan, F.H.; Bakht, M.A.; Ahanger, M.A.; Raja, V.; Ahmad, P. 24-Epibrassinolide (EBR) confers tolerance against NaCl stress in soybean plants by up-regulating antioxidant system, ascorbate-glutathione cycle, and glyoxalase system. Biomolecules 2019, 9, 640. [Google Scholar] [CrossRef] [Green Version]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physiol. Plant. 2020, 168, 256–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) supplementation alleviates NaCl toxicity in mung bean [Vigna radiata (L.) Wilczek] through the modifications of physio-biochemical attributes and key antioxidant enzymes. J. Plant Growth Reg. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Bibi, F.; Yasir, M.; Al-Sofyani, A.; Naseer, M.I.; Azhar, E.I. Antimicrobial activity of bacteria from marine sponge Suberea mollis and bioactive metabolites of Vibrio sp. EA348. Saudi J. Biol. Sci. 2020, 27, 1139–1147. [Google Scholar] [CrossRef]
- Haidar, R.; Roudet, J.; Bonnard, O.; Dufour, M.C.; Corio-Costet, M.F.; Fert, M.; Fermaud, M. Screening and modes of action of antagonistic bacteria to control the fungal pathogen Phaeomoniella chlamydospora involved in grapevine trunk diseases. Microbiol. Res. 2017, 192, 172–184. [Google Scholar] [CrossRef]
- Cray, J.A.; Houghton, J.D.; Cooke, L.R.; Hallsworth, J.E. A simple inhibition coefficient for quantifying potency of biocontrol agents against plant-pathogenic fungi. Biol. Control. 2015, 81, 93–100. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | Primer Sequence (5′-3′) | PCR Product Size Expected/Detected (pb) | References |
|---|---|---|---|---|
| 16S rRNA | 27F 1492R | AGAGTTTGATC AC TGGCTCAG CGG CTTACCTTGTTACGACTT | 1500/yes | [29] |
| ituC | ITUC-F1 ITUC-R1 | CCCCCTCGG TCAAGTGAATA TTGGTTAAGCCCTGATGCTC | 594/yes | [18] |
| nifH | nif h-F1 nif h-R1 | TATGATCCAAAAGCAGA ATAGCCATCATTTCACC | 360/yes | [30] |
| acds | acds-F1 acds-R1 | GGCAAGGTCGACATCTATGC GGCTTGCCATTCAGCTATG | 560/yes | [31] |
| Bacterial Traits | Results |
|---|---|
| Morphological and colony features | Medium size colony, wrinkled growth, off-white, smooth colonies at 32 °C after 1 d on LB agar. |
| Bacterial cell traits | Gram-positive, under microscope, showed scattered arrangement of short rod-shaped cells. |
| Biochemical properties | Positive for atmospheric nitrogen-fixing ability, potassium solubilization, zinc solubilization, IAA production, siderophore, ammonia, ACC deaminase, pectinase, protease, amylase, catalase, cellulase, chitinase, and exopolysaccharides production. |
| Treatments | Germination % | Root Length (cm) | Shoot Length (cm) | Fresh Weight (g) | Dry Weight (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Un-inoculated | Inoculated | Un-inoculated | Inoculated | Un-inoculated | Inoculated | Un-inoculated | Inoculated | Un-inoculated | Inoculated | |
| Control | 68.3 ± 2.3 b | 88.3 ± 6.2 a | 17.7 ± 0.55 b | 20.7 ± 0.71 a | 13.13 ± 0.27 b | 14.74 ± 0.05 a | 0.39 ± 0.02 b | 0.50 ± 0.06 a | 0.16 ± 0.2 b | 0.20 ± 0 a |
| NaCl 1M | 40. ± 4 c | 65 ± 4.8 b | 3. ± 0.46 d | 5.96 ± 0.57 c | 1.74 ± 0.07 e | 7.66 ± 0.69 c | 0.15 ± 0.01 d | 0.28 ± 0 c | 0.07 ± 0 d | 0.12 ± 0.08 c |
| NaCl 2 M | 23.3 ± 2.3 d | 48.5 ± 4.7 c | 0.5 ± 0.08 e | 2.73 ± 0.16 d | 1.08 ± 0.29 f | 4.46 ± 0.85 d | 0.04 ± 0 e | 0.22 ± 0.01 c | 0.01 ± 0 e | 0.09 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehmood, S.; Khan, A.A.; Shi, F.; Tahir, M.; Sultan, T.; Munis, M.F.H.; Kaushik, P.; Alyemeni, M.N.; Chaudhary, H.J. Alleviation of Salt Stress in Wheat Seedlings via Multifunctional Bacillus aryabhattai PM34: An In-Vitro Study. Sustainability 2021, 13, 8030. https://0-doi-org.brum.beds.ac.uk/10.3390/su13148030
Mehmood S, Khan AA, Shi F, Tahir M, Sultan T, Munis MFH, Kaushik P, Alyemeni MN, Chaudhary HJ. Alleviation of Salt Stress in Wheat Seedlings via Multifunctional Bacillus aryabhattai PM34: An In-Vitro Study. Sustainability. 2021; 13(14):8030. https://0-doi-org.brum.beds.ac.uk/10.3390/su13148030
Chicago/Turabian StyleMehmood, Shehzad, Amir Abdullah Khan, Fuchen Shi, Muhammad Tahir, Tariq Sultan, Muhammad Farooq Hussain Munis, Prashant Kaushik, Mohammed Nasser Alyemeni, and Hassan Javed Chaudhary. 2021. "Alleviation of Salt Stress in Wheat Seedlings via Multifunctional Bacillus aryabhattai PM34: An In-Vitro Study" Sustainability 13, no. 14: 8030. https://0-doi-org.brum.beds.ac.uk/10.3390/su13148030