
The Effects of Density on the Growth and Temperature Production of Tenebrio molitor Larvae


Insect Research Centre, Inagro, Ieperseweg 87, 8800 Rumbeke-Beitem, Belgium
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(10), 6234; https://0-doi-org.brum.beds.ac.uk/10.3390/su14106234
Submission received: 4 May 2022
/
Revised: 16 May 2022
/
Accepted: 17 May 2022
/
Published: 20 May 2022
(This article belongs to the Collection Sustainable Insect Farming: Feed the Future)
Abstract
:Tenebrio molitor larvae live, at least partially, inside their feed. Hence, they do not live on a 2D plane but in a 3D environment. However, previous studies mainly focused on the optimal number of larvae for a given surface area, not the available volume. The goal of this study was to assess the growth and survival of mealworms in a standardized semi-industrial setting with a varying density (cm3) and substrate height. A full factorial experimental design was used with five larval densities (0.5–8 larvae/cm3) and four feed heights (1–8 cm) in 60 × 40 cm crates. Furthermore, the in-crate temperature was monitored and linked to the density. The results of this study clearly indicate that mealworm larvae prefer a low density (cm3). At low larvae densities, the substrate height was less important, with a slight preference for a thicker layer. In contrast, at high(er) larval densities, a lower layer thickness resulted in better growth. The in-crate week temperature varied up to 14 °C (25–39 °C) between treatments and could be predicted well based on the number and size of the larvae. These results may help the industry to improve their production efficiency in terms of larvae density, substrate height and room temperature.
1. Introduction
Conventional terrestrial livestock (e.g., cattle) live in a 2D world, such as a pasture or pigsty. The optimal stocking densities, in terms of animal welfare, growth and economic return, are based on surface area and can be expressed as the number of individuals per square meter [1,2]. In the fairly new field of insect rearing for feed and food, it may be more difficult to calculate the optimal density. Two of the economically most important insect species (Tenebrio molitor and Hermetia illucens) live, at least partially, inside their feed. Hence, they do not live on a 2D plane but in a 3D environment.
Previous studies on T. molitor focused mainly on the (optimization of the) number of larvae for the available surface area (2D). Morales-Ramos and Rojas [3] observed significant adverse effects on growth, feed consumption and efficiency with an increase in larval density (per cm2). However, in a study conducted by Deruytter and Coudron [4], the differences in growth, the feed conversion ratio and frass production between 0.6 and 10.4 larvae/cm2 were limited when the larvae were provided with an equal volume of feed. The latter indicates that the available volume is a better growth predictor compared to the available area for wheat bran. However, the study conducted by Deruytter and Coudron [4] used a single volumetric density of 1 larvae/cm3. Therefore, the optimal number of larvae per given volume is unknown. Further complicating the estimation of the optimal density is the possibility that the vertical distribution of the mealworm larvae may not be perfectly homogeneous throughout the entire available volume. Some work has been carried out with respect to the vertical distribution of stored product pest species [5], but the authors of the present study are not aware of any scientific study on this in a Tenebrio-rearing facility. Heterogeneous vertical distribution would result in different densities throughout the layers. Density differences may result in differences in growth and pupation speed, as seen in other tenebrionid beetles [6], or differences in mortality, as seen in gypsy moths [7]. Therefore, the substrate depth could affect the optimal density and should be assessed simultaneously.
As suggested by Morales-Ramos and Rojas [3], one of the potential factors that may cause a change in larval performance with a varying density is an increase in crate temperature (vs. ambient temperature), with an increase in larval activity being due to the production of metabolic heat. This change may be beneficial when the crate temperature reaches more optimal temperatures [8,9] compared to the ambient temperature or may also decrease growth when exceeding this optimum. The difference between the core crate temperature and the ambient temperature may not only depend on the number of larvae but also the size/age of the larvae and the substrate height and composition (insulation capacity).
Knowledge on the density vs. growth relationship is important for mealworm farmers trying to optimize their own production. It may also help with the translation from benchtop experiments to the industry and, finally, improve the comparability of different studies’ results. The latter is due to a lack of regulations on the number of larvae, substrate height and mandatory information in scientific publications, resulting in many different experimental designs [10,11,12]. Hence, a strong density vs. growth relationship would induce an additional variation that needs to be taken into account when comparing results.
