
Investigations on the Use of Dried Food Residues as a Potential Dietary Ingredient for Cats


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets
2.2. Animals and Study Design
2.3. Calculation of the Apparent Nutrient Digestibility
2.4. Fecal Microbiota and Microbial Metabolites
2.5. Plasma and Urine Analysis
2.6. Statistical Data Analysis
3. Results
3.1. Animal Health, Body Weight, Fecal Dry Matter, Feed Intake
3.2. Apparent Nutrient Digestibility
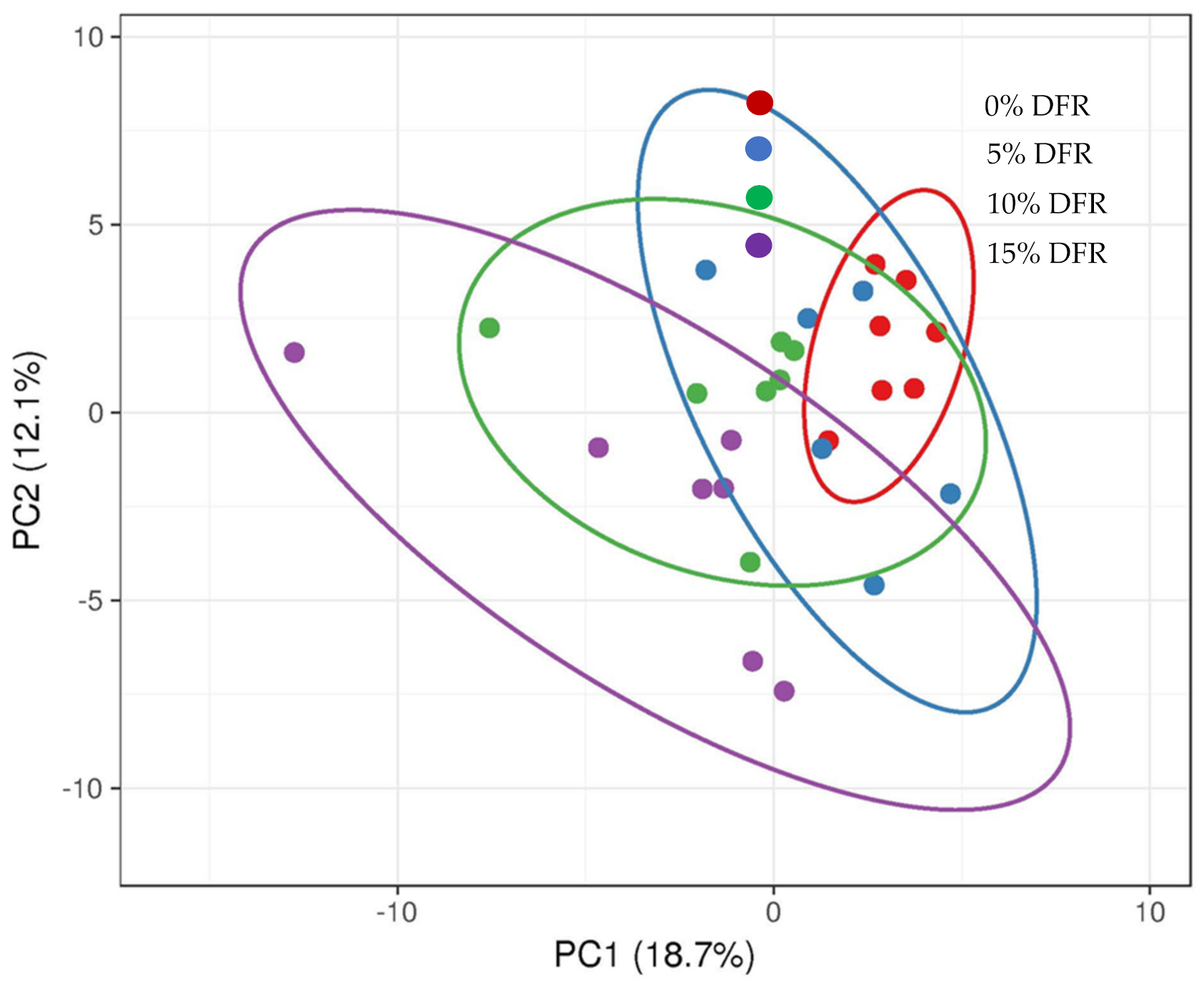
3.3. Fecal Microbiota
3.4. Fecal Microbial Metabolites
3.5. Urine pH and Composition, Phenols and Indoles in the Urine and Plasma
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Westendorf, M.L. Food waste as animal feed: An introduction. In Food Waste to Animal Feed; Westendorf, M.L., Ed.; Iowa State University Press: Ames, IA, USA, 2000; pp. 91–111. [Google Scholar]
- Regulation (EC) No 1069/2009 of the European Parliament and of the Council of 21 October 2009 Laying down Health Rules as Regards Animal by-Products and Derived Products Not Intended for Human Consumption and Repealing Regulation (EC) No 1774/2002 (Animal by-Products Regulation). 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32009R1069&from=EN (accessed on 2 August 2021).
