
Changes in the Platycodin Content and Physiological Characteristics during the Fruiting Stage of Platycodon grandiflorum under Drought Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
- (1)
- Prior to withholding water (0 d): The drought group was saturated with water at one time; the soil water content was about 33%. The CK was under normal management; water was supplied by simulated artificial precipitation between 5:00 p.m. and 6:00 p.m. every 2 days to ensure the normal growth and development of PG. The water supply scheme is shown in Table 1.
- (2)
- After withholding water (AW): The drought group was no longer watered after the drought stress experiment began until two-thirds of the leaves of PG had withered.
- (3)
- After rewatering (AR): When two-thirds of the PG leaves had withered, water was supplied to the drought group again between 5:00 p.m. and 6:00 p.m. that day.
- (4)
- Autumn period (AP): The water resupply in the drought group was managed the same as that in CK until a unified harvest on 20 October 2019.
2.3. Experimental Treatment
2.4. Determination of Fresh and Dry Root Weight
2.5. Determination of Photosynthesis Parameters
2.6. Determination of Chlorophyll Fluorescence Parameters
2.7. Contents of Osmoregulatory Substances and Malondialdehyde
2.8. Antioxidant Enzyme Activities
2.9. Measurement of Platycodin Content
2.10. Measurement of the Total Platycodin Content
2.11. Statistical Analysis
3. Results
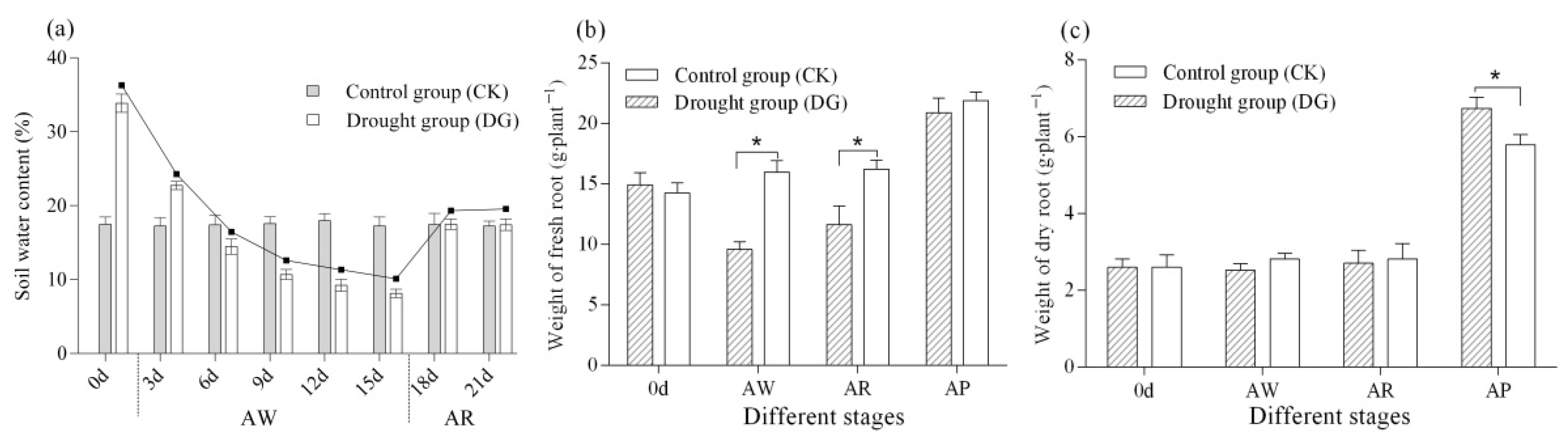
3.1. Soil Water Content
3.2. Root Growth
3.3. Physiological and Biochemical Indexes
