Effect of Phytohormones Supplementation under Nitrogen Depletion on Biomass and Lipid Production of Nannochloropsis oceanica for Integrated Application in Nutrition and Biodiesel


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Growth
2.2. Experimental Design
2.3. Growth Determination
2.4. Chlorophyll Content, Fatty Acid Profile, and FAMEs Characteristics
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effect on Biomass Production
3.2. Effect on the Lipid Accumulation
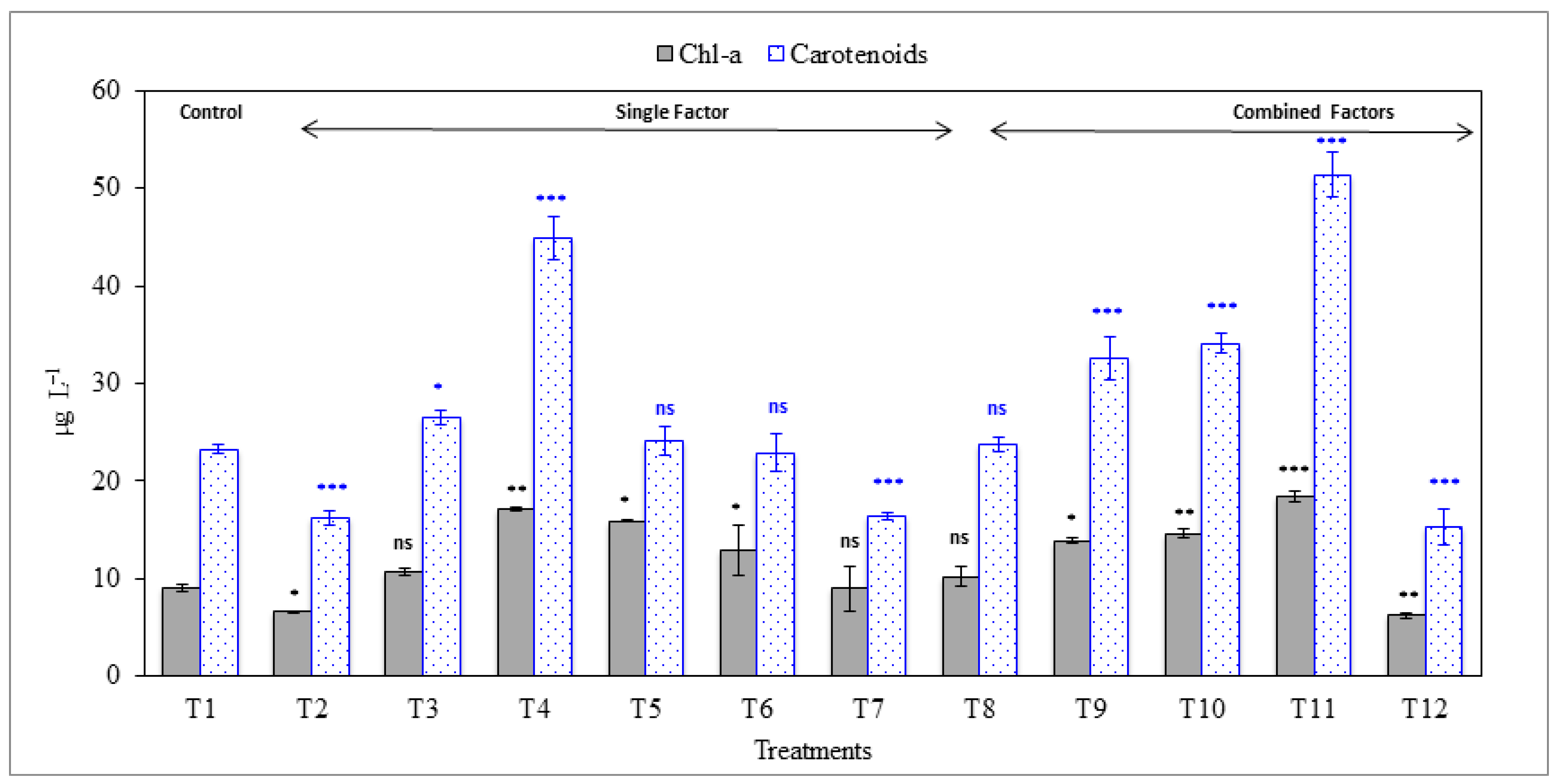
3.3. Effect on Pigments
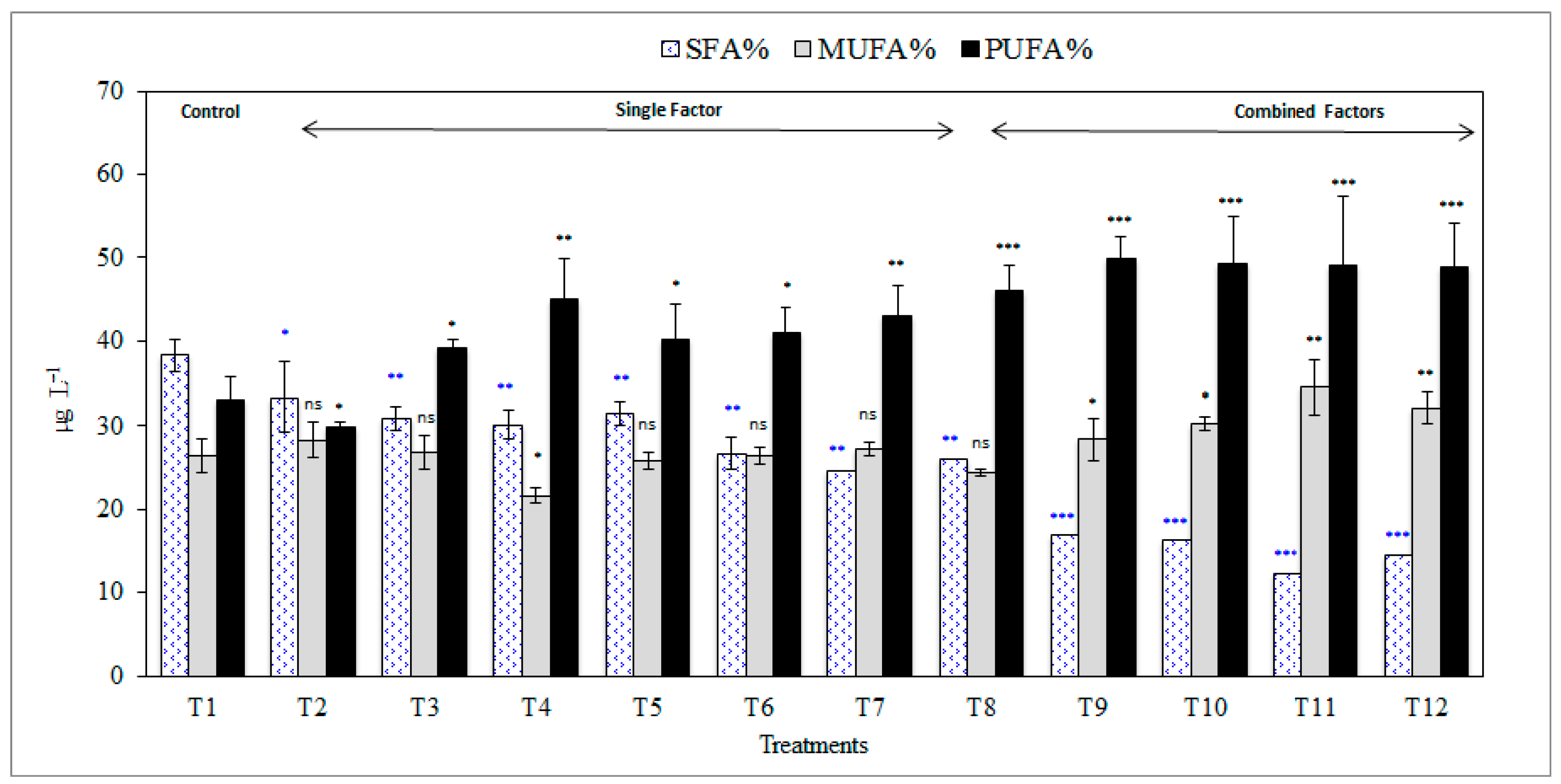
3.4. Effects on Fatty Acids Profile and Biodiesel Properties.
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Almutairi, A.W. Effects of nitrogen and phosphorus limitations on fatty acid methyl esters and fuel properties of Dunaliella salina. Environ. Sci. Pollut. Res. 2020, 27, 32296–32303. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, A.W. Improvement of Chemical Composition of Tisochrysis lutea Grown Mixotrophically under Nitrogen Depletion towards Biodiesel Production. Molecules 2020, 25, 4609. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, A.W.; El-Sayed, A.E.-K.B.; Reda, M.M. Combined effect of salinity and pH on lipid content and fatty acid composition of Tisochrysis lutea. Saudi J. Biol. Sci. 2020, 27, 3553–3558. [Google Scholar] [CrossRef] [PubMed]
- Huete-Ortega, M.; Okurowska, K.; Kapoore, R.V.; Johnson, M.P.; Gilmour, D.J.; Vaidyanathan, S. Effect of ammonium and high light intensity on the accumulation of lipids in Nannochloropsis oceanica (CCAP 849/10) and Phaeodactylum tricornutum (CCAP 1055/1). Biotechnol. Biofuels 2018, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Abomohra, A.E.-F.; Jin, W.; Tu, R.; Han, S.-F.; Eid, M.; Eladel, H. Microalgal biomass production as a sustainable feedstock for biodiesel: Current status and perspectives. Renew. Sustain. Energy Rev. 2016, 64, 596–606. [Google Scholar] [CrossRef]
- Abomohra, A.; Zheng, X.; Wang, Q.; Huang, J.; Ebaid, R. Enhancement of biodiesel yield and characteristics through in-situ solvo-thermal co-transesterification of wet microalgae with spent coffee grounds. Bioresour. Technol. 2020, 31, 124640. [Google Scholar] [CrossRef]
