
Trace Metals and Diatom Stratigraphy along the Sill between Lakes Telaga Warna and Telaga Pengilon, Dieng, Central Java, Indonesia


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sediment and Core Collection
2.3. Diatom and Geochemical Analyses
2.4. Data Analysis
3. Results
3.1. Vertical Distribution of Diatoms
3.2. Zone I (150–110 cm)
3.3. Zone II (110–96 cm)
3.4. Zone III (96–60 cm)
3.5. Zone IV (60–0 cm)
3.6. Variations in Metal Concentrations
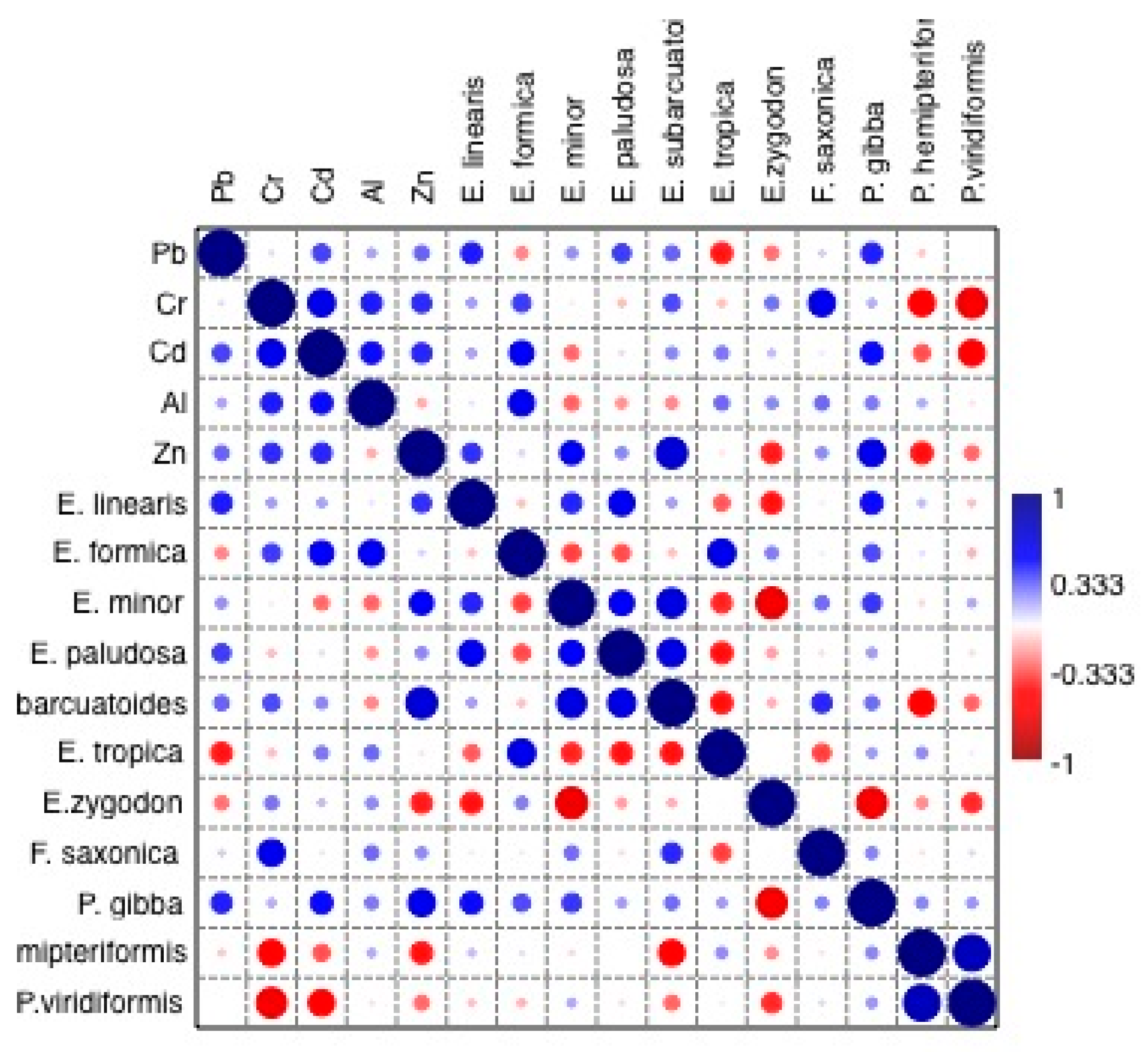
3.7. Correlation between Diatom Assemblages and Metal Concentrations
4. Discussion
4.1. Implications of Spatiotemporal Variations in Diatoms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | pH (Range) | Species | pH (Range) |
|---|---|---|---|
| Achanthes lanceolata Lange Bertalot | 2.4–3.3 | Gomphonema olivacoides Hantzsch | 3.1–3.2 |
| Coconeis placentula Ehrenberg | 2.8–3.3 | Gomphonema parvulum (Kützing) Kützing | 2.4–3.1 |
| Diatoma tenuis Agardh | 3.3 | Gomphonema subtile Ehrenberg | 3.0–3.1 |
| Eunotia bilunaris var. linearis (Okuno) Lange-Bertalot | 2.9–3.4 | Hantzschia amphioxys (Ehrenberg) Grunow | 2.8–3.5 |
| Eunotia glacialis Meister | 1.8–3.5 | Navicula crytochepala (Kützing) | 3.0–3.4 |
| Eunotia minor (Kützing) Grunow | 3 | Nitzschia Linearis (Agardh) W.Sm. | 2.8–3.2 |
| Eunotia monodon Ehrenberg | 2.9–3.4 | Nitzschia palea (Kutzing) W. Smith | 1.8–3.4 |
| Eunotia pectinalis (Ralfs) Rabenhorst | 2.8–3.4 | Pinnularia borealis Ehrenberg | 2.8–3.3 |
| Eunotia steineneckii Peterson | 3.0–3.1 | Pinnularia major (Kützing) Rabenhorst | 3.2 |
| Fragilaria construens Marciniak | 3.2 | Pinnularia subcapitata W. Gregory | 3.0–3.4 |
| Gomphonema angustatum (Kutzing) Rabenhorst | 3.3 | Eunotia bilunaris var. linearis Nitzsch | 3.2–3.4 |
4.2. Spatiotemporal Variations in Metals
5. Conclusions
- (1)
- The high abundance of acid-tolerant Eunotia species throughout the core indicate that pH fluctuated at the coring site. The observed variations in pH were presumably related to the influence of water level fluctuations at the coring site in response to both seasonal and inter-annual variations in precipitation. Variations in water levels were also likely to result in differences in the amount of hydrologic mixing between lake waters. During periods of low water, more acidic conditions characterized by minimal lake water mixing was likely. During higher water levels, pH was likely to increase, in part due to the mixing of the acidic waters in lake Telaga Warna with the higher pH waters in lake Telaga Pengilon.