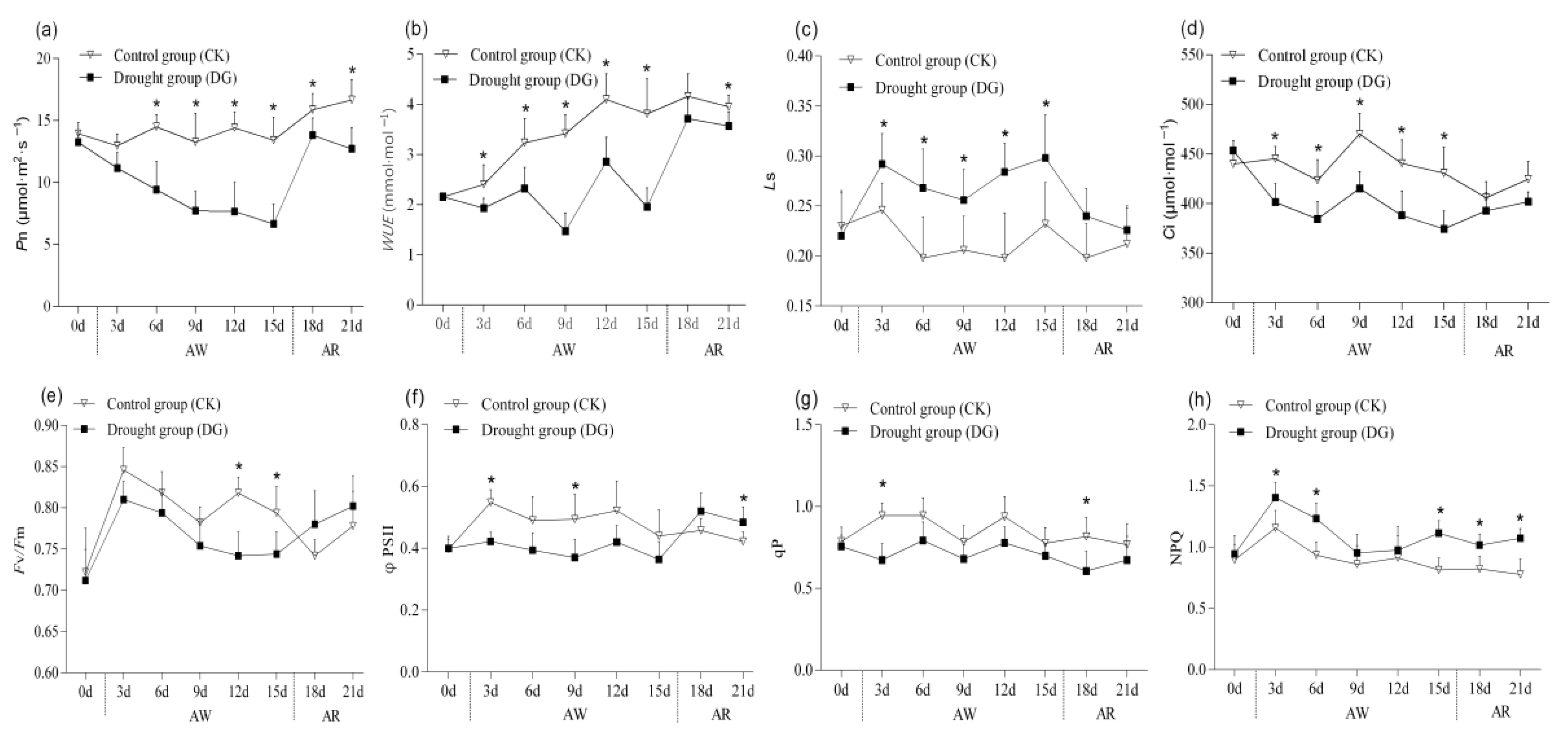
3.3.1. Photosynthesis
3.3.2. Chlorophyll Fluorescent Parameters
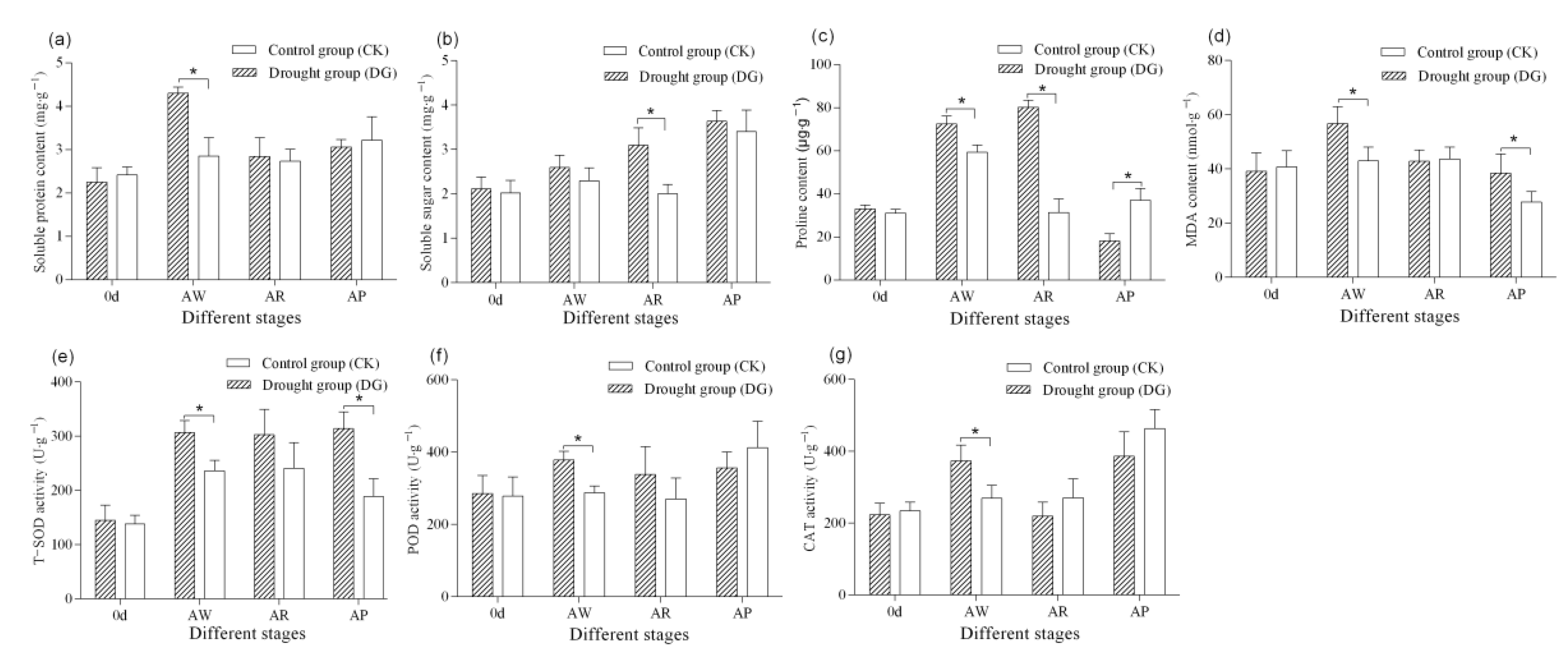
3.3.3. Osmotic Substances and Malondialdehyde
3.3.4. Protective Enzyme
3.4. Platycodins
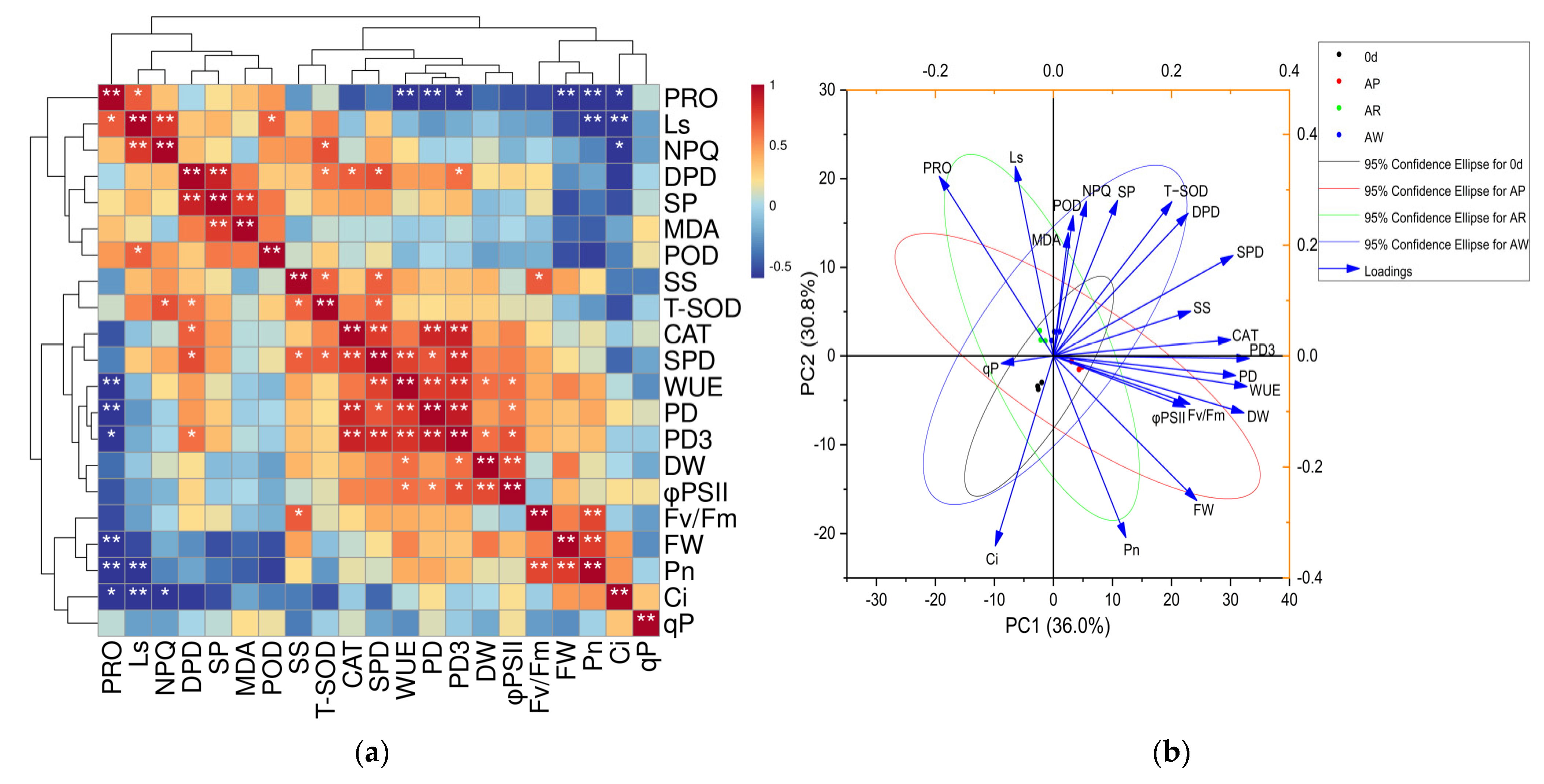
3.5. Principal Component Analysis and Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- China National Pharmacopoeia Commission. First Pharmacopoeia of China; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]
- Cuizhu, W.; Nanqi, Z.; Zhenzhou, W.; Zeng, Q.; Hailin, Z.; Bingzhen, Z. Nontargeted metabolomic analysis of four different parts of Platycodon grandiflorum grown in northeast China. Molecules 2017, 22, 1280. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Song, I.B.; Lee, H.K.; Lim, J.H.; Cho, E.S.; Son, H.Y. Platycodin D, a triterpenoid sapoinin from Platycodon grandiflorum, ameliorates cisplatin-induced nephrotoxicity in mice. Food Chem. Toxicol. 2012, 50, 4254–4259. [Google Scholar] [CrossRef] [PubMed]
- Ihuoma, S.O.; Madramootoo, C.A. Recent advances in crop water stress detection. Comput. Electron. Agric. 2017, 141, 267–275. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. C. R. Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Olga, B.; Eija, V.; Kurt, V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Hassan, N.; El-Bastawisy, Z.; Ebeed, H.; Nemat Alla, M. Role of defense enzymes, proteins, solutes and Δ1-pyrroline-5-carboxylate synthase in wheat tolerance to drought. Rend. Lincei 2015, 26, 281–291. