1. Introduction
Studies on plant growth-promoting rhizobacteria (PGPR) have attracted much attention in the last decade. Application of PGPR with promising traits was considered as an environmentally friendly approach to sustain plant growth, which also reduced the requirement of chemical fertilization [
1]. Among the beneficial bacteria, rhizobia that are able to form a symbiotic relationship with their host legumes are well studied [
2,
3]. To establish proper nodule formation, bacterial infection and nodule organogenesis need to be coordinated in plant tissue through signaling cascades [
4]. It is a considerable challenge to understand the mechanisms by which legumes recognize and discriminate rhizobia from pathogens, as well as between distinct rhizobia species and strains that differ in their symbiotic performance [
5]. Besides, a variety of PGPR have been demonstrated to enhance the legume–rhizobia symbiosis through improving the nodulation and nitrogen fixation [
6].
Soybean (
Glycine max L.) is an agronomically and economically relevant leguminous plant cultivated worldwide. It also showed potential to increase soil fertility when used as green manure or cover crop [
7,
8,
9]. Studies on bacterial community inside soybean nodules may help to explore effective participants involving in symbiotic nitrogen fixation and plant growth. It was reported that rhizobial subcommunities comprised mainly
Ensifer and
Bradyrhizobium, while the non-rhizobial groups were dominated by
Proteobacteria and
Firmicutes after screening nodule endophytes from fifty soybean fields [
10]. Through the microbiome’s analyses of nodules from nine soybean cultivars, Sharaf et al. [
11] indicated that rhizobia were mainly composed of the family
Bradyrhizobiaceae. Besides, the non-rhizobia such as
Pseudomonadaceae and
Enterobacteriaceae also dominated in soybean nodules. Within
Bradyrhizobiaceae, three genera, namely,
Bradyrhizobium,
Nitrobacter, and
Tardiphaga, were consistently detected in almost 193 root nodules of nine soybean plants [
12]. The OTU analysis further demonstrated that
Bradyrhizobium dominated during the entire stage of soybean growth, while the ratio of non-rhizobial bacteria showed an increasing trend as the soybean growth progressed [
13]. From a meta-analysis of studies (from 1987 to 2018) conducted on soybean, Zeffa et al. [
14] summarized that co-inoculation with
Bradyrhizobium and PGPR resulted in a significant increase in nodule number (11.40%), nodule biomass (6.47%), root biomass (12.84%), and shoot biomass (6.53%). Among these PGPR, members belonging to genera
Azospirillum,
Bacillus, and
Pseudomonas were more effective than
Serratia in the improvement of nodule formation and soybean growth.
In our previous study, soybean intercropping was demonstrated to ameliorate tea-cultivated environments through changing bacterial communities as well as edaphic properties [
9]. However, the roots of soybeans failed to form nodules during the experimental period, which was probably because of the strongly acidic nature of the tea plantation soils. As different species and strains differ in their symbiotic performance, it is crucial to explore effective rhizobia and non-rhizobia participating in the nodulation and growth promotion of soybean, especially in strongly acidic soils. In the present study, attempts were made to trap acidotolerant nodule-associated bacteria from soybean grown in a slightly acidic tea plantation. Effective rhizobial isolates were screened by nodulation test, while potentially plant growth-promoting and plant constituent-hydrolyzing traits were determined in non-rhizobial isolates. Effects of co-inoculation with beneficial rhizobial and non-rhizobial isolates on symbiotic nitrogen fixation and soybean growth were evaluated in sterile water agar (flask experiment) and non-sterile strongly acidic soils (pot experiment). This was used to test the hypothesis that acidotolerant nodule-inhabitants participated in the establishment of effective association with soybean, which further enhanced symbiotic nitrogen fixation and plant growth in the strongly acidic soils.
2. Materials and Methods
2.1. Isolation and Identification of Bacteria from Root Nodules
The green manure soybean (
Glycine max L.) cultivar Tainan no. 7 was cultivated in a slightly acidic (pH 6.5) tea plantation for two months. Root nodules were collected and surface sterilized with 2% of sodium hypochlorite for 1 min followed by 70% of ethanol for 5 min. Another rinse with sterile water was repeated five times and the final rinse water was used to assess the sterilization process. Nodules were crushed and effluents were collected, serially diluted, and plated on acidic (pH 5) yeast extract mannitol agar (YEMA) [
15]. Colonies that appeared within seven days of cultivation were picked and transferred several times to assure purity.