The goal of this study was to assess the growth and survival of mealworms fed with wheat bran in a standardized semi-industrial setting with a varying density (cm3) and substrate height. Furthermore, the results were linked to the crate temperature. The results may help the industry to improve production efficiency.
2. Materials and Methods
2.1. Experimental Setup
Tenebrio molitor was reared at a temperature of 27 ± 1 °C and relative humidity (RH) of 60 ± 3% in the dark [13] at the Insect Research Centre of Inagro in 60 × 40 cm plastic crates (surface area of 2000 cm2). The larvae for the experiment were obtained by using 11 × 250 g of beetles with an average age of 4 weeks. The beetles were allowed to oviposit for 8 days. Thereafter, the eggs and feed (Insectus, Mijten bv. Belgium) were collected and placed in the same climate conditions. Two weeks after oviposition, agar gel (25 g agar/L, Brouwland bv, Belgium, [14]) was provided as wet feed ad libitum. Four weeks after oviposition, the frass was sifted out (0.5 mm), and the remaining larvae/feed mixture was gently homogenized. Three subsamples were taken to estimate the number and average weight of the larvae at the start of the experiments. This process was repeated four times to set up the total experiment in quadruplicate.
A full factorial experimental design was used with 5 larval densities and 4 feed heights in 60 × 40 × 10 cm crates (total 4 × 20 crates, Table 1). The necessary amount of wheat bran was added to each crate as dry feed (0.2 kg/L, particle size <2 mm). The original volume of wheat bran was maintained by sifting out the frass every week (in a 0.5 mm sieve), determining the residual volume of the crate and adding fresh wheat bran (0.2 kg/L, particle size <2 mm) up to the original intended volume. For the high-density crates (4 and 8 larvae/cm3), the rate at which new wheat bran was provided increased by the end of the experiment due to the voracious feeding of the larvae. Well-distributed wet feed (agar 25 g/L) was provided ad libitum. The presence of the wet feed was confirmed twice a day, and agar was added or replaced when required. To assess mealworm growth and mortality, the total weight and volume of the substrate + larvae of each crate were determined each week using an ICS425 balance (Mettler Toledo, Zaventem, Belgium) and a measuring cup of the appropriate size. After homogenizing the content, a subsample was taken to assess the number and average weight of the mealworm larvae using a ME203T (Mettler Toledo, Zaventem, Belgium, on average 150 larvae/subsample). Nine weeks after oviposition, the larvae started pupating, and the experiment was terminated. The high-density crates (8 larvae/cm3) were harvested after 4 weeks, as larvae health issues prohibited further continuation. In 3 of the 4 replicates, a temperature logger (Trix-8 LogTag®, Gennep, The Netherlands) was placed on the bottom in the center of the crate, and it recorded the temperature every 15 min for the duration of the experiment.
2.2. Data Analysis
A statistical analysis was performed using R 4.1.2 statistical software (R Foundation for Statistical Computing, Vienna, Austria). Due to the longitudinal nature of the data, taking into account the differences in initial weight of the larvae and the potential different growth curves between the replicates, linear mixed-effect modeling was necessary (Lme4 package). A correction on the theoretical density for each replicate was made based on the estimated number of mealworms in the first 2 weeks (n = 40) instead of using the initial estimate (n = 3), resulting in a more accurate density modeling. The matches between the actual and theoretical densities were 100% for R1, 88% for R2, 89% for R3 and 111% for R4.
To fully understand the relationship between density, substrate and mealworm growth, two different models were assessed. The first model describes the growth curve of the mealworm larvae over time for different densities and substrate heights (full model, Equation (1)):
where AW = log10 average weight (mg), T = time (weeks), D = log10 density (larvae/cm3) and H = log10 substrate height (cm). To determine the optimal model, the full model (Equation (1)) was reduced via backward selection until all parameters in the model were significant (p-value < 0.05).
AW = T × D × H + T2 + D2 + H2
The second model (Equation (2)) describes the changes in the potential growth rate of different mealworm sizes for the different densities and substrate heights.
where GR = potential growth rate defined as the average weight at the end of the week/average weight at the start of the week, AW = log10 average weight (mg), D = log10 density (larvae/cm3) and H = log10 substrate height (cm).