- (2)
- The occurrence of diatoms with broken frustules at the bottom of Zone II with relatively high peak metal concentrations of Cd, Cr, Pb, and Zn at the top of Zone I and the bottom of Zone II suggest that lake conditions were impacted by the input of volcanic debris from a local eruption. This argument is supported by diatom assemblages found in the sediments overlying Zone II, which includes pioneering species that are known to follow eruptions in the area.
- (3)
- Metal concentrations, including Al, did not increase towards the surface of the sediments, suggesting that recent agricultural activities and the associated impact on hillslope soil erosion has not significantly impacted the lakes. This is not surprising given that in contrast to adjacent areas, the lake basin is located in a protected conservation area that is not extensively farmed.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaennudin, A. The characteristic of eruption of Indonesian active volcanoes in the last four decades. J. Lingkung. Bencana Geol. 2010, 1, 113–129. [Google Scholar]
- Edmonds, M.; Grattan, J.; Michnowicz, S. Volcanic gases: Silent killers. In Observing the Volcano World; Fearnley, C.J., Bird, D.K., Haynes, K., McGuire, W.J., Jolly, G., Eds.; Advances in Volcanology (An Official Book Series of the International Association of Volcanology and Chemistry of the Earth’s Interior–IAVCEI); Springer: Barcelona, Spain, 2015; pp. 65–89. [Google Scholar]
- Soeprobowati, T.R.; Suedy, S.W.A.; Hadiyanto. Diatoms and water quality of Telaga Warna Dieng, Java Indonesia. In Proceedings of the 2nd International Conference on Tropical and Coastal Region Eco Development, IOP Conference Series: Earth and Environmental Science, Bali, Indonesia, 25–27 October 2016; Volume 55, pp. 1–6. [Google Scholar]
- Balai Konservasi Sumber Daya Alam. Rencana Pengelolaan Jangka Panjang Taman Wisata Alam Telogo Warno Telogo Pengilon Periode 2013 Sampai Dengan 2022 Provinsi Jawa Tengah; Kementrian Kehutanan Direktorat Jendral Perlindungan Hutan dan Konservasi Alam: Semarang, Indonesia, 2021; Unpublished Work.