Genomic DNA was isolated from the three-day fresh cultures using the UltraClean Microbial Genomic DNA Isolation Kit (MO BIO Laboratories, Carlsbad, CA, USA) following the manufacturer’s recommendations. 16S rDNA was amplified by PCR with bacterial universal primers, purified, and sequenced as described in [
16], and bacterial identification was then performed by comparing the 16S rDNA sequences of isolates and their most closely related type strains using the EzBioCloud 16S-based ID function [
17]. The 16S rDNA sequences obtained in this study were deposited in the NCBI GenBank database under Accession Numbers MZ798477–MZ798484.
2.2. Characterization of nifH and nodD Genes in Rhizobial Isolates
The
nifH gene, which encodes the dinitrogenase reductase, was amplified by PCR with primer pair nifHF and nifHI [
18]. The amplification cycles included an initial denaturation for 3 min at 94 °C followed by 30 cycles of 1 min at 94 °C, 1 min at 57 °C, and 2 min at 72 °C, with a final extension of 3 min at 72 °C. The
nodD gene, which encodes the transcriptional regulator LysR family, was amplified by PCR with primer pair nodD7f and aboxp2 [
19]. The amplification cycles included an initial denaturation for 30 s at 95 °C followed by 40 cycles of 30 s at 96 °C, 1 min at 53 °C, and 30 s at 72 °C, with a final extension of 5 min at 72 °C. Both the
nifH and
nodD amplicons were purified using DNA Clean & Concentrator Kit (ZYMO RESEARCH CORP., USA). Cycle sequencing and determination of the nucleotide sequence of the amplicons were performed at Genomics BioSci&Tech Ltd., Taiwan. The NCBI GenBank database was used to identify the closely related sequences with the amplicons using the BLASTn program. Concatenated sequences from 16S rDNA,
nifH, and
nodD genes were aligned using ClustalX program version 2 [
20] and distances and clustering with the neighbor-joining method were performed using the software package MEGA version 11 [
21]. The
nifH and
nodD gene sequences obtained in this study were deposited in the NCBI GenBank database under Accession Numbers MZ803186–MZ803191.
2.3. Nodulation Test of Rhizobial Isolates
Isolates that were classified as rhizobia were tested for their abilities to induce nodulation of soybean in a growth chamber (8 h day/16 h night cycles) at 25 °C for 45 days. Isolates were grown on YEM for three days, and cells were then harvested by centrifugation, washed twice, and resuspended in YEM to obtain bacterial suspension. Seeds were surface sterilized with 30% of hydrogen peroxide for 5 min, 70% of alcohol for 5 min, and rinsed five times with sterile water. After soaking in the bacterial suspension for 3 h, three seeds were placed on the filter paper. Filter papers were then rolled up and placed in the beaker containing 500 mL of sterile water. After one week of cultivation, seedlings were thinned down to one on each filter paper. Every two weeks, 10 mL of nutrient solution [
15] was supplemented. After 25 days of cultivation, 5 mL of the bacterial suspension was re-inoculated in bacterial inoculation treatments. Regarding the uninoculated treatment, which represented the control, only 5 mL of YEM was supplemented. Six replications were conducted for each treatment. After 45 days of cultivation, the number of nodules and the growth of soybean including root length, shoot length, and number of leaves were determined. Symbiotic nitrogen-fixing activities of nodules were assessed as described in [
22] using an acetylene reduction assay [
23].