GR= AW × D × H + AW2 + D2 + H2
In order to assess the heat production in each crate, the difference between the crate temperature and the ambient temperature was calculated. The analysis only included temperature data from between 8:00 p.m. and 6:00 a.m. to eliminate any artificial temperature fluctuations that may occur during the daytime working hours, e.g., during sampling. Thereafter, the average temperature difference per week was calculated. A modeling assumption was made that the increase in temperature would mainly be due to the metabolic activity of the larvae, and potential microbial contribution was ignored; therefore, the following model was used (Equation (3)):
where TD = temperature difference between the core crate temperature and the ambient temperature (°C), AW = average weight (mg) and #larvae = estimated total number of larvae in the crate. A more simplified and practical model was also made solely based on the log10 biomass of the crate.
TD = AW × #larvae + AW2 + #larvae2
3. Results
With an average survival of 95% (SD 12%) for all assessed crates, larval mortality was limited, except for the highest density combined with the highest substrate height (Object 20, Table 1), where survival was on average 68% at the end of the experiment. As only 1 of the 20 objects had decreased survival, the statistical analysis did not result in a usable, robust, predictive model.
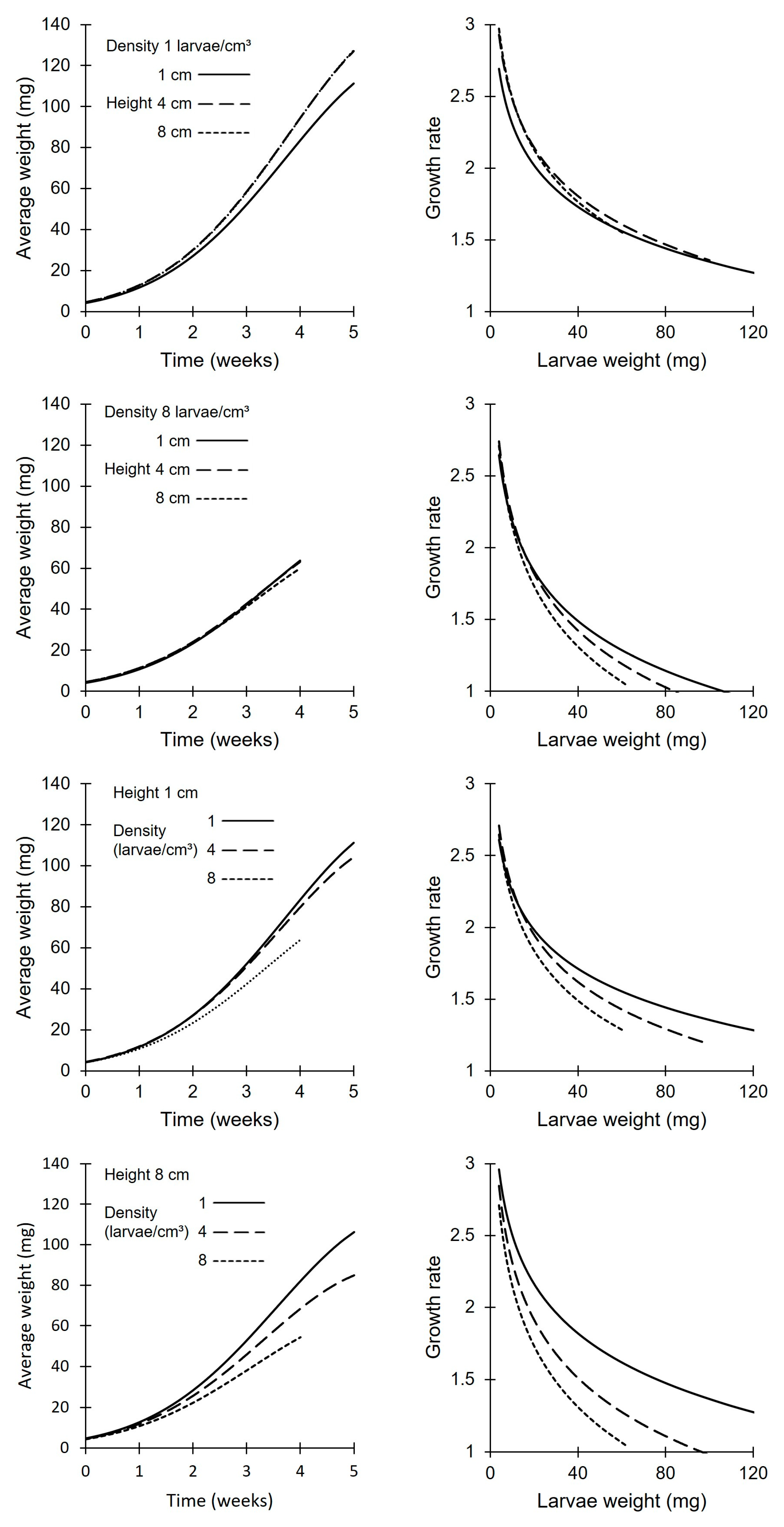
Mealworm larvae growth could be modeled well when taking into account time, larval density (per cm3) and substrate height (marginal R2: 0.94). All three parameters, interactions and quadratic effects were significant (Table 2). At a low density, larvae grew better with a high substrate height, while the opposite was observed for high densities. Irrespective of the substrate height, lower densities resulted in a faster