
Spatial Patterns and Scales of Collembola Taxonomic and Functional Diversity in Urban Parks


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Design
2.2. Species Traits
2.3. Data Analysis
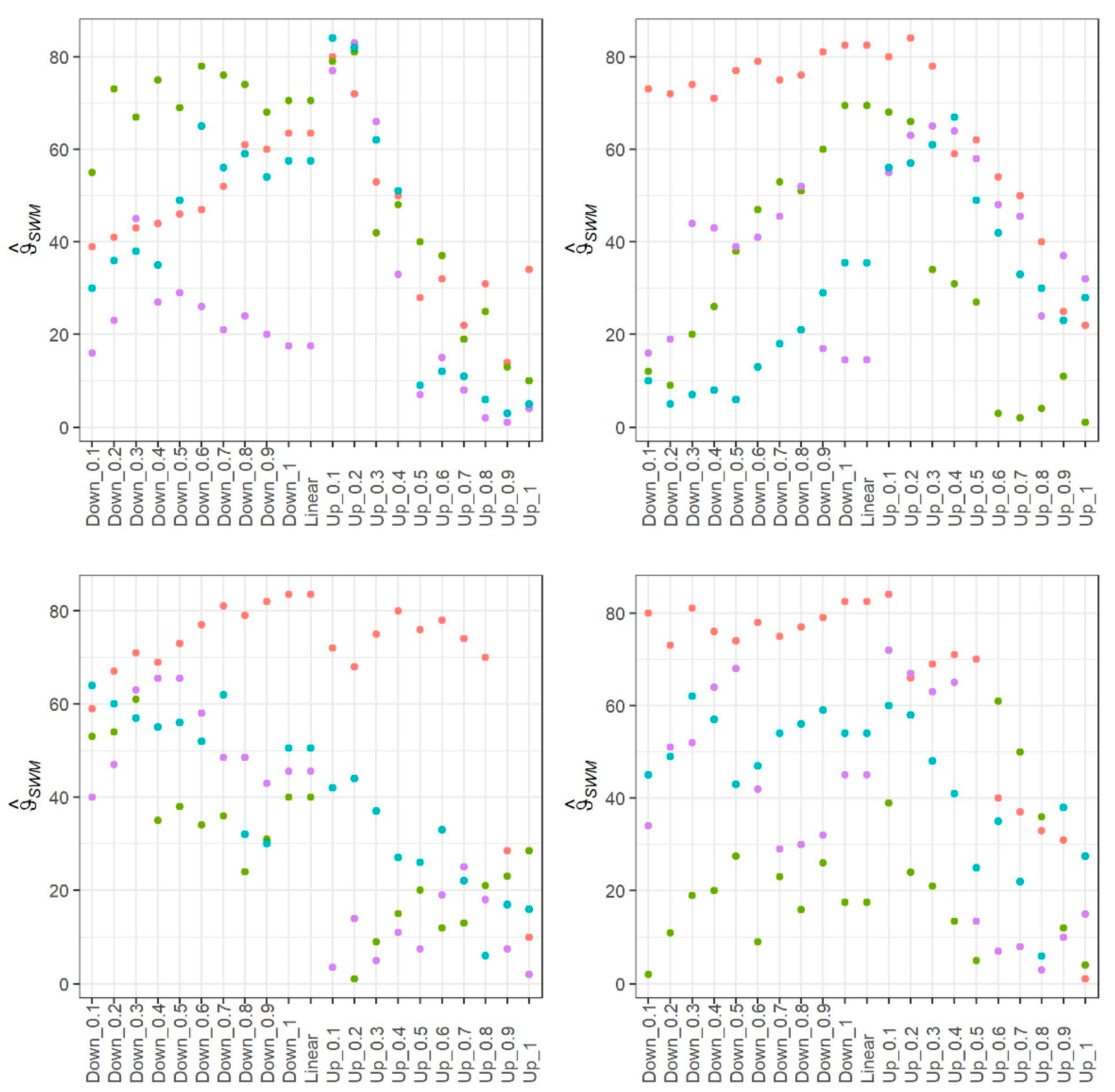
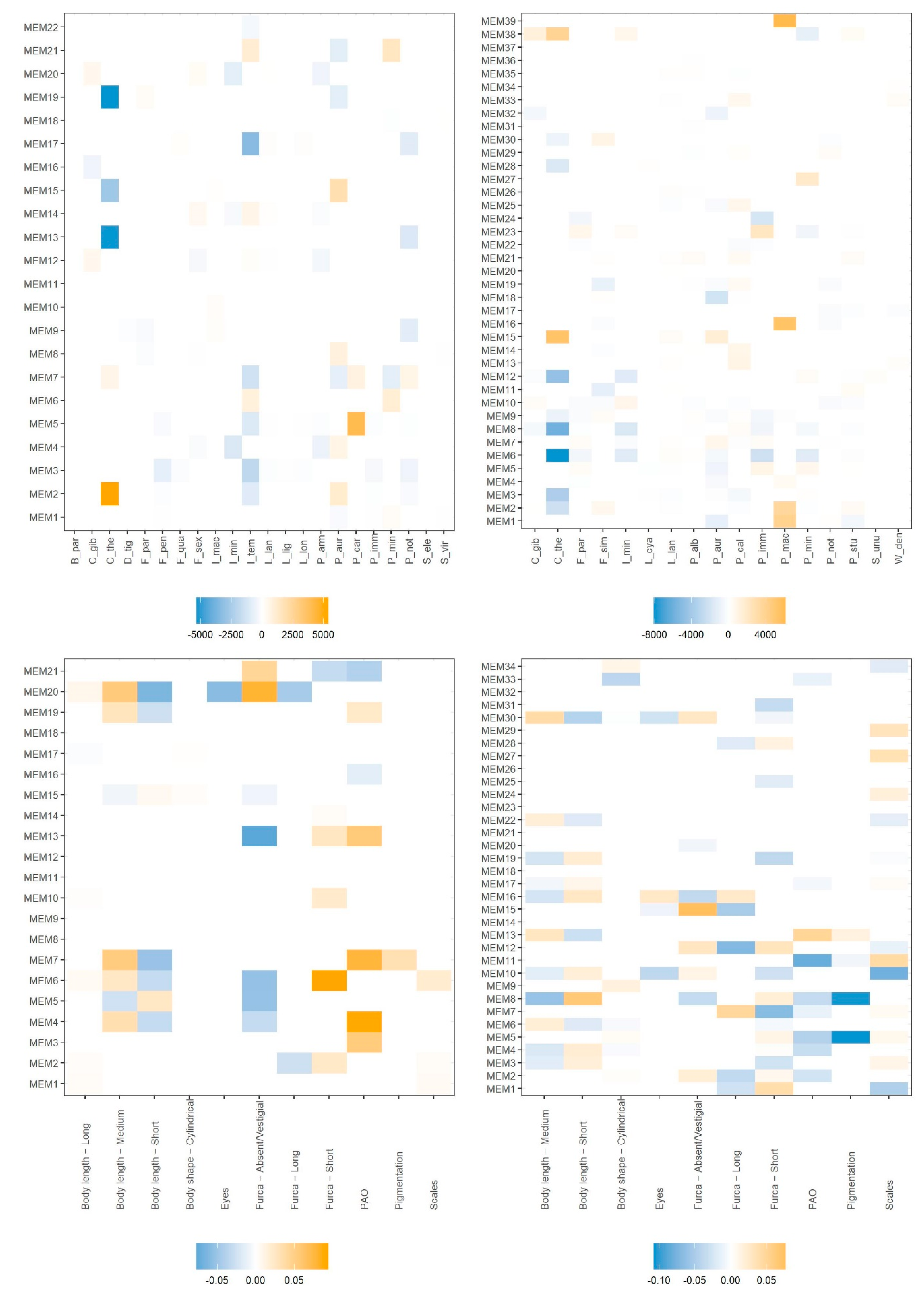
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vergnes, A.; Le Viol, I.; Clergeau, P. Green corridors in urban landscapes affect the arthropod communities of domestic gardens. Biol. Conserv. 2012, 145, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Morel, J.L.; Chenu, C.; Lorenz, K. Ecosystem services provided by soils of urban, industrial, traffic, mining, and military areas (SUITMAs). J. Soils Sediments 2015, 15, 1659–1666. [Google Scholar] [CrossRef]
- Guilland, C.; Maron, P.A.; Damas, O.; Ranjard, L. Biodiversity of urban soils for sustainable cities. Environ. Chem. Lett. 2018, 16, 1267–1282. [Google Scholar] [CrossRef]
- Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Junior, W.; Wilke, A.B.B.; Marrelli, M.T. Mosquitoes in urban green spaces: Using an island biogeographic approach to identify drivers of species richness and composition. Sci. Rep. 2017, 7, 17826. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S.; Mantoni, C.; De Simoni, L.; Galassi, D. Island biogeography of insect conservation in urban green spaces. Environ. Conserv. 2018, 45, 1–10. [Google Scholar] [CrossRef]
- Mardiastuti, A. Urban park design for bird diversity: Theory and application in landscape and site scales. IOP Conf. Ser. Earth Environ. Sci. 2020, 501, 012026. [Google Scholar] [CrossRef]
- Bräuniger, C.; Knapp, S.; Kühn, I.; Klotz, S. Testing taxonomic and landscape surrogates for biodiversity in an urban setting. Landsc. Urban Plan. 2010, 97, 283–295. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, Y.; Li, S.; von Gadow, K. The effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban For. Urban Green. 2015, 14, 1027–1039. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Williams, N.S.G.; Hahs, A.K.; Livesley, S.J. Approaches to urban vegetation management and the impacts on urban bird and bat assemblages. Landsc. Urban Plan. 2016, 153, 28–39. [Google Scholar] [CrossRef]
- Vergnes, A.; Pellissier, V.; Lemperiere, G.; Rollard, C.; Clergeau, P. Urban densification causes the decline of ground-dwelling arthropods. Biodivers. Conserv. 2014, 23, 1859–1877. [Google Scholar] [CrossRef]