2.4. Determination of Plant Growth-Promoting and Plant Constituent-Hydrolyzing Traits of Isolate Bra6 and Non-Rhizobial Isolates
The three-day fresh colonies grown on YEMA were subjected to plant growth-promoting and plant constituent-hydrolyzing characterization. The free-living nitrogen-fixing activities of the isolates were assessed as described in [
16] using an acetylene reduction assay [
23]. To test for isolates with phosphate-solubilizing activities, colonies were picked and inoculated on/in tricalcium phosphate-containing medium, which was modified from Pikovskaya medium [
24] and contained (L
−1 distilled water: glucose 10 g, (NH
4)
2SO
4 0.5 g, Ca
3(PO
4)
2 5 g, KCl 0.2 g, MgSO
4 · 7H
2O, 0.1 g, MnSO
4 · 5H
2O 0.001 g, FeSO
4 · 7H
2O 0.001 g, yeast extract 0.5 g, the pH was adjusted to 7.0 before autoclaving, and 2.5% of bacteriological agar powder was added when solid plates were prepared). To test for isolates with IAA-producing activities, colonies were picked and inoculated in YEM (pH 5 and 7) supplemented with 500 µg tryptophan mL
−1. The phosphate-solubilizing and IAA-producing activities of the five-day cultures were determined by the colorimetric method as described in [
25]. Production of siderophore was evaluated on CAS agar plate as described in [
26].
The decomposition of plant constituents was assessed by culturing the substrates tested into nutrient agar (Difco, Detroit, MI, USA) and inoculating the medium with isolates. The cellulolytic, pectinolytic, and amylolytic activities of isolates were evaluated by growing them on media supplemented with 0.5% of carboxymethylcellulose, pectin, and soluble starch, respectively. As for proteolytic activity determination, 1.5% of skimmed milk-containing nutrient agar was prepared. Bacterial cultures were cultivated at 30 °C for five days. The appearance of a clear zone was measured after the addition of specific reagents as described in [