- Soeprobowati, T.R.; Suedy, S.W.A.; Hadiyanto; Lubis, A.A.; Gell, P. Diatom assemblage in the 24 cm upper sediment associated with human activities in Lake Warna Dieng Plateau Indonesia. Environ. Technol. Innov. 2018, 10, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Soeprobowati, T.R.; Jumari; Hariyati, R.; Gell, P. Paleolimnology record of human impact on a lake ecosystem: The case of shallow lakes in Central Java. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 276, p. 012015. [Google Scholar]
- Sari, K.; Hidayat, J.W.; Soeprobowati, T.R. Community structure and species diversity of Diatom in area between Telaga Warna and Telaga Pengilon, Indonesia. Int. Res. J. Environ. Sci. 2018, 7, 1–6. [Google Scholar]
- Hermawan, H.; Priato, E.; Setyowati, E. Evaluasi termal ruang luar desa wisata Dieng Wonosobo. J. Penelit. Pengabdi. Kpd. Masy. 2014, 2, 115–122. [Google Scholar]
- Harriyadi, H. Pertimbangan pemilihan lokasi kompleks Candi Dieng. J. Penelit. Pengemb. Arkeol. 2019, 37, 123–138. [Google Scholar] [CrossRef]
- Pradana, A.; Sara, F.H.; Wahdaningrum, W. The analysis of environmental degradation and carica agroforestry system as an attempt of environmental restoration in Dieng Plateau. Int. J. Environ. Sci. Dev. 2015, 6, 861. [Google Scholar] [CrossRef] [Green Version]
- Nurfahma, P.; Malika, R.F.; Saria, R.P.; Nurkholis, A. Influence of local wisdom to prevent disappearance of Cebong Lake in Sembungan village, Wonosobo district. In Proceedings of the International Conference on Appropriate Technology Development, Bandung, Indonesia, 5–7 October 2015; pp. 165–168. [Google Scholar] [CrossRef]
- Supriyanto, H.; Lestari, S. Danau-danau volkanik di Dataran Tinggi Dieng: Pemanfaatan dan Masalah Lingkungan Hidup yang Dihadapi. Teknosains 2015, 5, 36–48. [Google Scholar]
- Lastiantoro, C.Y.; Putra, P.B.; Andy, C.S. Dampak Penyedotan Air Telaga Dalam Usaha Tani Kentang di Telaga Pengilon Dieng Wonosobo; Muhammadiyah University Press: Surakarta, Indonesia, 2017; pp. 241–254. [Google Scholar]
- Pudjiastuti, H. Impacts of agricultural practices and tourism activities on the sustainability of Telaga Warna and Telaga Pengilon Lakes, Dieng Plateau, Central Java. In Proceedings of the 2nd International Conference on Energy, Environmental and Information System ICENIS, Semarang, Indonesia, 15–16 August 2018; Volume 31, pp. 1–6. [Google Scholar] [CrossRef]
- Liu, W.; Ma, L.; Abuduwaili, J. Anthropogenic influences on environmental changes of lake Bosten, the largest inland freshwater lake in China. Sustainability 2020, 12, 711. [Google Scholar] [CrossRef] [Green Version]
- Mitchelutti, N.; Wolfe, A.P.; Cooke, C.A.; Hobbs, W.O.; Vuille, M.; Smol, J.P. Climate change forces new ecological states in tropical andean lakes. PLoS ONE 2015, 10, e0115338. [Google Scholar] [CrossRef] [Green Version]
- Smol, J.P. Pollution of Lakes and Rivers a Paleoenvironmental Perspective, 2nd ed.; Blackwell Publishing: Hoboken, NJ, USA, 2008. [Google Scholar]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification and Use as Bioindicators; John Wiley & Sons Ltd.: Chichester, UK, 2010. [Google Scholar]
- Pardo, I.; Delgado, C.; Abraín, R.; Gómez-Rodríguez, C.; García-Roselló, E.; García, L.; Reynoldson, T.B. A predictive diatom-based model to assess the ecological status of streams and rivers of northern Spain. Ecol. Indic. 2018, 170, 519–528. [Google Scholar] [CrossRef]
- Nhiwatiwa, T.; Dalu, T.; Sithole, T. Assessment of river quality in a subtropical Austral river system: A combined approach using benthic diatoms and macroinvertebrates. Appl. Water Sci. 2017, 7, 4785–4792. [Google Scholar] [CrossRef] [Green Version]
- Sonneman, J.A.; Walsh, C.J.; Breen, P.F.; Sharpe, A.K. Effects of urbanization on streams of the Melbourne region, Victoria, Australia. II. Benthic diatom communities. Freshw. Biol. 2001, 46, 553–565. [Google Scholar] [CrossRef]