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Xiuli, Z. Study on Effect of Drought Stress on Synthesis of Pharmacodynamic Components of Atracylodes chinensis and Its Regulation Mechanism. Master’s Thesis, Changchun University of Traditional Chinese Medicine, Changchun, China, 2020. [Google Scholar] [CrossRef]
- Sgherri, C.; Kadlecová, Z.; Pardossi, A.; Navari-Izzo, F.; Izzo, R. Irrigation with diluted seawater improves the nutritional value of cherry tomatoes. J. Agric. Food Chem. 2008, 56, 3391–3397. [Google Scholar] [CrossRef]
- Hassan, M.A.L.; Fuertes, M.M.; Sanchez, F.J.R.; Vicente, O.; Boscaiu, M. Effects of salt and water stress on plant growth and on accumulation of osmolytes and antioxidant compounds in cherry tomato. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 1–11. [Google Scholar] [CrossRef]
- Ghorbanli, M.; Gafarabad, M.; Amirkian, T.; Mamaghani, B.A. Investigation on proline, total protein, chlorophyll ascorbate and dehydroascorbate changes under drought stress in Akria and Mobil tomato cultivars. Iran. J. Plant Physiol. 2013, 3, 651–658. [Google Scholar]
- Meena, Y.K.; Kaur, N. Towards an Understanding of Physiological and Biochemical Mechanisms of Drought Tolerance in Plant. Annu. Res. Rev. Biol. 2019, 31, 1–13. [Google Scholar] [CrossRef]
- Weiwei, D.; Xxiaohong, Y.; Hongwen, H. Effect of environmental stress on tripterine of Tripterygium wilfordii. J. Plant Ecol. 2009, 33, 180–185. [Google Scholar]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Limin, Y.; Yonggang, Z.; Hongmei, L.; Zhongming, H.; Li, Y. Progress in quality formation theory and control technology of Chinese medicinal materials. J. Jilin Agric. Univ. 2012, 34, 119–124. [Google Scholar] [CrossRef]