- Amossé, J.; Dózsa-Farkas, K.; Boros, G.; Rochat, G.; Sandoz, G.; Fournier, B.; Mitchell, E.A.D.; Le Bayon, R.-C. Patterns of earthworm, enchytraeid and nematode diversity and community structure in urban soils of different ages. Eur. J. Soil Biol. 2016, 73, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Francini, G.; Hui, N.; Jumpponen, A.; Kotze, D.J.; Romantschuk, M.; Allen, J.A.; Setälä, H. Soil biota in boreal urban greenspace: Responses to plant type and age. Soil Biol. Biochem. 2018, 118, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Milano, V.; Cortet, J.; Baldantoni, D.; Bellino, A.; Dubs, F.; Nahmani, J.; Strumia, S.; Maisto, G. Collembolan biodiversity in Mediterranean urban parks: Impact of history, urbanization, management and soil characteristics. Appl. Soil Ecol. 2017, 119, 428–437. [Google Scholar] [CrossRef]
- Milano, V.; Maisto, G.; Baldantoni, D.; Bellino, A.; Bernard, C.; Croce, A.; Dubs, F.; Strumia, S.; Cortet, J. Impact of the neighboring cityscape on Collembola biodiversity: A Mediterranean case study. Landsc. Urban Plan. 2018, 180, 135–147. [Google Scholar] [CrossRef]
- Li, X.; Zhu, H.; Geisen, S.; Bellard, C.; Hu, F.; Li, H.; Chen, X.; Liu, M. Agriculture erases climate constraints on soil nematode communities across large spatial scales. Glob. Change Biol. 2020, 26, 919–930. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Evolutionary diversification and the origin of the diversity-environment relationship. Ecology 2006, 87, S3–S13. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015; International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- ISO 23611-2. Soil Quality. Sampling of Soil Invertebrates. Part 2. Sampling and Extraction of Micro-Arthropods (Collembola and Acarina); International Organization for Standardization: Geneva, Switzerland, 2006. [Google Scholar]
- Macfadyen, A. Improved funnel-type extractors for soil arthropods. J. Anim. Ecol. 1961, 30, 171–184. [Google Scholar] [CrossRef]
- Pey, B.; Nahmani, J.; Auclerc, A.; Capowiez, Y.; Cluzeau, D.; Cortet, J.; Decaëns, T.; Deharveng, L.; Dubs, F.; Joimel, S.; et al. Current use of and future needs for soil invertebrate functional traits in community ecology. Basic Appl. Ecol. 2014, 15, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Bretfeld, G. Synopses on Palaeartic Collembola Pt 2: Symphpleona; Staatliches Museum für Naturkunde: Görlitz, Germany, 1999. [Google Scholar]