- Khudher, E.K.; Al-Jasimee, A.S. Diatoms (Bacillariophyta) as bio-indicators. Int. J. Res. Pharm. Sci. 2019, 10, 1562–1565. [Google Scholar] [CrossRef] [Green Version]
- Potapova, M.; Charles, D.F. Diatom metrics for monitoring eutrophication in rivers of the United States. Ecol. Indic. 2007, 7, 48–70. [Google Scholar] [CrossRef]
- Szczepocka, E.; Zeelazna-Wieczorek, J. Diatom biomonitoring—Scientific foundations, commonly discussed issues and fre-quently made errors. Oceanol. Hydrobiol. Stud. 2018, 47, 314–325. [Google Scholar] [CrossRef]
- Feret, L.; Bouchez, A.; Rimet, F. Benthic diatom communities in high altitude lakes: A large scale study in the French Alps. Ann. Limnol. Int. J. Limnol. 2017, 53, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Harding, W.R.; Taylor, J.C. Diatoms as Indicators of Historical Water Quality: A Comparison of Samples Taken in the Wem-mershoek Catchment (Western Province, South Africa) in 1960 and 2008; Water Research Commission: Pretoria, South Africa, 2014; pp. 601–606. [Google Scholar]
- Allard, P.; Dajlevic, D.; Delarue, C. Origin of carbon dioxide emanation from the 1979 Dieng eruption, Indonesia: Implications for the origin of the 1986 Nyos catastrophe. J. Volcanol. Geotherm. Res. 1989, 39, 195–206. [Google Scholar] [CrossRef]
- Van Bergen, J.M.; Alain, B.; Sri, S.; Terry, S.; Kastiman, S. Crater Lakes of Java: Dieng, Kelud, Ijen—Excursion Guidebook; IAVCEI General Assembly: Bali, Indonesia, 2000. [Google Scholar]
- Battarbee, R.W.; Kernan, M.; Livingstone, D.M.; Nickus, U.; Verdonschot, P.; Hering, D.; Moss, B.; Wright, R.F.; Evans, C.D.; Grimalt, J.O.; et al. Freshwater ecosystem responses to climate change: The Euro-Limpacs Project. In The Water Framework Directive-Ecological and Chemical Status Monitoring; Quevauviller, P., Borchers, U., Thompson, C., Simonart, T., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 313–354. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa, Band 2/1; Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1986; pp. 1–876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa, Band 2/2; Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1988; pp. 1–596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süsswasserflora von Mitteleropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1991; pp. 1–576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae), Gomphonema Gesamtliteraturverzeichnis Teil 1-4 [second revised edition] [With “Ergänzungen und Revisionen” by H. Lange Bertalot]. In Suesswasserflora von Mitteleuropa; Ettl, H., Ed.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2004; Volume 2, pp. 1–468. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 5. Teil: Bacillariophyceae. In Süsswasserflora von MitteleuropaII; Springer Spektrum Verlag: Jena, Germany, 2000; Volume 2, pp. 1–311. [Google Scholar]
- Taylor, J.C.; Harding, W.R.; Archibald, C.G.M. An Illustrated Guide to Some Common Diatom Species from South Africa; Water Research Commission: Pretoria, South Africa, 2007; pp. 1–225. [Google Scholar]
- Gell, P.; Tibby, J.; Fluin, P.; Leahy, M.; Reid, K.; Adamson, S.; Bulpin, A.; Macgregor, P.; Wallbrink, G.; Hancock; et al. An Illustrated Key to Common Diatom Genera from Southern Australia; CRC for Freshwater Ecology: Thurgoona, Australia, 1999; pp. 1–68. [Google Scholar]
- Sonneman, J.A. An Illustrated Guide to Common Stream Diatom Species from Temperate Australia; Cooperative Research Centre for Freshwater Ecology: Hurgoona, Australia, 2000; pp. 11–168. [Google Scholar]
- Bahls, L.L. Diatoms from Western North America 1. Some New and Notable Biraphid Species; Montana Diatom Collection; Lubrecht & Cramer: Helena, MT, USA, 2017; pp. 1–52. [Google Scholar]
- Bahls, L.; Boynton, B.; Johnston, B. Atlas of diatoms (Bacillariophyta) from diverse habitats in remote regions of western Canada. PhytoKeys 2018, 1–186. [Google Scholar] [CrossRef] [Green Version]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https:www.algaebase.org (accessed on 10 August 2020).