growth with a higher average mealworm weight at harvest. The data indicate that larval density was more important than substrate height within the current experimental limits (Figure 1). Similar conclusions were reached when modeling the growth rate (per week) based on the density, substrate height and the average weight of the larvae (marginal R2: 0.91). The initial weekly growth rate started between 2.5 and 3 and declined, as expected, to 1 (no growth) near the end of the experiment. At a low density, the growth rate decreased with an increase in the larval weight, nearly irrespective of the substrate height. At higher densities, a lower substrate height resulted in a better growth rate. Conversely, the growth rate at a lower density always outperformed the growth rate at higher densities, irrespective of the size of the larvae.
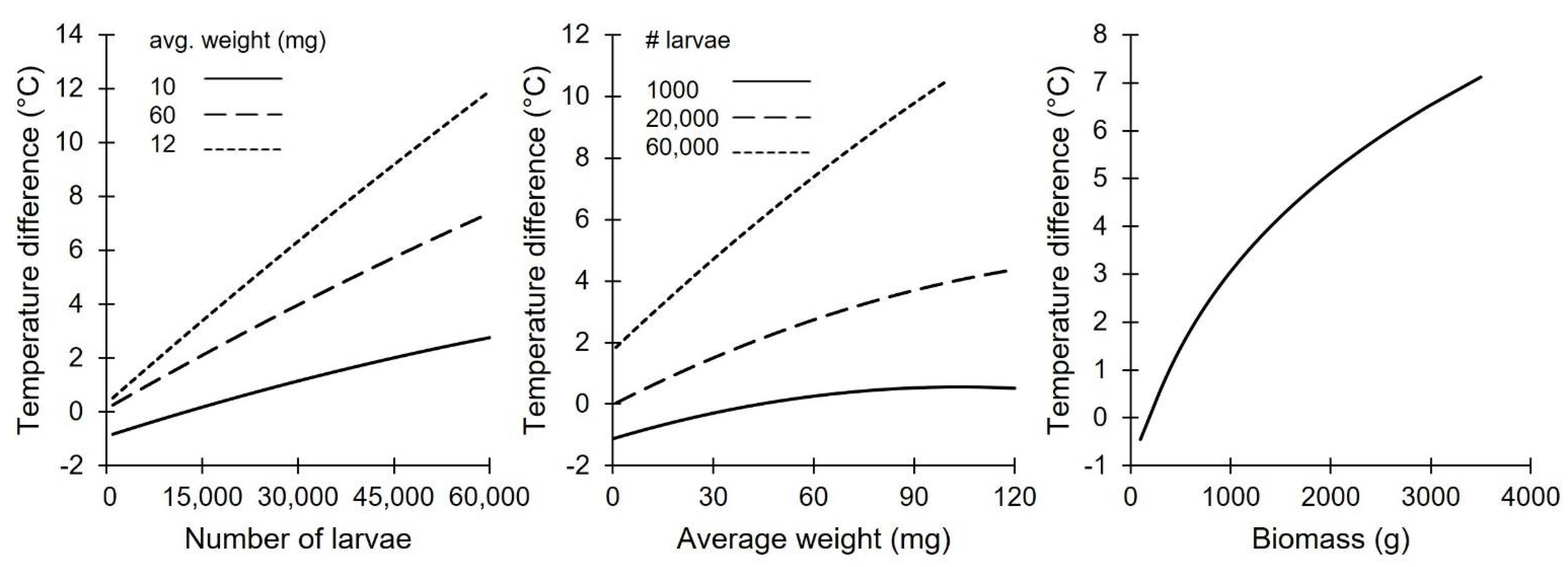
An increase in crate temperature was observed in most crates. The core temperature exceeded 41 °C at the highest density/substrate combination for all replicates, with an absolute maximum of 43.9 °C in replicate 1 for a short time. The core temperature varied between 25 °C and 39 °C between treatments. As expected, the difference between the ambient and core temperatures was significantly and positively correlated with the number of larvae and average size of the larvae (R2: 0.88, Table 3 and Figure 2). However, both the data and the model indicate that a low number of larvae combined with a low larval weight (low biomass) may result in a negative difference between the ambient and core temperatures. The model can be simplified with only biomass as an explanatory variable, as both the number of larvae and the average larval size seem to have a near identical influence on the core temperature; this results in only a 5% decrease in the explained variation (R2 of 0.83%).