25].
2.5. Co-Inoculation Test in Sterile Water Agar through Flask Experiment
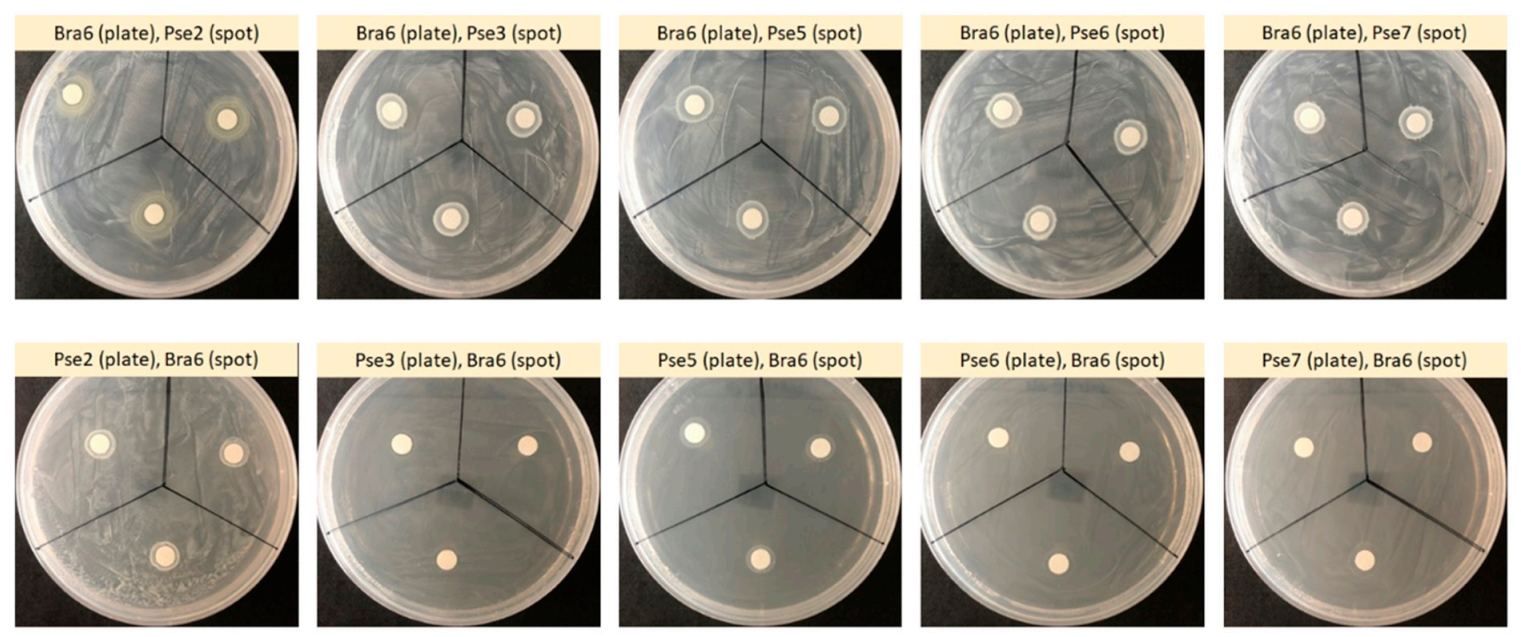
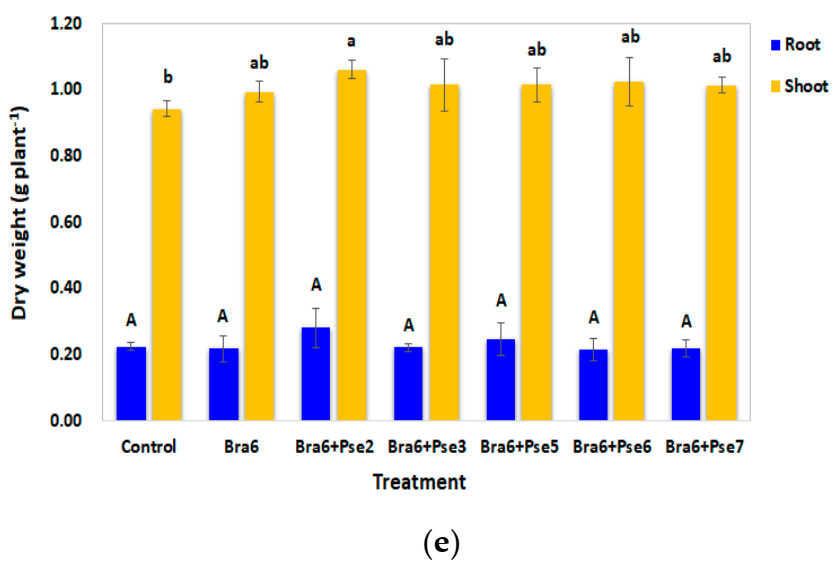
A preliminary co-cultural analysis was conducted to assess the compatibility of Bradyrhizobium isolate Bra6 and various Pseudomonas isolates. The three-day fresh cultures of each tested isolate were spread on the YEMA, and three paper discs filled with another tested cultures were placed on the same YEMA. After seven days of cultivation, the growths of both tested isolates were evaluated.
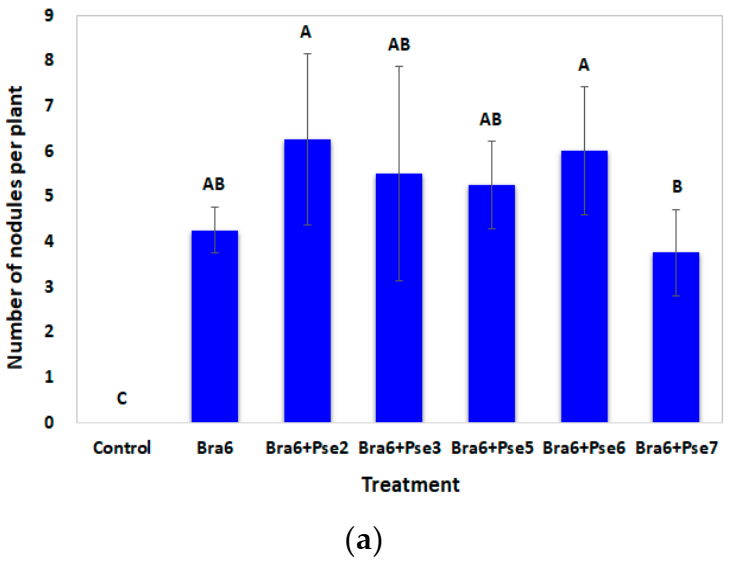
In the co-inoculation test, soybean was grown in sterile water agar-containing flasks for 60 days in greenhouse. The treatments included an uninoculated YEM control, sole inoculation of isolate Bra6, co-inoculation with Bra6, and various Pseudomonas isolates (Bra6 + Pse2, Bra6 + Pse3, Bra6 + Pse5, Bra6 + Pse6, Bra6 + Pse7). Seeds were surface sterilized with 2% of sodium hypochlorite for 5 min followed by 70% of ethanol for 5 min. Another rinse with sterile water was repeated five times and the final rinse water was used to assess the sterilization process. After soaking in the three-day cultures for 30 min, one seed was placed in the flask containing 200 mL of 0.7% water agar. After two weeks of cultivation, 2 mL of nutrient solution was supplemented. After three weeks of cultivation, 2 mL of the bacterial suspension was re-inoculated in bacterial inoculation treatments. As for the uninoculated treatment, only 2 mL of YEM was supplemented. Four replications were conducted for each treatment.