- Lin, C.; Mei, H.; Limin, Y.; Li, Y.; Zhuo, S.; Tao, Z. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crops Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Wang, F.; Ma, Y.; Jiao, G.; Li, S.; Qiu, X. Effect of water stress on the growth and development of Platycodon grandiflorum. J. Northwest Norm. Univ. (Nat. Sci. Ed.) 2010, 46, 82–85. [Google Scholar]
- Chen, X.; Wang, L.; Mao, Y.; Yang, C. Plant terpenoid biosynthesis and insect resistance response. Life Sci. 2015, 27, 813–818. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Freeman, J.; Grene, R.; Tokuhisa, J. A rootstock provides water conservation for a grafted commercial tomato (Solanum lycopersicum L.) line in response to mild-drought conditions: A focus on vegetative growth and photosynthetic parameters. PLoS ONE 2014, 9, e115380. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, M.; Jiang, X.; Zhao, S.; Yang, L. Physiological and ecological response of Scutellaria baicalensis to drought rehydration. China J. Tradit. Chin. Med. Pharm. 2013, 38, 3845–3850. [Google Scholar]
- Tan, L.; Hou, X.; Hu, Z. Determination of total saponin and saponin D in different origin. Chin. Herb. Med. 2015, 46, 1682–1684. [Google Scholar]
- Shen, S.; Wu, X.; Zheng, Y. Advances in studying drought stress response mechanisms in plants-from phenotype to molecular. Biotechnol. Prog. 2017, 7, 169–176. [Google Scholar] [CrossRef]