4. Discussion
With a rising world population and a rising demand for protein, mealworm larvae may be a promising alternative to conventional sources of animal protein [15]. They are considered nutritious [16,17], as well as having a small environmental footprint [18]. However, there are some knowledge gaps on the basic rearing of this species, and the optimal rearing density is one of those that was addressed in this study.
4.1. Density
The results of this study clearly indicate that mealworm larvae prefer a low density in a 3D environment, with the lowest assessed density resulting in the best growth (0.5 larvae/cm3). The true optimal larvae density may be even lower. This preference is not related to the substrate height; furthermore, the absence of intersecting lines in Figure 1 indicates that the preference for a low density is not related to the weight (age) of the larvae. This implies that, within the assessed range, there are no growth benefits to be gained by, for example, housing smaller larvae more densely compared to larger larvae. However, from economic and ecological points of view, using the optimal density for growth (0.5 larvae/cm3) results in two major disadvantages when using the current setup. Firstly, in this study, the larvae had more feed (2 cm3 or 400 mg) than they could efficiently consume, resulting in increased feed costs and feed waste. Secondly, the resulting total harvest was (very) low at this density compared to that of the other treatments, resulting in a larger production footprint. To ameliorate these issues, an economically and ecologically more optimal density on wheat bran is 1 larvae/cm3 for a single feeding system. At this density, all bran can be consumed by the time the larvae are fully grown [4], and the total mealworm harvest can reach up to 2.1 kg per 60*40 crate. With multiple (or daily) feeding systems, it is possible to rear up to 4 larvae/cm3 in good health, although with a lower maximum individual weight. Combined with the highest substrate layer, 4 larvae/cm3 resulted in the highest yield in this experiment: 4.2 kg/crate (SD: 0.1 kg). Unsurprisingly, it was impossible to keep 8 larvae/cm3 healthy in the current experimental setup. Growth was slow; daily feeding was necessary at the end; and, especially with a high substrate, the temperature increased to unfavorable conditions (>40 °C) or even crossed the lethal threshold [8,19,20], resulting in excessive mortality. In contrast to all other crates, where we assumed that the temperature increased predominantly due to metabolic activity, the temperature in the highest density crates with a thick substrate layer (object 20, Table 1) was at least partially due to microbial activity. However, with the current experimental design, it was impossible to make a distinction between these two important factors.
4.2. Substrate Height
The optimal substrate layer height does depend on larval density. At low densities, the substrate height was of lesser importance with a slight preference for a thicker layer. These results concur with a previous study [4]. In that study, only limited effects were observed for different substrate heights at a larvae density of 1 larvae/cm3 with a slightly slower growth with low substrate height. This may be because low densities combined with a low substrate height do not have the biomass to produce enough metabolic heat, resulting in crate temperatures that are similar to or even lower than ambient temperature. Negative deviations may be due to the evaporation of the wet feed. These results do concur with the data of Weaver and McFarlane [21], which indicated that very low larvae numbers did not perform well. In contrast, at high(er) larval densities, a lower layer thickness was clearly preferred. It is hypothesized, again, that this is due to the heat production of the larvae and the insulation capacity of the wheat bran. This concurs with the decreased growth rate when increasing the number of larvae per square cm at higher volumetric densities (estimated between 2.4 and 5.8 larvae/cm3) observed by Morales-Ramos and Rojas [3].