After 60 days of cultivation the number of nodules, fresh weight and dry weight (drying at 70 °C until a constant weight was obtained) of roots and shoots were determined. Plant N was measured by the Kjeldahl digestion method [
27], as described in [
28]. The P, K, Ca, Mg, S, Fe, Mn, Cu, Zn, B, Na, and Al contents of the dried and powdered plant materials were digested in a mixture (5:1) of nitric acid and perchloric acid at 120–180 °C [
29] before measurements by ICP-OES ULTIMA 2C with a sequential JY 138 ULTRACE spectrometer (HORIBA Jobin Yvon Inc., Edison, NJ, USA).
2.6. Co-Inoculation Test in Non-Sterile Strongly Acidic Soils through Pot Experiment
In another co-inoculation test, soybean was grown in strongly acidic soil-containing pots for 65 days in greenhouse. The properties of the soils were analyzed according to the procedures described in [
9], and the results were as follows: pH, 4.06; EC, 0.098 dS m
−1; total N, 0.11%; available P, 175.11 mg kg
−1; available K, 176.33 mg kg
−1; available Ca, 441.37 mg kg
−1; available Mg, 44.22 mg kg
−1; available Fe, 554.35 mg kg
−1; available Mn, 16.82 mg kg
−1; available Cu, 1.6 mg kg
−1; available Zn, 2.07 mg kg
−1; and available B, 0.5 mg kg
−1. The treatments included an uninoculated YEM control, sole inoculation of isolate Bra6, co-inoculation with Bra6, and various