- Commission Regulation (EU) No 142/2011 of 25 February 2011 Implementing Regulation (EC) No 1069/2009 of the European Parliament and of the Council Laying down Health Rules as Regards Animal by-Products and Derived Products Not Intended for Human Consumption and Implementing Council Directive 97/78/EC as Regards Certain Samples and Items Exempt from Veterinary Checks at the Border under that Directive 2011. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32011R0142&from=EN (accessed on 2 August 2021).
- FAO. Global Food Losses and Food Waste–Extent, Causes and Prevention; FAO: Rome, Italy, 2011; Available online: https://www.fao.org/3/mb060e/mb060e00.pdf (accessed on 2 August 2021).
- Paßlack, N.; Galliou, F.; Manios, T.; Lasaridi, K.; Tsiplakou, E.; Vahjen, W.; Zentek, J. Impact of the dietary inclusion of dried food residues on the apparent nutrient digestibility and the intestinal microbiota of dogs. Arch. Anim. Nutr. 2021, 1–17. [Google Scholar] [CrossRef]
- Bosch, G.; Hagen-Plantinga, E.A.; Hendriks, W.H. Dietary nutrient profiles of wild wolves: Insights for optimal dog nutrition? Br. J. Nutr. 2015, 113, S40–S54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council (NRC). Nutrient Requirements of Dogs and Cats; The National Academic Press: Washington, DC, USA, 2006. [Google Scholar]
- American Society for Testing and Materials (ASTM). Standard Test Method for Determination of the Composition of Unprocessed Municipal Solid Waste, ASTM D5231 (Waste Management Standards)-D5231-92(2008); ASTM International: West Conshohocken, PA, USA, 2008; Available online: https://www.astm.org/Standards/D5231.htm (accessed on 25 September 2017).
- Scientific Opinion of the Panel on Biological Hazards on a request from the Health and Consumer Protection, Directorate General, European Commission on Microbiological Risk Assessment in feeding stuffs for food-producing animals. EFSA J. 2008, 720, 1–84.
- European Feed Manufacturers’ Guide (EFMC). Community Guide to Good Practice for the Industrial EU Compound Feed and Premixtures Manufacturing Sector for Food Producing Animals. Version 1.2. November 2014. Available online: https://www.fefac.eu/wp-content/uploads/2020/07/efmc_1_2_e_final.pdf (accessed on 15 June 2021).
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs. 2005. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32005R2073&from=EN (accessed on 15 June 2021).