4.3. Temperature
Although the crate temperature may explain some of the observed effects, it cannot explain everything. According to Bjørge et al. [8], a temperature around 31 °C would result in optimal growth for T. molitor. The current experiment was performed at an ambient temperature of 27 °C; therefore, a 4 °C increase could potentially benefit the growth of the larvae. The model (Table 3) indicates, as expected, a positive relationship between the number of larvae, the size of the larvae and the core temperature. If only the temperature would influence the growth difference, one could expect a size-dependent density preference, but this was not observed. Although the temperature models and density models seem to contradict each other, it is more nuanced. Both the size of the larvae and the number of larvae have a significant negative quadratic component resulting in a non-linear fit. This indicates that, one gram of small larvae and/or fewer larvae produce more heat than one gram of big larvae or dense crates (e.g., 4.2 °C for 100,000 larvae of 10 mg or 2.2 °C for 10,000 larvae of 100 mg). Furthermore, it is currently unknown if an age-dependent optimal growth temperature exists and what the true optimal temperature is. Bjørge et al. [8] indicated that 31 °C was the best assessed temperature; however, the other closest assessed temperatures were 25.4 or 37 degrees, still leaving a wide window to explore. Because the temperature was not actively changed but only monitored, it is impossible to determine the optimal, age-specific crate temperature with the current data. Finally, it is important to note that the results presented in this study are true for the assessed conditions: plastic 60*40 crates with wheat bran and agar as feed at 27 °C. Although commonly used equipment and environment conditions, changing these may change the outcome. For example, metal crates may dissipate the heat faster allowing for higher densities just as lowering the ambient temperature may increase the difference [9], or a different feed [22] may have a different volumetric density resulting in a different vertical distribution and thermal insulation characteristics and feed conversion. Future research could also assess if lower ambient temperatures are possible or feasible at a high-density rearing facility.
4.4. Conclusions
The results of this study indicate that larvae density and substrate layer thickness do influence the growth of larvae. Optimal growth is achieved at lower densities with a thick substrate layer. This information should be mentioned in future papers and taken into account in future (comparative) studies or reviews. The industry can use this data to optimize their production efficiency, but a balance between optimal growth an economic efficiency should be found. Future research is needed to assess if the conclusions hold true for different rearing conditions.
Author Contributions
D.D. was involved in conceptualization, data acquisition, statistical analysis and original draft preparation. C.L.C. was involved in conceptualization and reviewing the draft. J.C. was involved in reviewing the draft and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement No. 861976.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank L. De Praetere and A. Devos for the many larvae counts.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gholami, M.; Chamani, M.; Seidavi, A.; Sadeghi, A.A.; Aminafschar, M. Effects of stocking density and environmental conditions on performance, immunity, carcase characteristics, blood constitutes, and economical parameters of cobb 500 strain broiler chickens. Ital. J. Anim. Sci. 2020, 19, 524–535. [Google Scholar] [CrossRef]
- Sørensen, P.; Su, G.; Kestin, S.C. Effects of age and stocking density on leg weakness in broiler chickens. Poult. Sci. 2000, 79, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.A.; Rojas, M.G. Effect of larval density on food utilization efficiency of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2015, 108, 2259–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deruytter, D.; Coudron, C.L. The effects of density on the growth, survival and feed conversion of Tenebrio molitor larvae. J. Insects Food Feed 2022, 8, 141–146. [Google Scholar] [CrossRef]
- Anukiruthika, T.; Jian, F.; Jayas, D.S. Movement and behavioral response of stored product insects under stored grain environments-A review. J. Stored Prod. Res. 2021, 90, 101752. [Google Scholar] [CrossRef]
- Tschinkel, W.R.; Willson, C.D. Inhibition of pupation due to crowding in some tenebrionid beetles. J. Exp. Zool. 1971, 176, 137–145. [Google Scholar] [CrossRef]
- Elkinton, J.S.; Liebhold, A.M. Population dynamics of gypsy moth in North America. Annu. Rev. Entomol. 1990, 35, 571–596. [Google Scholar] [CrossRef]