Pseudomonas isolates (Bra6 + Pse2, Bra6 + Pse3, Bra6 + Pse5, Bra6 + Pse6, Bra6 + Pse7).
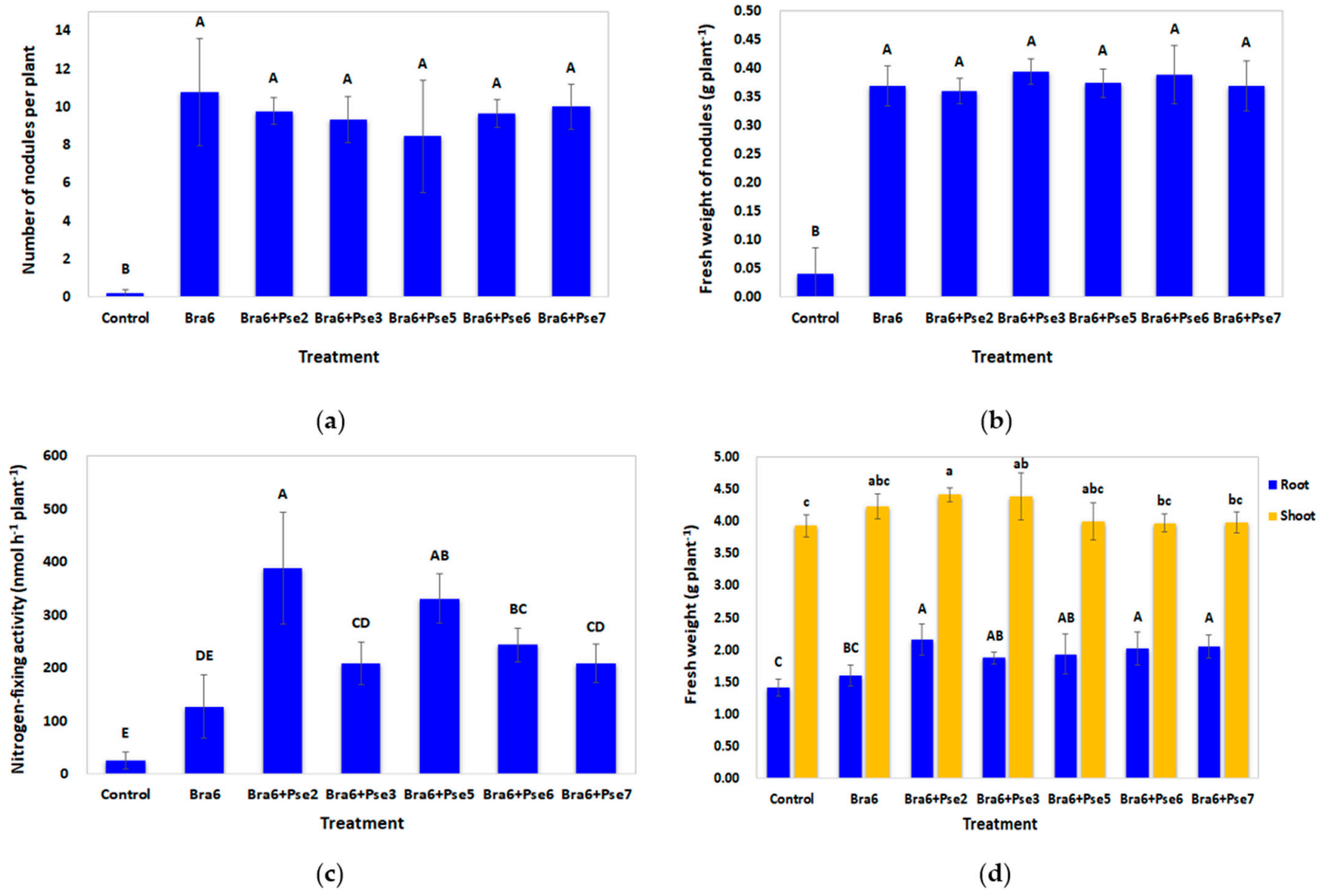
Seeds were surface sterilized and soaked in the three-day cultures as described previously. Five seeds were sown at a depth of 2 cm in the plastic pot (diameter, 10 cm; depth, 8.5 cm) containing 500 g of acidic soils. The pots were sprinkled with water. After one week of cultivation, seedlings were thinned down to three in each pot. Every week, 10 mL of nutrient solution was supplemented. After two weeks of cultivation, 2 mL of the bacterial cultures was re-inoculated in bacterial inoculation treatments. Regarding the uninoculated treatment representing the control, only 2 mL of YEM was supplemented. Three replications were conducted for each treatment. After 65 days of cultivation, the number and fresh weight of nodules, as well as fresh weight and dry weight of roots and shoots, were determined. Symbiotic nitrogen-fixing activity of nodules and nutrients of the above ground part were measured as described previously.
The results obtained for symbiotic nitrogen-fixing activity and plant growth were presented as mean values. One-way ANOVA (analysis of variance) and Duncan’s test (p < 0.05) were used to evaluate the significant differences between treatments using XLSTAT statistical software (New York, USA).