- Udayan, A.; Sabapathy, H.; Arumugam, M. Stress hormones mediated lipid accumulation and modulation of specific fatty acids in Nannochloropsis oceanica CASA CC201. Bioresour. Technol. 2020, 310, 123437. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Voß, U.; Bishopp, A.; Farcot, E.; Bennett, M.J. Modelling hormonal response and development. Trends Plant Sci. 2014, 19, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Kumari, S.; Guldhe, A.; Misra, R.; Rawat, I.; Bux, F. Trends and novel strategies for enhancing lipid accumulation and quality in microalgae. Renew. Sustain. Energy Rev. 2016, 55, 1–16. [Google Scholar] [CrossRef]
- Gomaa, M.; Refaat, M.; Salim, T.; El-Khair, A.; El-Sayed, B.; Bekhit, M. Identification of Green Alga Chlorella vulgaris Isolated from Freshwater and Improvement Biodiesel Productivity via UV Irradiation. Microbiol. Biotechnol. Lett. 2019, 47, 381–389. [Google Scholar] [CrossRef]
- Xin, L.; Hu, H.Y.; Ke, G.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yuan, H.; Li, B.; Yang, J. Significance evaluation of the effects of environmental factors on the lipid accumulation of Chlorella minutissima UTEX 2341 under low-nutrition heterotrophic condition. Bioresour. Technol. 2014, 152, 177–184. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.B. Some Physiological Studies on Green Algae; Plant Physiology Department, Faculty of Agriculture Cairo University: Cairo, Egypt, 1999. [Google Scholar]
- El-Sayed, A.; El-Fouly, M.; El-Sayed, A. Utilization efficiency of elevated nitrogen, phosphorous and potassium concentrations by the green alga Scenedesmus sp. In Proceedings of the 17th International Symposium of CIEC “Plant Nutrient Management under Stress Conditions”, Cairo, Egypt, 24–27 November 2008. [Google Scholar]