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.H. Role of temperature on growth and metabolic rate in the tenebrionid beetles Alphitobius diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Coudron, C.L.; Deruytter, D.; Craeye, S.; Bleyaert, P. Entomoponics: Combining insect rearing and greenhouse vegetable production—A case study with Tenebrio molitor and high-wire cucumber cultivation. J. Insects Food Feed 2021, 8, 427–437. [Google Scholar] [CrossRef]
- Montalbán, A.; Sánchez, C.J.; Hernández, F.; Schiavone, A.; Madrid, J.; Martínez-Miró, S. Effects of Agro-Industrial Byproduct-Based Diets on the Growth Performance, Digestibility, Nutritional and Microbiota Composition of Mealworm (Tenebrio molitor L.). Insects 2022, 13, 323. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Oonincx, D.G.A.B.; Karapanagiotidis, I.T.; Vrontaki, M.; Gourgouta, M.; Asimaki, A.; Mente, E.; Athanassiou, C.G. Agricultural by-products from Greece as feed for yellow mealworm larvae: Circular economy at a local level. J. Insects Food Feed 2022, 8, 9–22. [Google Scholar] [CrossRef]
- Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Istvan Varga, Z.; Aleksza, L.; Gyuricza, C. Evaluation of organic wastes as substrates for rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus larvae as alternative feed supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Eberle, S.; Schaden, L.M.; Tintner, J.; Stauffer, C.; Schebeck, M. Effect of Temperature and Photoperiod on Development, Survival, and Growth Rate of Mealworms, Tenebrio molitor. Insects 2022, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Deruytter, D.; Coudron, C.L.; Claeys, J. The influence of wet feed distribution on the density, growth rate and growth variability of Tenebrio Molit. J. Insects Food Feed 2021, 7, 141–149. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Lee, S.M.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Flores, D.R.; Casados, L.E.; Velasco, S.F.; Ramírez, A.C.; Velázquez, G. Comparative study of composition, antioxidant and antimicrobial activity of two adult edible insects from Tenebrionidae family. BMC Chem. 2020, 14, 55. [Google Scholar] [CrossRef]
- Oonincx, D.G.; De Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans–a life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef] [Green Version]
- Punzo, F.; Mutchmor, J.A. Changes in temperature tolerance as a function of relative humidity during the larval stage of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kans. Entomol. Soc. 1978, 51, 207–212. [Google Scholar]
- Punzo, F.; Mutchmor, J.A. Effects of temperature, relative humidity and period of exposure on the survival capacity of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kans. Entomol. Soc. 1980, 53, 260–270. [Google Scholar]
- Weaver, D.K.; McFarlane, J. The effect of larval density on growth and development of Tenebrio molitor. J. Insect Physiol. 1990, 36, 531–536. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Visualization of the growth (left) and growth ratio (right) models for different densities and substrate heights. Time starts at the beginning of the experiments with mealworms of 4 weeks of age.
Figure 1.
Visualization of the growth (left) and growth ratio (right) models for different densities and substrate heights. Time starts at the beginning of the experiments with mealworms of 4 weeks of age.

Figure 2.
The modeled in-crate temperature deviation from the ambient temperature in a 60 × 40 crate using the number of larvae (left), larval weight (middle) or the total biomass (right). # = number of larvae.
Figure 2.
The modeled in-crate temperature deviation from the ambient temperature in a 60 × 40 crate using the number of larvae (left), larval weight (middle) or the total biomass (right). # = number of larvae.


{kind=link}
{kind=link}
Table 1.
The 20 different densities and feed height combinations that were assessed in this study and the corresponding volume (in a 60 × 40 × 10 crate), mass, areal density (2D) and amount of mealworms. Each object was set-up 4 times. #mw = number of mealworms.
Table 1.
The 20 different densities and feed height combinations that were assessed in this study and the corresponding volume (in a 60 × 40 × 10 crate), mass, areal density (2D) and amount of mealworms. Each object was set-up 4 times. #mw = number of mealworms.
| Object | Volumetric Density | Volume | Mass | Height | Areal Density | #mw |
|---|---|---|---|---|---|---|
| Larvae/cm3 | dm3 | Kg | cm | Larvae/cm2 | ×1000 | |
| 1 | 0.5 | 2 | 0.4 | 1 | 0.5 | 1 |
| 2 | 0.5 | 4 | 0.8 | 2 | 1 | 2 |
| 3 | 0.5 | 8 | 1.6 | 4 | 2 | 4 |
| 4 | 0.5 | 16 | 3.2 | 8 | 4 | 8 |
| 5 | 1 | 2 | 0.4 | 1 | 1 | 2 |
| 6 | 1 | 4 | 0.8 | 2 | 2 | 4 |
| 7 | 1 | 8 | 1.6 | 4 | 4 | 8 |
| 8 | 1 | 16 | 3.2 | 8 | 8 | 16 |
| 9 | 2 | 2 | 0.4 | 1 | 2 | 4 |
| 10 | 2 | 4 | 0.8 | 2 | 4 | 8 |
| 11 | 2 | 8 | 1.6 | 4 | 8 | 16 |
| 12 | 2 | 16 | 3.2 | 8 | 16 | 32 |
| 13 | 4 | 2 | 0.4 | 1 | 4 | 8 |
| 14 | 4 | 4 | 0.8 | 2 | 8 | 16 |
| 15 | 4 | 8 | 1.6 | 4 | 16 | 32 |
| 16 | 4 | 16 | 3.2 | 8 | 32 | 64 |
| 17 | 8 | 2 | 0.4 | 1 | 8 | 16 |
| 18 | 8 | 4 | 0.8 | 2 | 16 | 32 |
| 19 | 8 | 8 | 1.6 | 4 | 32 | 64 |
| 20 | 8 | 16 | 3.2 | 8 | 64 | 128 |
Table 2.