4. Discussion
The genus
Sinorhizobium has been found to be a dominant group in nodules of soybean grown in alkaline (pH 7.6–8.0) soils, whereas
Bradyrhizobium was widely distributed and dominated in neutral to slightly acidic (pH 6.9–7.7) soils [
30]. Comparative genomic analysis further revealed that genus-specific genes, known to be involved in alkaline–saline adaptations, likely contributed to the observed biogeographic patterns of these two genera in soybean [
31]. Zhang et al. [
10] also proposed that the grouping of
Ensifer- (the heterotypic synonym of
Sinorhizobium) and
Bradyrhizobium-dominated clusters from soybean nodules was significantly related to soil pH. Recently,
Bacillus cereus group was shown to promote and suppress the growth of
Sinorhizobium and
Bradyrhizobium, respectively, which affected rhizobial colonization and soybean nodulation under saline–alkali (pH 8) conditions [
32]. Bakari et al. [
33] demonstrated that soybean inoculation with
Bradyrhizobium strains effectively increased nodulation and nitrogen fixation in moderately acidic (pH 5.6–5.9) soils. Furthermore, co-application of lime and rhizobial inoculation showed potential to increase nodulation and nitrogen fixation in strongly acidic (pH 4.3–4.8) soils. In this study, the acidotolerant isolate Bra6, which shared high similarity with
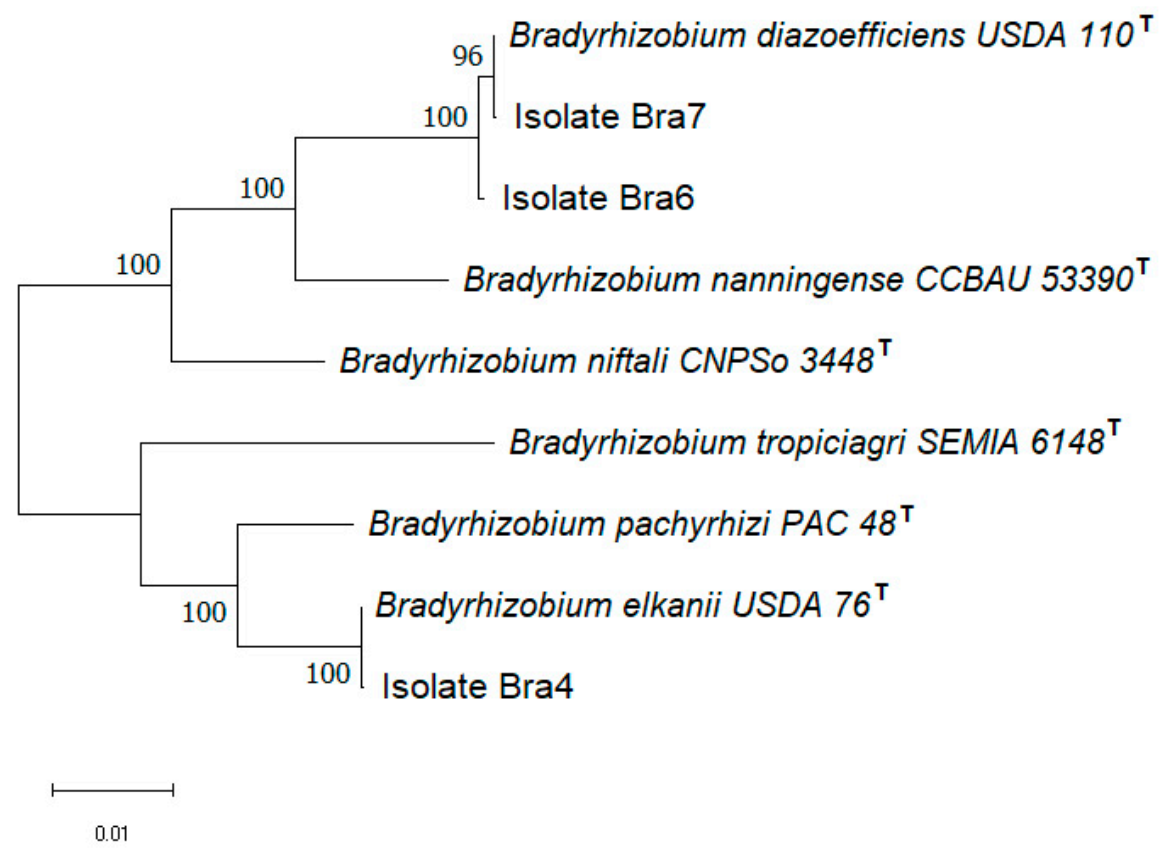
Bradyrhizobium diazoefficiens USDA 110
T in 16S rDNA (99.6%),
nifH (100%), and
nodD (99.7%), was recognized as the potential inoculant to induce nodulation of soybean in the strongly acidic (pH 4.06) soils. Previously, several strains belonging to
Bradyrhizobium diazoefficiens have also been found to nodulate and fix nitrogen when in symbiosis with soybean [
34]. Besides, the genetic diversity and distribution of
Bradyrhizobium diazoefficiens were explored in the rhizosphere of soybean grown in red soil [
35].