- Ji, F.; Hao, R.; Liu, Y.; Li, G.; Zhou, Y.; Dong, R. Isolation of a novel microalgae strain Desmodesmus sp. and optimization of environmental factors for its biomass production. Bioresour. Technol. 2013, 148, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process. Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Gao, B.; Liu, J.; Zhang, C.; Van de Waal, D.B. Biological stoichiometry of oleaginous microalgal lipid synthesis: The role of N:P supply ratios and growth rate on microalgal elemental and biochemical composition. Algal Res. 2018, 32, 353–361. [Google Scholar] [CrossRef]
- Roessler, P.G. Purification and Characterization of Acetyl-CoA Carboxylase from the Diatom Cyclotella cryptica. Plant Physiol. 1990, 92, 73. [Google Scholar] [CrossRef] [Green Version]
- Khozin-Goldberg, I.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef]
- Jeyakumar, B.; Asha, D.; Varalakshmi, P.; Kathiresan, S. Nitrogen repletion favors cellular metabolism and improves eicosapentaenoic acid production in the marine microalga Isochrysis sp. CASA CC 101. Algal Res. 2020, 47, 101877. [Google Scholar] [CrossRef]
- Fawley, K.P.; Fawley, M.W. Observations on the Diversity and Ecology of Freshwater Nannochloropsis (Eustigmatophyceae), with Descriptions of New Taxa. Protist 2007, 158, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Boussiba, S.; Vonshak, A.; Cohen, Z.; Avissar, Y.; Richmond, A. Lipid and biomass production by the halotolerant microalga Nannochloropsis salina. Biomass 1987, 12, 37–47. [Google Scholar] [CrossRef]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.-W.; Abomohra, A.E.-F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Wagner, M.; El-Sheekh, M.; Hanelt, D. Lipid and total fatty acid productivity in photoautotrophic fresh water microalgae: Screening studies towards biodiesel production. J. Appl. Phycol. 2012, 25, 931–936. [Google Scholar] [CrossRef]
- Roy, S.; Llewellyn, C.A.; Egeland, E.S.; Johnsen, G. (Eds.) Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Jin, W.; El-Sheekh, M. Enhancement of lipid extraction for improved biodiesel recovery from the biodiesel promising microalga Scenedesmus obliquus. Energy Convers. Manag. 2016, 108, 23–29. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties, and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; El-Naggar, A.H.; Baeshen, A.A. Potential of macroalgae for biodiesel production: Screening and evaluation studies. J. Biosci. Bioeng. 