- Passlack, N.; Zentek, J. Urinary Calcium and Oxalate Excretion in Healthy Adult Cats Are Not Affected by Increasing Dietary Levels of Bone Meal in a Canned Diet. PLoS ONE 2013, 8, e70530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Paßlack, N.; Kohn, B.; Vahjen, W.; Zentek, J. Effects of dietary cellobiose on the intestinal microbiota and excretion of nitrogen metabolites in healthy adult dogs. J. Anim. Physiol. Anim. Nutr. 2021. [Google Scholar] [CrossRef]
- Paßlack, N.; Vahjen, W.; Zentek, J. Dietary inulin affects the intestinal microbiota in sows and their suckling piglets. BMC Vet. Res. 2015, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Paßlack, N.; Zentek, J.; Larsen, J.A.; Westropp, J.L.; Fascetti, A.J. Impact of hyperlipidaemia on intermediary metabolism, faecal microbial metabolites and urinary characteristics of lipoprotein lipase deficient vs. normal cats. J. Anim. Physiol. Anim. Nutr. 2018, 102, e139–e146. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.N.; Chou, C.C.; Tsai, P.S.J.; Lee, Y.J. Plasma indoxyl sulfate concentration predicts progression of chronic kidney disease in dogs and cats. Vet. J. 2018, 232, 33–39. [Google Scholar] [CrossRef]
- Eisenhauer, L.; Vahjen, W.; Dadi, T.; Kohn, B.; Zentek, J. Effects of brewer’s spent grain and carrot pomace on digestibility, fecal microbiota, and fecal and urinary metabolites in dogs fed low or high protein diets. J. Anim. Sci. 2019, 97, 4124–4133. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. Clustvis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Marsilio, S.; Pilla, R.; Sarawichitr, B.; Chow, B.; Hill, S.L.; Ackermann, M.R.; Scot Estep, J.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Characterization of the fecal microbiome in cats with inflammatory bowel disease or alimentary small cell lymphoma. Sci. Rep. 2019, 9, 19208. [Google Scholar] [CrossRef] [Green Version]
- Summers, S.C.; Quimby, J.M.; Isaiah, A.; Suchodolski, J.S.; Lunghofer, P.J.; Gustafson, D.L. The fecal microbiome and serum concentrations of indoxyl sulfate and p-cresol sulfate in cats with chronic kidney disease. J. Vet. Intern. Med. 2019, 33, 662–669. [Google Scholar] [CrossRef]
- Kieler, I.N.; Osto, M.; Hugentobler, L.; Puetz, L.; Gilbert, M.T.P.; Hansen, T.; Pedersen, O.; Reusch, C.E.; Zini, E.; Lutz, T.A.; et al. Diabetic cats have decreased gut microbial diversity and a lack of butyrate producing bacteria. Sci. Rep. 2019, 9, 4822. [Google Scholar] [CrossRef]
- Deng, P.; Swanson, K.S. Gut microbiota of humans, dogs and cats: Current knowledge and future opportunities and challenges. Br. J. Nutr. 2015, 113, S6–S17. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Hess, J.M.; Gould, T.J.; Slavin, J.L. Prebiotic Dietary Fiber and Gut Health: Comparing the in Vitro Fermentations of Beta-Glucan, Inulin and Xylooligosaccharide. Nutrients 2017, 9, 1361. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Nanda, A.; Khadka, B. Novel molecular, structural and evolutionary characteristics of the phosphoketolases from bifidobacteria and Coriobacteriales. PLoS ONE 2017, 12, e0172176. [Google Scholar] [CrossRef]
- Saha, S.; Jeon, B.-H.; Kurade, M.B.; Govindwar, S.P.; Chatterjee, P.K.; Oh, S.-E.; Roh, H.-S.; Lee, S.S. Interspecies microbial nexus facilitated methanation of polysaccharidic wastes. Bioresour. Technol. 2019, 289, 121638. [Google Scholar] [CrossRef]
- Fekete, S.G.; Hullár, I.; Andrásofszky, E.; Kelemen, F. Effect of different fibre types on the digestibility of nutrients in cats. J. Anim. Physiol. Anim. Nutr. (Berl.) 2004, 88, 138–142. [Google Scholar] [CrossRef]
- Fischer, M.M.; Kessler, A.M.; de Sá, L.R.M.; Vasconcellos, R.S.; Roberti Filho, F.O.; Nogueira, S.P.; Oliveira, M.C.C.; Carciofi, A.C. Fiber fermentability effects on energy and macronutrient digestibility, fecal traits, postprandial metabolite responses, and colon histology of overweight cats. J. Anim. Sci. 2012, 90, 2233–2245. [Google Scholar] [CrossRef] [PubMed]
- Kienzle, E.; Schuhknecht, A. Struvite stone dietetics: 1. Effect of different feed rations on the urine pH value of cats. Dtsch. Tierarztl. Wochenschr. 1993, 100, 198–203. [Google Scholar]
- Glorieux, G.; Gryp, T.; Perna, A. Gut-Derived Metabolites and Their Role in Immune Dysfunction in Chronic Kidney Disease. Toxins 2020, 12, 245. [Google Scholar] [CrossRef] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet⁻Microbe⁻Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Bermingham, E.N.; Kittelmann, S.; Henderson, G.; Young, W.; Roy, N.C.; Thomas, D.G. Five-week dietary exposure to dry diets alters the faecal bacterial populations in the domestic cat (Felis catus). Br. J. Nutr. 2011, 106, S49–S52. [Google Scholar] [CrossRef] [Green Version]
- Alltech’s 10th-Annual Global Feed Survey. 2021. Available online: https://one.alltech.com/wp-content/uploads/2021/01/2021_alltech_global_feed_survey-1.pdf (accessed on 9 August 2021).