- Juggins, S. C2 Software for Ecological and Palaeoecological Data Analysis and Visualization; Version 1.7.7; University of New Castle: New Castle, UK, 2016; Available online: https://www.sta.ncl.ac.uk/stephen.juggins/software/C2Home.htm (accessed on 28 October 2020).
- Oksanem, F.; Guillaume, B.; Roeland, K.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.K.L.; Solymos, P.; Henry, M.H.; Wagner, S.; et al. Vegan: Community Ecology Package, R Package Version 2.0-10. 2013. Available online: http://CRAN.R-pro-ject.org/package=vegan (accessed on 21 December 2020).
- Ter Braak, C.J.F.; Juggins, S. Weighted averaging partial least squares regression (WA-PLS): An improved method for recon-structing environmental variables from species assemblages. In Twelfth International Diatom Symposium, Developments in Hydrobiology; Van Dam, H., Ed.; Springer: Dordrecht, The Netherlands, 1993; Volume 90, pp. 485–502. [Google Scholar]
- Hammer, O. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Version 2.17c. 2017. Available online: http://priede.bf.lu.lv/ftp/pub/TIS/datu_analiize/PAST/2.17c/download.html (accessed on 26 December 2020).
- Hustedt, F. Systematische und ökologische untersu-chungen über die diatomeen-flora von Java, Bali und Sumatra nach dem material der deutschen limnologischen Sunda-Expedition. Arch. Hydrobiol. Suppl. 1937, 15, 187–296. [Google Scholar]
- Schmidt, A. Atlas der Diatomaceen-Kunde 1874–1959; O.R. Reisland: Leipzig, Germany, 1909; pp. 1–120. [Google Scholar]
- Vyverman, W. Diatoms from Papua New Guinea. Bibl. Diatomol. 1995, 22, 1–244. [Google Scholar]
- Glushchenko, A.M.; Kulikovskiy, M.S. Taxonomy and distribution of the genus Eunotia Ehrenberg in water ecosystems of Vietnam. Inland Water Biol. 2017, 10, 130–139. [Google Scholar] [CrossRef]
- Taylor, J.C.; Cocquyt, C.; Mayama, S. Navicula nielsfogedii sp. nov; A new diatom (Bacillariophyta) from tropical and sub–tropical Africa. Fottea 2016, 16, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Geo Dipa Energi. INO: Geothermal Power Development Project—Dieng Unit 2 Project Component; Initial Environmental Examination; INO: Plymouth Meeting, PA, USA, 2019; pp. 1–195. [Google Scholar]
- Pudjoarianto, A.; Cushing, E.J. Pollen-stratigraphic evidence of human activity at Dieng, Central Java. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 171, 329–340. [Google Scholar] [CrossRef]
- DeNicola, D.M. A review of diatoms found in highly acidic environments. Hydrobiologia 2004, 433, 111–122. [Google Scholar] [CrossRef]
- De Faria, D.M.; Guimarães, A.T.B.; Ludwig, T.A.V. Responses of periphytic diatoms to mechanical removal of Pistia stratiotes L. in a hypereutrophic subtropical reservoir: Dynamics and tolerance. Braz. J. Biol. 2013, 73, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Cruches, F.; Urrutia, R.; Parra, O.; Araneda, A.; Treutler, H.; Berttrand, S.; Fagel, N.; Torres, L.; Barra, R.; Chirimos, L. Change in diatom assemblages in an Andean lake in response to a recent volcanic event. Arch. Hydrobiol. 2006, 165, 23–25. [Google Scholar] [CrossRef]