2018, 125, 231–237. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Chivkunova, O.B.; Gorelova, O.A.; Reshetnikova, I.V.; Solovchenko, A.E.; Khozin-Goldberg, I.; Cohen, Z. Effect of Nitrogen Starvation on Optical Properties, Pigments, and Arachidonic Acid Content of the Unicellular Green Alga Parietochloris Incisa (Trebouxiophyceae, Chlorophyta)1. J. Phycol. 2007, 43, 833–843. [Google Scholar] [CrossRef]
- Ozioko, F.; Chiejina, N.; Ogbonna, J. Effect of some phytohormones on growth characteristics of Chlorella sorokiniana IAM-C212 under photoautotrophic conditions. Afr. J. Biotechnol. 2015, 14, 2367–2376. [Google Scholar] [CrossRef]
- Magierek, E.; Krzemińska, I.; Tys, J. Stimulatory effect of indole-3-acetic acid and continuous illumination on the growth of Parachlorella kessleri. Int. Agrophys. 2017, 31, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Moussa, I.; Chtourou, H.; Karray, F.; Sayadi, S.; Dhouib, A. Nitrogen or phosphorus repletion strategies for enhancing lipid or carotenoid production from Tetraselmis marina. Bioresour. Technol. 2017, 238, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Almarashi, J.Q.M.; El-Zohary, S.E.; Ellabban, M.A.; Abomohra, A.E.-F. Enhancement of lipid production and energy recovery from the green microalga Chlorella vulgaris by inoculum pretreatment with low-dose cold atmospheric pressure plasma (CAPP). Energy Convers. Manag. 2020, 204, 112314. [Google Scholar] [CrossRef]
- El-Sheekh, M.; Abomohra Ael, F.; Hanelt, D. Optimization of biomass and fatty acid productivity of Scenedesmus obliquus as a promising microalga for biodiesel production. World J. Microbiol. Biotechnol. 2013, 29, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Esakkimuthu, S.; Krishnamurthy, V.; Wang, S.; Hu, X.; Swaminathan, K.; Abomohra, A.E.-F. Application of p-coumaric acid for extraordinary lipid production in Tetradesmus obliquus: A sustainable approach towards enhanced biodiesel production. Renew. Energy 2020, 157, 368–376. [Google Scholar] [CrossRef]