- Dunger, W.; Schlitt, B. Synopses on Palaeartic Collembola Pt 6/1: Tullbergiidae; Senckenberg Museum of Natural History: Görlitz, Germany, 2011. [Google Scholar]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola); Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Jordana, R. Synopses on Palaeartic Collembola Pt 7/1: Capryinae & Entomobryini; Senckenberg Museum of Natural History: Görlitz, Germany, 2012. [Google Scholar]
- Potapov, M. Synopses on Palaearctic Colembola Pt 3: Isotomidae; Museum of Natural History: Görlitz, Germany, 2001. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; da Gama Assalino, M.M. Synopses on Palaeartic Collembola Pt 4: Hypogastruridae; Museum of Natural History: Görlitz, Germany, 2004. [Google Scholar]
- Zimdars, B.; Dunger, W. Synopses on Palaeartic Collembola Pt 1: Tullbergiinae; Museum of Natural History: Görlitz, Germany, 1994. [Google Scholar]
- Gisin, H. Okologie und Lebengemeinschaften der Collembolen im Schweizerischen Exkursionsgebiet Basels. Rev. Suisse Zool. 1943, 50, 131–224. [Google Scholar]
- Joimel, S.; Schwartz, C.; Bonfanti, J.; Hedde, M.; Krogh, P.H.; Pérès, G.; Pernin, C.; Rakoto, A.; Salmon, S.; Santorufo, L.; et al. Functional and taxonomic diversity of Collembola as complementary tools to assess land use effects on soils biodiversity. Front. Ecol. Evol. 2021, 9, 450. [Google Scholar] [CrossRef]
- Hawes, T.C. The unusually colourful ‘clothing’ changes of Lepidophorella australis Carpenter, 1925 (Collembola: Tomoceridae). N. Z. Entomol. 2017, 40, 22–29. [Google Scholar] [CrossRef]
- Podani, J. Extending Gower’s general coefficient of similarity to ordinal characters. Taxon 1999, 48, 331. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Bill, S.G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.R.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Bellino, A.; Alfani, A.; De Riso, L.; Gregorio, R.; Pellegrino, T.; Baldantoni, D. Long-established and new active biomonitors jointly reveal potentially toxic element gradients across spatial scales in freshwater ecosystems. Ecol. Indic. 2020, 118, 106742. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Dray, S.; Dufour, A.-B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zinger, L.; Taberlet, P.; Schimann, H.; Bonin, A.; Boyer, F.; De Barba, M.; Gaucher, P.; Gielly, L.; Giguet-Covex, C.; Iribar, A.; et al. Body size determines soil community assembly in a tropical forest. Mol. Ecol. 2019, 283, 528–543. [Google Scholar] [CrossRef]
- Piano, E.; Souffreau, C.; Merckx, T.; Baardsen, L.F.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob. Change Biol. 2020, 26, 1196–1211. [Google Scholar] [CrossRef]