{kind=link}
| Ingredient (%) | 0% DFR | 5% DFR | 10% DFR | 15% DFR |
|---|---|---|---|---|
| Ground beef | 81.0 | 78.9 | 76.5 | 75.1 |
| Rice flour | 12.4 | 10.52 | 8.51 | 5.59 |
| Rapeseed oil | 3.18 | 2.56 | 2.09 | 1.68 |
| Cellulose | 0.80 | 0.56 | 0.39 | 0.17 |
| Vitamin and Mineral premix | 0.44 | 0.45 | 0.46 | 0.48 |
| Potassium hydrogen carbonate | 0.20 | 0.14 | 0.08 | - |
| Sodium chloride | 0.30 | 0.17 | 0.08 | - |
| Blood meal | 0.51 | 0.60 | 0.70 | 0.80 |
| Magnesium supplement | 0.02 | 0.02 | 0.01 | - |
| B Vitamin Supplement | 0.07 | 0.08 | 0.10 | 0.08 |
| Dicalcium phosphate | 0.12 | 0.09 | 0.08 | - |
| Calcium carbonate | 0.58 | 0.56 | 0.50 | 0.50 |
| Cod liver oil | 0.18 | 0.19 | 0.20 | 0.20 |
| DFR 1 | - | 4.96 | 10.1 | 15.2 |
| Titanium dioxide | 0.20 | 0.20 | 0.20 | 0.20 |
| 0% DFR 1 | 5% DFR | 10% DFR | 15% DFR | |
|---|---|---|---|---|
| g/100 g | ||||
| Dry matter | 42.0 | 43.2 | 44.6 | 45.3 |
| g/100 g dry matter | ||||
| Crude ash | 5.07 | 5.02 | 5.23 | 5.15 |
| Crude protein | 41.8 | 41.8 | 41.5 | 42.3 |
| Crude fat | 24.8 | 25.0 | 25.5 | 26.4 |
| Crude fiber | 1.22 | 1.36 | 1.08 | 1.57 |
| Calcium | 0.67 | 0.66 | 0.61 | 0.65 |
| Phosphorus | 0.52 | 0.53 | 0.54 | 0.53 |
| Potassium | 0.93 | 0.88 | 0.92 | 0.92 |
| Magnesium | 0.12 | 0.12 | 0.12 | 0.13 |
| Sodium | 0.45 | 0.43 | 0.48 | 0.49 |
| Titanium dioxide | 0.43 | 0.45 | 0.44 | 0.44 |
| mg/100 g dry matter | ||||
| Copper | 1.02 | 1.15 | 1.21 | 1.18 |
| Zinc | 20.6 | 20.5 | 18.6 | 19.3 |
| Iron | 13.3 | 14.2 | 14.3 | 15.3 |
| Manganese | 1.75 | 1.77 | 1.63 | 1.71 |
| 0% DFR | 5% DFR | 10% DFR | 15% DFR | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|
| Lin. | Quadr. | |||||
| Feed intake (g dry matter/kg body weight/day) | 10.1 ± 1.51 | 10.1 ± 1.27 | 9.60 ± 1.16 | 9.33 ± 1.25 | 0.169 | 0.661 |
| Body weight (kg) | 5.36 ± 1.14 | 5.79 ± 1.19 | 5.99 ± 1.08 | 5.95 ± 1.17 | 0.135 | 0.092 |
| Fecal dry matter (%) | 44.4 ± 11.5 | 41.0 ± 11.9 | 43.6 ± 6.80 | 41.3 ± 6.10 | 0.974 | 0.335 |
| Apparent crude protein digestibility (%) | 93.9 ± 2.06 | 92.8 ± 1.90 | 91.4 ± 1.57 | 90.0 ± 1.91 | 0.007 | 0.870 |
| Apparent crude fat digestibility (%) | 97.3 ± 2.23 | 97.4 ± 0.56 | 96.5 ± 1.90 | 96.0 ± 1.31 | 0.062 | 0.196 |