- Gonzalez, L.E.; Bashan, Y. Increased growth of the microalga Chlorella vulgaris when coimmobilized and cocultured in alginate beads with the plant-growth-promoting bacterium Azospirillum brasilense. Appl. Environ. Microbiol. 2000, 66, 1527–1531. [Google Scholar] [CrossRef] [Green Version]
- Dao, G.-H.; Wu, G.-X.; Wang, X.-X.; Zhang, T.-Y.; Zhan, X.-M.; Hu, H.-Y. Enhanced microalgae growth through stimulated secretion of indole acetic acid by symbiotic bacteria. Algal Res. 2018, 33, 345–351. [Google Scholar] [CrossRef]
- Mandal, M.K.; Chanu, N.K.; Chaurasia, N. Exogenous addition of indole acetic acid and kinetin under nitrogen-limited medium enhances lipid yield and expression of glycerol-3-phosphate acyltransferase & diacylglycerol acyltransferase genes in indigenous microalgae: A potential approach for biodiesel production. Bioresour. Technol. 2020, 297, 122439. [Google Scholar] [CrossRef]
- ASTM D6751-08. Standard Specification for Biodiesel Fuel Blend Stock (B100) for Middle Distillate Fuels; ASTM International: West Conshohocken, PA, USA, 2008. [Google Scholar]
- EN14214. Automotive Fuels D Fatty Acid Methylesters (FAME) for Diesel Engines. Requirements and Test Methods; European Community Stand: London, UK, 2008. [Google Scholar]
- Abomohra, A.E.-F.; Elsayed, M.; Esakkimuthu, S.; El-Sheekh, M.; Hanelt, D. Potential of fat, oil and grease (FOG) for biodiesel production: A critical review on the recent progress and future perspectives. Prog. Energy Combust. Sci. 2020, 81, 100868. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Butyric Acid (C4:0) | 3.09 ± 0.09 | 3.63 ± 0.11 | 3.01 ± 0.19 | 2.61 ± 0.26 | 4.46 ± 0.31 | 3.14 ± 0.27 | 3.66 ± 0.45 | 3.08 ± 0.31 | 2.13 ± 0.24 | 3.01 ± 0.40 | 2.04 ± 0.24 | 3.19 ± 0.20 |
| Lauric Acid (C12:0) | 7.19 ± 0.52 | 5.46 ± 0.29 | 6.19 ± 0.48 | 5.43 ± 0.55 | 4.98 ± 0.68 | 5.87 ± 0.67 | 6.12 ± 0.86 | 4.92 ± 0.65 | 3.18 ± 0.22 | 4.14 ± 0.43 | 3.87 ± 0.13 | 2.58 ± 0.42 |
| Myristic (14:0) | 2.48 ± 0.40 | 2.44 ± 0.16 | 2.19 ± 0.20 | 3.18 ± 0.32 | 3.68 ± 0.32 | 3.07 ± 0.07 | 2.01 ± 0.35 | 2.94 ± 0.62 | 2.41 ± 0.37 | 2.04 ± 0.21 | 2.00 ± 0.38 | 3.45 ± 0.45 |