- Bahram, M.; Kohout, P.; Anslan, S.; Harend, H.; Abarenkov, K.; Tedersoo, L. Stochastic distribution of small soil eukaryotes resulting from high dispersal and drift in a local environment. ISME J. 2016, 10, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Coulson, S.J.; Hodkinson, I.D.; Webb, N.R. Microscale distribution patterns in high Arctic soil microarthropod communities: The influence of plant species within the vegetation mosaic. Ecography 2003, 26, 801–809. [Google Scholar] [CrossRef]
- Gao, M.; He, P.; Zhang, X.; Liu, D.; Wu, D. Relative roles of spatial factors, environmental filtering and biotic interactions in fine-scale structuring of a soil mite community. Soil Biol. Biochem. 2014, 79, 68–77. [Google Scholar] [CrossRef]
- Gao, M.; Liu, D.; Lin, L.; Wu, D. The small-scale structure of a soil mite metacommunity. Eur. J. Soil Biol. 2016, 74, 69–75. [Google Scholar] [CrossRef]
- Arribas, P.; Andújar, C.; Salces-Castellano, A.; Emerson, B.C.; Vogler, A.P. The limited spatial scale of dispersal in soil arthropods revealed with whole-community haplotype-level metabarcoding. Mol. Ecol. 2021, 30, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Hopkin, S.P. A Key to the Collembola (Springtails) of Britain and Ireland; Field Studies Council: Telford, UK, 2007. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark—Part II: Entomobryomorpha and Symphypleona; Fauna Entomologica Scandinavica; Brill: Leiden, The Netherlands, 2007; Volume 42. [Google Scholar]
- Collins, G.E.; Hogg, I.D.; Convey, P.; Barnes, A.D.; McDonald, I.R. Spatial and temporal scales matter when assessing the species and genetic diversity of springtails (Collembola) in Antarctica. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Dunger, W.; Schulz, H.-J.; Zimdars, B. Colonization behaviour of Collembola under different conditions of dispersal. Pedobiologia 2002, 46, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Salmon, S.; Ponge, J.F. Species traits and habitats in springtail communities: A regional scale study. Pedobiologia 2012, 55, 295–301. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isotomidae | Entomobryidae | ||||
| Cryptopygus thermophilus Axelson | C_the | B | Lepidocyrtus cyaneus Tullberg | L_cya | M |
| Desoria tigrina Nicolet | D_tig | N | Lepidocyrtus lanuginosus Gmelin | L_lan | B |
| Folsomia penicula Bagnall | F_pen | N | Lepidocyrtus lignorum Fabricius | L_lig | N |
| Folsomia quadrioculata Tullberg | F_qua | N | Pseudosinella alba Packard | P_alb | M |