| 0% DFR (n = 7) | 5% DFR (n = 6) | 10% DFR (n = 7) | 15% DFR (n = 7) | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Richness | 33.0 ± 6.38 | 36.5 ± 4.04 | 46.9 ± 5.58 | 51.4 ± 5.77 | 0.001 | 0.075 |
| Shannon | 1.46 ± 0.45 | 1.91 ± 0.21 | 2.17 ± 0.16 | 2.37 ± 0.14 | 0.011 | 0.297 |
| Evenness | 0.42 ± 0.12 | 0.53 ± 0.05 | 0.56 ± 0.03 | 0.60 ± 0.04 | 0.043 | 0.194 |
| Simpson | 0.33 ± 0.14 | 0.24 ± 0.05 | 0.18 ± 0.04 | 0.15 ± 0.04 | 0.043 | 0.310 |
| 0% DFR | n 1 | 5% DFR | n | 10% DFR | n | 15% DFR | n | Polynomial contrasts (P value) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||||||
| Actinobacteria | 42.3 ± 33.7 | (7) | 42.7 ± 31.8 | (6) | 52.8 ± 14.6 | (7) | 37.4 ± 8.19 | (7) | 0.674 | 0.008 |
| Bacteroidetes | 8.42 ± 16.3 | (5) | 8.42 ± 17.3 | (6) | 2.76 ± 2.03 | (7) | 9.07 ± 5.04 | (7) | 0.786 | 0.379 |
| Firmicutes | 51.5 ± 32.0 | (7) | 48.6 ± 28.8 | (6) | 44.3 ± 15.5 | (7) | 53.2 ± 7.36 | (7) | 0.686 | 0.031 |
| Fusobacteria | 0.08 ± 0.04 | (3) | 0.54 ± 0.45 | (3) | (0) | 0.34 ± 0.57 | (4) | * | * | |
| 0% DFR | n 1 | 5% DFR | n | 10% DFR | n | 15% DFR | n | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Lin. | Quadr. | |||||||||
| Bacteroidales | 8.41 ± 16.3 | (5) | 8.41 ± 17.3 | (6) | 2.76 ± 2.03 | (7) | 9.07 ± 5.04 | (7) | 0.786 | 0.380 |
| Bifidobacteriales | 32.0 ± 31.4 | (5) | 27.6 ± 25.0 | (5) | 21.9 ± 17.4 | (7) | 2.37 ± 3.29 | (7) | 0.231 | 0.064 |
| Clostridiales | 43.8 ± 27.7 | (7) | 44.9 ± 30.4 | (6) | 39.5 ± 16.5 | (7) | 42.1 ± 10.3 | (7) | 0.404 | 0.688 |
| Coriobacteriales | 19.4 ± 19.8 | (7) | 19.6 ± 11.5 | (6) | 30.9 ± 7.77 | (7) | 35.0 ± 7.92 | (7) | <0.001 | 0.113 |
| Erysipelotrichales | 3.22 ± 2.38 | (7) | 1.72 ± 1.17 | (6) | 2.93 ± 1.66 | (7) | 5.54 ± 3.59 | (7) | 0.051 | 0.013 |
| Fusobacteriales | 0.08 ± 0.04 | (3) | 0.54 ± 0.45 | (3) | (0) | 0.34 ± 0.57 | (4) | * | * | |
| Lactobacillales | 6.87 ± 13.1 | (4) | 1.04 | (1) | 0.03 | (2) | 0.55 ± 1.02 | (4) | * | * |
| Selenomonadales | 0.56 ± 0.72 | (5) | 1.80 ± 2.50 | (6) | 2.23 ± 1.55 | (6) | 5.25 ± 6.54 | (7) | 0.493 | 0.809 |
| Unknown Firmicutes | 0.73 | (2) | (0) | (0) | 0.05 | (2) | * | * | ||
| 0% DFR | n 1 | 5% DFR | n | 10% DFR | n | 15% DFR | n | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Lin. | Quadr. | |||||||||
| Alloprevotella | 0.74 ± 0.89 | (4) | 1.47 ± 2.51 | (5) | 0.15 ± 0.08 | (6) | 3.31 ± 4.79 | (5) | 0.538 | 0.467 |