| Palmitic (16:0) | 12.08 ± 1.02 | 10.28 ± 0.01 | 9.13 ± 0.68 | 8.02 ± 0.20 | 8.61 ± 0.70 | 5.42 ± 0.64 | 5.14 ± 0.98 | 6.21 ± 0.93 | 4.19 ± 0.33 | 3.15 ± 0.36 | 1.18 ± 0.19 | 2.07 ± 0.26 |
| Stearic (18:0) | 5.19 ± 1.04 | 4.29 ± 0.64 | 2.45 ± 0.44 | 3.82 ± 0.41 | 2.52 ± 0.47 | 3.58 ± 0.53 | 2.09 ± 0.22 | 2.66 ± 0.01 | 0.95 ± 0.19 | 0.84 ± 0.14 | 0.87 ± 0.16 | 1.04 ± 0.15 |
| Arachidilic (C20:0) | 8.29 ± 0.96 | 7.28 ± 0.73 | 7.91 ± 1.02 | 6.97 ± 1.09 | 7.21 ± 0.93 | 6.58 ± 0.33 | 6.61 ± 0.49 | 6.27 ± 0.98 | 3.98 ± 0.79 | 3.14 ± 0.40 | 2.27 ± 0.38 | 2.18 ± 0.35 |
| Myristoleic (14:1) | 3.89 ± 0.41 | 2.89 ± 0.50 | 2.45 ± 0.31 | 2.51 ± 0.33 | 4.08 ± 0.10 | 4.28 ± 0.59 | 5.26 ± 0.44 | 3.17 ± 0.38 | 4.97 ± 0.89 | 4.96 ± 0.26 | 3.97 ± 0.58 | 4.07 ± 0.21 |
| Palmitoleic (16:1) | 6.09 ± 0.98 | 6.13 ± 0.77 | 8.08 ± 0.95 | 6.42 ± 0.94 | 7.19 ± 1.11 | 7.11 ± 1.16 | 6.27 ± 0.78 | 4.63 ± 0.37 | 7.16 ± 0.75 | 8.74 ± 1.12 | 8.56 ± 0.98 | 8.22 ± 0.83 |
| Oleic (18:1) | 15.39 ± 1.84 | 19.31 ± 1.74 | 16.33 ± 2.06 | 12.65 ± 0.57 | 14.55 ± 0.75 | 15.09 ± 0.93 | 15.63 ± 0.89 | 16.52 ± 0.75 | 16.21 ± 0.51 | 16.54 ± 1.59 | 21.00 ± 0.97 | 19.81 ± 1.26 |
| Linoleic (18:2; ω-6) | 12.56 ± 1.13 | 16.08 ± 1.04 | 16.07 ± 0.22 | 24.09 ± 0.34 | 16.27 ± 0.60 | 15.22 ± 0.70 | 16.24 ± 0.75 | 16.01 ± 1.26 | 20.24 ± 0.99 | 19.65 ± 0.67 | 20.81 ± 0.89 | 19.88 ± 1.43 |
| α-Linolenic (18:3; ω-3) | 4.36 ± 0.90 | 4.62 ± 1.11 | 5.95 ± 0.95 | 6.28 ± 0.93 | 8.41 ± 1.26 | 6.46 ± 1.47 | 7.14 ± 0.90 | 9.42 ± 0.65 | 8.19 ± 0.96 | 8.43 ± 0.40 | 6.91 ± 0.91 | 8.09 ± 0.31 |
| γ-Linolenic (18:3; ω-6) | 5.28 ± 0.64 | 6.15 ± 1.06 | 5.81 ± 0.01 | 5.81 ± 1.82 | 6.19 ± 0.77 | 7.99 ± 1.08 | 8.07 ± 0.01 | 10.37 ± 1.20 | 9.23 ± 0.69 | 10.16 ± 1.74 | 9.16 ± 1.19 | 8.34 ± 1.02 |
| Arachidonic (20:4) | 7.19 ± 0.84 | 6.12 ± 0.40 | 7.63 ± 0.69 | 5.92 ± 0.75 | 6.28 ± 1.05 | 8.41 ± 0.64 | 9.48 ± 0.66 | 6.23 ± 0.74 | 9.15 ± 0.85 | 8.24 ± 1.76 | 9.17 ± 0.77 | 9.08 ± 0.47 |
| Eicosapentaenoic (20:5; ω-3) | 3.66 ± 0.67 | 2.82 ± 0.30 | 3.65 ± 0.79 | 2.89 ± 0.52 | 3.08 ± 0.61 | 2.98 ± 0.33 | 2.18 ± 0.67 | 4.08 ± 0.43 | 3.17 ± 0.54 | 2.91 ± 0.46 | 3.08 ± 0.27 | 3.87 ± 0.23 |