- Sajekti, A.S. An Indication of Holocene Environmental Change Based on the Palynological Research in Telaga Cebong, Dieng Plateau, Central Java, Indonesia; Muséum national d’Histoire naturelle: Paris, France, 2009; pp. 1–93. [Google Scholar]
- Alles, E.; Nörpel-Schempp, M.; Lange-Bertalot, H. Taxonomy and ecology of characteristic Eunotia species in headwaters with low electric conductivity. Nova Hedwig. 1991, 53, 171–213. [Google Scholar]
- Puussepp, L.; Kangur, M. Linking diatom community dynamics to changes in terrestrial vegetation: A palaeolimnological case study of Lake Ķūži, Vidzeme Heights (Central Latvia). Est. J. Ecol. 2010, 59, 259–280. [Google Scholar] [CrossRef]
- Newhall, C.G.; Dzurisin, D. Historical Unrest at Large Calderas of the World; Technical Report 1855; Department of the Interior, US Geological Survey: Washington, DC, USA, 1988; pp. 1–1108. [Google Scholar]
- Burge, D.R.L. Relations of Water Quality, Land Use Buffers, and Diatom Communities of Connected Depressions within the Cache River Watershed, Arkansas, USA. Master’s Thesis, Arkansas State University, Jonesboro, AR, USA, 2019; pp. 1–142. [Google Scholar]
- Miller, J.R.; Mackin, G.; Orbock Miller, S.M. Application of Geochemical Tracers to Fluvial Sediment; Springer: Cham, Switzerland, 2015; pp. 1–142. [Google Scholar]
- Fan, Z.; Wang, W.; Tang, C.; Li, Y.; Wang, Z.; Lin, S.; Zeng, F. Targeting remediation dredging by ecological risk assessment of heavy metals in lake sediment: A case study of Shitang Lake, China. Sustainability 2019, 11, 7251. [Google Scholar] [CrossRef] [Green Version]
- Haryono, S.; Mulyana, A.R.; Subagyo, A. Application of SABO technique in controlling the water destructive force in dataran tinggi Dieng. J. Tek. Hidraul. 2012, 3, 157–168. [Google Scholar]
- Nugraha, S.B.; Akhsin, W.; Benardi, A.I. Pemanfaatan teknologi sig untuk pemetaan tingkat ancaman longsor di Kecamatan Kejajar, Wonosobo. J. Geogr. 2015, 12, 203–221. [Google Scholar]
- Mardiatno, D. Teknik Transplanting Tanaman Hutan; Balai Penelitian dan Pengembangan Teknologi Pemgelolaan Daerah Aliran Sungai. 2018; Volume 4, p. 2. Available online: https://www.forda-mof.org/files/Majalah_Cerdas_Vol_4_No_2_Oktober_2018.pdf (accessed on 17 March 2021).




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sari, K.; Soeprobowati, T.R.; Jumari, J.; Hariyati, R.; Miller, J.R. Trace Metals and Diatom Stratigraphy along the Sill between Lakes Telaga Warna and Telaga Pengilon, Dieng, Central Java, Indonesia. Sustainability 2021, 13, 3821. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073821
Sari K, Soeprobowati TR, Jumari J, Hariyati R, Miller JR. Trace Metals and Diatom Stratigraphy along the Sill between Lakes Telaga Warna and Telaga Pengilon, Dieng, Central Java, Indonesia. Sustainability. 2021; 13(7):3821. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073821
Chicago/Turabian StyleSari, Kenanga, Tri Retnaningsih Soeprobowati, Jumari Jumari, Riche Hariyati, and Jerry R. Miller. 2021. "Trace Metals and Diatom Stratigraphy along the Sill between Lakes Telaga Warna and Telaga Pengilon, Dieng, Central Java, Indonesia" Sustainability 13, no. 7: 3821. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073821