Despite the fact that
Bradyrhizobium spp. dominated in soybean nodules, many endophytic bacteria within nodules also showed influences on the soybean–microbe symbiosis and plant growth.
Pseudomonas has been found as the dominant non-rhizobial group associated with soybean nodules [
36]. The synergism between nodule endophyte
Pseudomonas aeruginosa and
Bradyrhizobium sp. LSBR-3 was studied, which showed the improvement in plant growth and nutrient acquisition in soybean after dual inoculation [
37]. Co-inoculation with
Bacillus thuringiensis and
Bradyrhizobium japonicum was demonstrated to enhance soybean nodulation and growth [
38,
39]. The number and weight of nodules increased, and root weight increased more often than shoot weight under greenhouse and field conditions. Compared with the uninoculated control, we also demonstrated a significant (
p < 0.05) increment in the biomass of nodules and fresh weight of roots after co-inoculation with
Bradyrhizobium and
Pseudomonas isolates in strongly acidic soils.
Pseudomonas isolates with potentially plant growth-promoting traits showed different influences on rhizobia-mediated symbiotic nitrogen fixation and soybean growth. Most of the
Pseudomonas species closely related to our isolates have not been reported as soybean-associated bacteria, except that
Pseudomonas glycinae MS586
T was isolated from the soybean rhizosphere [
40]. In this study, the non-rhizobial isolates Pse2, which shared the highest sequence similarity of 16S rDNA with
Pseudomonas punonensis CECT 8089
T, was demonstrated as the superior partner, showing a synergistic interaction with the rhizobial isolate Bra6.
Zeffa et al. [
14] demonstrated that the effects of co-inoculation on plant growth varied according to the PGPR genus used as co-inoculant, as well as with the experimental conditions. Co-inoculation of soybean plants with seed-borne
Bacillus amyloliquefaciens and
Bradyrhizobium japonicum significantly improved nodulation, which could be due in part to the production of phytohormones [
41]. High levels of IAA produced by nodule endophyte
Bacillus megaterium or rhizobacterial
Bacillus velezensis were also considered to aid the development of mature nodules, which thereby improved the nodular nitrogen fixation [
42,
43]. Defez et al. [
44] further demonstrated that IAA-overproducing
Ensifer meliloti inside nodules increased the activity of nitrogen-fixing apparatus and photosynthetic function. Besides,
Pseudomonas fluorescens has been found to enhance the nitrogen fixation of soybean through promoting the growth and colonization of
Bradyrhizobium japonicum [
45]. Soybean symbiotic performance was improved by co-inoculation with
Bradyrhizobium japonicum and
Azospirillum brasilense through quorum-sensing communication [
46]. In this study, all the
Pseudomonas isolates showed potential to produce IAA, which might influence the symbiotic outcome by altering phytohormonal homeostasis of soybean [
47]. The possibility that metabolites such as siderophores might enhance nodule formation has also been proposed previously [
48].
The entrance and survival of the endophytes in roots are generally in relation to their capabilities to hydrolyze plant constituents [
49,
50]. However, the constitutive release of plant cell wall-degrading enzymes such as pectinase by endophytic bacteria may confer plant pathogenicity [
51]. Huang et al. [
25] also proposed that endophytes with pectinase activity may be one of the factors contributing to the negative effects on plant growth. In this study, the pectinase activity was recorded in isolates Pse5, Pse6, and Pse7. Besides, the starch, which serves as the storage form of carbohydrates in soybean plant, was only utilized by isolate Bra6 and Pse2. It is assumed that both isolates proliferate using starch instead of pectin as its carbon source, which might be more compatible to the host plant. Moreover, the proteolytic activity was found in all four
Pseudomonas isolates, except for Pse2. The ability of endophytes to hydrolyze protein might also have an influence on the rhizobium-mediated symbiotic nitrogen fixation, which needs to be clarified in the future.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}