| Docosahexanoic (22:6; ω-3) | 2.49 ± 0.32 | 2.03 ± 0.27 | 2.94 ± 0.11 | 2.65 ± 0.57 | 2.44 ± 0.49 | 4.89 ± 0.46 | 4.91 ± 0.62 | 3.18 ± 0.32 | 4.62 ± 0.50 | 3.87 ± 0.17 | 4.89 ± 0.25 | 3.98 ± 0.57 |
| Properties | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 | US | EN |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ADU | 1.41 | 1.44 | 1.61 | 1.60 | 1.57 | 1.78 | 1.84 | 1.80 | 2.01 | 1.96 | 2.05 | 2.01 | - | - |
| KV | 4.31 | 4.30 | 4.19 | 4.20 | 4.21 | 4.08 | 4.05 | 4.07 | 3.94 | 3.97 | 3.91 | 3.94 | 1.9–6.0 | 3.5–5.0 |
| SG | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.88 | 0.85–0.9 | - |
| CP | 1.11 | 0.82 | −1.47 | −1.38 | −1.02 | −3.80 | −4.52 | −4.06 | −6.88 | −6.19 | −7.36 | −6.81 | - | - |
| CN | 53.45 | 53.30 | 52.16 | 52.20 | 52.39 | 51.00 | 50.64 | 50.87 | 49.46 | 49.80 | 49.22 | 49.49 | Min. 47 | 51–120 |
| IV | 117.86 | 119.48 | 132.22 | 131.75 | 129.71 | 145.21 | 149.22 | 146.66 | 162.39 | 158.51 | 165.01 | 161.98 | - | Max. 120 |
| HHV | 41.02 | 41.06 | 41.36 | 41.35 | 41.30 | 41.67 | 41.76 | 41.70 | 42.08 | 41.98 | 42.14 | 42.07 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touliabah, H.E.-S.; Almutairi, A.W. Effect of Phytohormones Supplementation under Nitrogen Depletion on Biomass and Lipid Production of Nannochloropsis oceanica for Integrated Application in Nutrition and Biodiesel. Sustainability 2021, 13, 592. https://0-doi-org.brum.beds.ac.uk/10.3390/su13020592
Touliabah HE-S, Almutairi AW. Effect of Phytohormones Supplementation under Nitrogen Depletion on Biomass and Lipid Production of Nannochloropsis oceanica for Integrated Application in Nutrition and Biodiesel. Sustainability. 2021; 13(2):592. https://0-doi-org.brum.beds.ac.uk/10.3390/su13020592
Chicago/Turabian StyleTouliabah, Hussein El-Sayed, and Adel W. Almutairi. 2021. "Effect of Phytohormones Supplementation under Nitrogen Depletion on Biomass and Lipid Production of Nannochloropsis oceanica for Integrated Application in Nutrition and Biodiesel" Sustainability 13, no. 2: 592. https://0-doi-org.brum.beds.ac.uk/10.3390/su13020592