| Bacteroides | 0.47 ± 0.46 | (5) | 0.58 ± 0.55 | (6) | 0.40 ± 0.21 | (7) | 0.73 ± 0.58 | (7) | 0.830 | 0.748 |
| Bifidobacterium | 32.0 ± 31.3 | (5) | 27.5 ± 24.9 | (5) | 21.8 ± 17.4 | (7) | 2.35 ± 3.27 | (7) | 0.230 | 0.064 |
| Blautia | 24.8 ± 20.4 | (7) | 20.8 ± 15.2 | (6) | 17.0 ± 7.61 | (7) | 16.4 ± 2.27 | (7) | 0.361 | 0.615 |
| Clostridium sensu stricto 1 | 6.89 ± 11.6 | (7) | 4.54 ± 6.81 | (6) | 1.73 ± 2.44 | (3) | 0.05 ± 0.03 | (3) | 0.298 | 0.386 |
| Collinsella | 11.0 ± 10.8 | (7) | 13.8 ± 8.62 | (6) | 23.9 ± 5.82 | (7) | 28.7 ± 8.70 | (7) | 0.003 | 0.799 |
| Faecalibacterium | (0) | 0.64 | (1) | 0.17 ± 0.16 | (6) | 0.12 ± 0.14 | (6) | * | * | |
| Fusobacterium | 0.08 ± 0.04 | (3) | 0.50 ± 0.42 | (3) | 0.34 ± 0.57 | (4) | * | * | ||
| Holdemanella | 1.18 ± 1.70 | (4) | 0.79 ± 1.04 | (5) | 1.00 ± 0.90 | (7) | 1.84 ± 1.39 | (6) | 0.175 | 0.348 |
| Lachnoclostridium | 1.68 ± 1.17 | (7) | 4.46 ± 3.38 | (6) | 3.68 ± 1.54 | (7) | 4.95 ± 1.87 | (7) | 0.027 | 0.234 |
| Lachnospira | (0) | 0.67 | (2) | 1.19 ± 1.16 | (3) | 0.34 ± 0.28 | (3) | * | * | |
| Lachnospiraceae NK4A136 group | 0.12 ± 0.07 | (5) | 0.56 ± 0.75 | (4) | 0.22 ± 0.17 | (7) | 0.17 ± 0.09 | (7) | 0.149 | 0.401 |
| Lactobacillus | (0) | 1.03 | (1) | 0.02 | (1) | 0.01 | (2) | * | * | |
| Libanicoccus | 0.12 ± 0.08 | (5) | 0.16 ± 0.14 | (5) | 0.46 ± 0.19 | (7) | 0.50 ± 0.15 | (7) | 0.003 | 0.945 |
| Marvinbryantia | 0.64 ± 0.14 | (4) | 1.37 ± 0.70 | (3) | 0.58 ± 0.33 | (5) | 0.39 ± 0.21 | (7) | 0.131 | 0.035 |
| Megamonas | 0.11 ± 0.14 | (4) | 1.11 ± 1.61 | (4) | 0.22 ± 0.30 | (5) | 0.30 ± 0.39 | (5) | 0.881 | 0.520 |
| Megasphaera | 0.26 ± 0.33 | (4) | 1.67 ± 2.52 | (3) | 1.60 ± 1.10 | (6) | 6.39 ± 6.88 | (5) | * | * |
| Olsenella | 14.1 ± 27.3 | (4) | 10.7 ± 14.0 | (3) | 6.77 ± 4.59 | (6) | 4.62 ± 5.49 | (7) | 0.914 | 0.480 |
| Paeniclostridium | 0.71 ± 0.54 | (6) | 0.27 ± 0.13 | (3) | (0) | (0) | * | * | ||
| Peptoclostridium | 3.54 ± 1.85 | (7) | 4.76 ± 4.27 | (6) | 3.65 ± 2.94 | (7) | 5.01 ± 2.65 | (7) | 0.588 | 0.812 |
| Prevotella 9 | 12.1 ± 19.3 | (3) | 6.50 ± 14.6 | (6) | 2.08 ± 1.75 | (7) | 5.68 ± 3.82 | (7) | 0.653 | 0.331 |
| Romboutsia | 0.21 ± 0.12 | (4) | 0.53 ± 0.72 | (4) | 0.22 ± 0.16 | (5) | 0.41 ± 0.23 | (4) | 0.027 | 0.396 |