The mealworm larvae growth and growth ratio models with Int. = intercept, T = time (w), H = substrate height, D = larvae density (cm3), W = average weight larvae and SE = standard error.
Table 2.
The mealworm larvae growth and growth ratio models with Int. = intercept, T = time (w), H = substrate height, D = larvae density (cm3), W = average weight larvae and SE = standard error.
| Growth Model | Growth Ratio Model | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Estimate | SE | p-Value | Parameter | Estimate | SE | p-Value |
| Int. | 0.630 | 0.0104 | <0.001 | Int | 3.27 | 0.0623 | <0.001 |
| T | 0.481 | 6.20 × 10−4 | <0.001 | W | −0.963 | 0.0386 | <0.001 |
| D | 0.0807 | 0.0137 | <0.001 | D | 0.316 | 0.0753 | <0.001 |
| H | 0.122 | 0.0193 | <0.001 | H | 0.730 | 0.120 | <0.001 |
| T2 | −0.0396 | 1.32 × 10−5 | <0.001 | ||||
| H2 | −0.102 | 0.0179 | <0.001 | H2 | −0.287 | 0.094 | 0.0024 |
| D2 | −0.107 | 0.0106 | <0.001 | D2 | −0.267 | 0.0542 | <0.001 |
| T × D | −0.0282 | 5.60 × 10−4 | <0.001 | W × D | −0.212 | 0.0440 | <0.001 |
| T × H | 7.09 × 10−3 | 4.67 × 10−4 | 0.04 | W × H | −0.269 | 0.0573 | <0.001 |
| H × D | 2.18 × 10−3 | 0.0217 | 0.92 | D × H | −0.263 | 0.0596 | <0.001 |
| T × D × H | −0.0380 | 9.95 × 10−4 | <0.001 | ||||
Table 3.
The larval temperature model where W is the average weight of the larvae and # larvae is the estimated number of larvae, and the Biomass temperature model where B is the total biomass in a crate, # = number of larvae.
Table 3.
The larval temperature model where W is the average weight of the larvae and # larvae is the estimated number of larvae, and the Biomass temperature model where B is the total biomass in a crate, # = number of larvae.
| Larvae Temperature | Biomass Temperature | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Estimate | SE | p-Value | Parameter | Estimate | SE | p-Value |
| Int. | −1.21 | 0.209 | <0.001 | Int. | 7.95 | 1.04 | <0.001 |
| W | 0.0316 | 6.36 × 10−3 | <0.001 | B | −9.34 | 0.828 | <0.001 |
| #larvae | 6.57 × 10−5 | 8.11 × 10−6 | <0.001 | B2 | 2.57 | 0.16 | <0.001 |
| #larvae2 | −2.61 × 10−10 | 4.97 × 10−11 | <0.001 | ||||
| W2 | −1.57 × 10−4 | 4.15 × 10−5 | <0.001 | ||||
| W × #larvae | 1.18 × 10−6 | 9.50 × 10−8 | <0.001 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deruytter, D.; Coudron, C.L.; Claeys, J. The Effects of Density on the Growth and Temperature Production of Tenebrio molitor Larvae. Sustainability 2022, 14, 6234. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106234
AMA Style
Deruytter D, Coudron CL, Claeys J. The Effects of Density on the Growth and Temperature Production of Tenebrio molitor Larvae. Sustainability. 2022; 14(10):6234. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106234
Chicago/Turabian StyleDeruytter, David, Carl L. Coudron, and Jonas Claeys. 2022. "The Effects of Density on the Growth and Temperature Production of Tenebrio molitor Larvae" Sustainability 14, no. 10: 6234. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106234
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.