| Folsomia sexoculata Tullberg | F_sex | N | Pseudosinella immaculata Lie-Pettersen | P_imm | B |
| Folsomia similis Bagnall | F_sim | M | |||
| Folsomides parvulus Stach | F_par | B | |||
| Isotomiella minor Schaeffer | I_min | B | |||
| Isotomodes templetoni Bagnall | I_tem | N | |||
| Isotomurus maculatus Schaeffer | I_mac | N | |||
| Parisotoma notabilis Schaeffer | P_not | B | |||
| Proctostephanus stuckeni Borner | P_stu | M | |||
| Proisotoma minuta Tullberg | P_min | B | |||
| Hypogastruridae | Tullbergiidae | ||||
| Ceratophysella gibbosa Bagnall | C_gib | B | Paratullbergia callipygos Borner | P_cal | M |
| Schoettella ununguiculata Tullberg | S_unu | M | Paratullbergia caroli Luciañez, Ruiz & Simon | P_car | N |
| Willemia denisi Mills | W_den | M | Paratullbergia macdougalli Bagnall | P_mac | M |
| Sminthuridae | Onychiuridae | ||||
| Sminthurinus elegans Fitch | S_ele | N | Protaphorura armata Tullberg | P_arm | N |
| Sminthurus viridis Linnaeus | S_vir | N | Protaphorura aurantiaca Ridley | P_aur | B |
| Neanuridae | Brachystomellidae | ||||
| Lathriopyga longiseta Caroli | L_lon | N | Brachystomella parvula Schaeffer | B_par | N |
| Trait | Coding | Rationale |
|---|---|---|
| Body length | Short (<1.5 mm) | Food resource use |
| Medium (1.5–3.0 mm) | ||
| Long (>3.0 mm) | ||
| Body shape | Spherical | Habitat preference |
| Cylindrical | ||
| Furca | Absent/Vestigial | Movement/dispersion |
| Short | ||
| Long | ||
| Pigmentation | Present | Habitat preference |
| Absent | ||
| Eyes | Present | Habitat preference |
| Absent | ||
| PAO | Present | Habitat preference |
| Absent | ||
| Scales | Present | Predation avoidance |
| Absent |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellino, A.; Baldantoni, D.; Milano, V.; Santorufo, L.; Cortet, J.; Maisto, G. Spatial Patterns and Scales of Collembola Taxonomic and Functional Diversity in Urban Parks. Sustainability 2021, 13, 13029. https://0-doi-org.brum.beds.ac.uk/10.3390/su132313029
Bellino A, Baldantoni D, Milano V, Santorufo L, Cortet J, Maisto G. Spatial Patterns and Scales of Collembola Taxonomic and Functional Diversity in Urban Parks. Sustainability. 2021; 13(23):13029. https://0-doi-org.brum.beds.ac.uk/10.3390/su132313029
Chicago/Turabian StyleBellino, Alessandro, Daniela Baldantoni, Vittoria Milano, Lucia Santorufo, Jérôme Cortet, and Giulia Maisto. 2021. "Spatial Patterns and Scales of Collembola Taxonomic and Functional Diversity in Urban Parks" Sustainability 13, no. 23: 13029. https://0-doi-org.brum.beds.ac.uk/10.3390/su132313029