| Sellimonas | 0.50 ± 0.42 | (6) | 0.77 ± 0.69 | (4) | 0.69 ± 0.54 | (7) | 0.42 ± 0.43 | (6) | 0.834 | 0.540 |
| Solobacterium | 2.28 ± 2.46 | (7) | 0.98 ± 0.77 | (5) | 1.68 ± 1.73 | (7) | 3.57 ± 2.67 | (7) | 0.384 | 0.164 |
| Streptococcus | 6.62 ± 12.6 | (4) | (0) | 0.04 | (1) | 1.06 | (2) | * | * | |
| Subdoligranulum | 0.07 | (2) | 2.50 ± 3.78 | (3) | 4.01 ± 7.57 | (7) | 3.77 ± 6.54 | (7) | * | * |
| Unknown Clostridiales Family XIII | 0.52 ± 0.39 | (6) | 0.35 ± 0.24 | (6) | 0.48 ± 0.59 | (7) | 0.98 ± 1.17 | (7) | 0.905 | 0.540 |
| Unknown Lachnospiraceae | 3.74 ± 2.26 | (7) | 4.98 ± 3.04 | (6) | 6.00 ± 2.06 | (7) | 7.03 ± 1.76 | (7) | 0.069 | 0.865 |
| 0% DFR | 5% DFR | 10% DFR | 15% DFR | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| µmol/g | ||||||
| Ammonium | 25.0 ± 6.17 | 21.4 ± 4.43 | 20.7 ± 6.96 | 23.4 ± 7.66 | 0.147 | 0.491 |
| L-lactate | 0.04 ± 0.07 | 0.15 ± 0.27 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.130 | 0.338 |
| D-lactate | 0.08 ± 0.11 | 0.30 ± 0.49 | 0.02 ± 0.03 | 0.05 ± 0.09 | 0.201 | 0.340 |
| Acetic acid | 69.4 ± 42.1 | 61.2 ± 15.2 | 67.4 ± 31.2 | 93.8 ± 49.0 | 0.188 | 0.253 |
| Propionic acid | 27.9 ± 15.5 | 29.8 ± 6.40 | 33.3 ± 11.8 | 45.8 ± 19.8 | 0.025 | 0.485 |
| i-butyric acid | 3.62 ± 1.14 | 3.22 ± 0.74 | 3.87 ± 0.96 | 4.72 ± 1.95 | 0.099 | 0.245 |
| n-butyric acid | 15.6 ± 7.78 | 13.5 ± 5.97 | 14.7 ± 3.86 | 15.2 ± 7.52 | 0.225 | 0.808 |
| i-valeric acid | 4.41 ± 1.31 | 3.57 ± 1.38 | 4.68 ± 1.21 | 5.19 ± 2.36 | 0.416 | 0.273 |
| n-valeric acid | 6.42 ± 3.13 | 5.68 ± 4.46 | 8.29 ± 2.73 | 9.19 ± 4.54 | 0.016 | 0.709 |
| Total SCFA | 127 ± 66.0 | 117 ± 22.8 | 127 ± 57.8 | 174 ± 81.8 | 0.089 | 0.215 |
| % SCFA | ||||||
| Acetic acid | 51.0 ± 9.28 | 52.1 ± 6.86 | 49.6 ± 4.72 | 52.9 ± 4.71 | 0.673 | 0.592 |
| Propionic acid | 21.4 ± 2.20 | 25.6 ± 3.32 | 25.3 ± 2.62 | 26.7 ± 2.35 | 0.003 | 0.158 |
| i-butyric acid | 3.44 ± 1.45 | 2.82 ± 0.79 | 3.08 ± 0.53 | 2.83 ± 0.62 | 0.325 | 0.463 |
| n-butyric acid | 13.8 ± 4.77 | 11.5 ± 3.88 | 11.6 ± 1.74 | 9.05 ± 2.25 | 0.032 | 0.883 |
| i-valeric acid | 4.58 ± 2.69 | 3.16 ± 1.38 | 3.77 ± 1.01 | 3.13 ± 0.88 | 0.243 | 0.571 |
| n-valeric acid | 5.77 ± 2.86 | 4.86 ± 3.48 | 6.60 ± 1.81 | 5.44 ± 1.40 | 0.401 | 0.685 |
| 0% DFR | 5% DFR | 10% DFR | 15% DFR | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|
| Lin. | Quadr. | |||||
| pH | 8.20 ± 0.22 | 7.94 ± 0.35 | 7.81 ± 0.30 | 7.67 ± 0.28 | 0.002 | 0.586 |
| µg/mL | ||||||
| Phenol | 0.14 ± 0.07 | 0.28 ± 0.16 | 0.17 ± 0.06 | 0.27 ± 0.18 | 0.386 | 0.452 |
| 4-ethylphenol | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.01 ± 0.02 | 0.05 ± 0.06 | 0.224 | 0.162 |
| 3-methylphenol | 0.10 ± 0.02 | 0.08 ± 0.03 | 0.10 ± 0.03 | 0.12 ± 0.10 | 0.447 | 0.884 |
| 7-methylphenol | 0.08 ± 0.04 | 0.03 ± 0.03 | 0.02 ± 0.01 | 0.01 ± 0.02 | 0.007 | 0.037 |
| p-cresol | 0.53 ± 0.47 | 0.62 ± 0.24 | 1.20 ± 0.35 | 1.78 ± 0.70 | 0.008 | 0.848 |
| Indole | 0.21 ± 0.48 | 0.08 ± 0.07 | 0.15 ± 0.10 | 0.19 ± 0.08 | 0.768 | 0.381 |
| Indoxyl sulfate | 2.43 ± 0.63 | 1.63 ± 1.17 | 2.02 ± 0.88 | 2.26 ± 1.17 | 0.315 | 0.069 |
| 0% DFR | 5% DFR | 10% DFR | 15% DFR | Polynomial Contrasts (P Value) | ||
|---|---|---|---|---|---|---|
| Lin. | Quadr. | |||||
| µg/mL | ||||||
| p-cresol | 0.52 ± 0.51 | 0.67 ± 0.92 | 0.57 ± 0.39 | 0.75 ± 0.45 | 0.373 | 0.945 |
| Phenol | 0.59 ± 0.17 | 0.53 ± 0.01 | 0.53 ± 0.01 | 0.56 ± 0.02 | 0.570 | 0.292 |
| mg/mL | ||||||
| Indoxyl sulfate | 3.01 ± 1.04 | 1.56 ± 1.31 | 2.82 ± 1.66 | 3.03 ± 1.07 | 0.033 | 0.037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paßlack, N.; Galliou, F.; Manios, T.; Papadaki, A.; Markakis, N.; Sambathianakis, I.; Lasaridi, K.; Fortatos, S.; Kyriacou, A.; Vahjen, W.; et al. Investigations on the Use of Dried Food Residues as a Potential Dietary Ingredient for Cats. Sustainability 2021, 13, 11603. https://0-doi-org.brum.beds.ac.uk/10.3390/su132111603
Paßlack N, Galliou F, Manios T, Papadaki A, Markakis N, Sambathianakis I, Lasaridi K, Fortatos S, Kyriacou A, Vahjen W, et al. Investigations on the Use of Dried Food Residues as a Potential Dietary Ingredient for Cats. Sustainability. 2021; 13(21):11603. https://0-doi-org.brum.beds.ac.uk/10.3390/su132111603
Chicago/Turabian StylePaßlack, Nadine, Fenia Galliou, Thrassyvoulos Manios, Akrivi Papadaki, Nikolaos Markakis, Ioannis Sambathianakis, Katia Lasaridi, Stathis Fortatos, Adamadini Kyriacou, Wilfried Vahjen, and et al. 2021. "Investigations on the Use of Dried Food Residues as a Potential Dietary Ingredient for Cats" Sustainability 13, no. 21: 11603. https://0-doi-org.brum.beds.ac.uk/10.3390/su132111603