- Centritto, M.; Brilli, F.; Fodale, R.; Loreto, F. Different sensitivity of isoprene emission, respiration and photosynthesis to high growth temperature coupled with drought stress in black poplar (Populus nigra) saplings. Tree Physiol. 2011, 31, 275–286. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Sheng, M.; Tang, M. Effects of Rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in Robinia pseudoacacia L. under drought stress. Front. Plant Sci. 2017, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of oxygen evolution and chlorophyll fluorescence characteristics at 77_K among vascular plants of diverse origin. Plant 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Functional Study of VyP5CR, a Drought Resistance Related Gene in Wild Yanshan Grape in China. Ph.D. Thesis, Northwest A&F University, Shanxi, China, 2020. [Google Scholar] [CrossRef]
- Jiang, S.; Zhu, Q.Z.; Zhang, Z.H. Effects of chitosan on wheat germination and activities of protective enzyme in seedlings under drought stess. Agric. Res. Arid. Areas 2011, 29, 206–209. [Google Scholar]
- Zhang, Z.M.; Dai, L.X.; Song, W.-W.; Hong, D.; Ci, D.-W.; Kang, T.; Ning, T.-Y.; Wan, S.-B. Effect of drought stresses at different growth stages on peanut leaf protective enzyme activities and osmoregulation substances content. Acta Agron. Sin. 2013, 39, 133–141. [Google Scholar] [CrossRef]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Sa-Nunes, A.; Faccioli, L.H. The activity of medicinal plants and secondary metabolites on eosinophilic inflammation. Pharmacol. Res. 2010, 62, 298–307. [Google Scholar] [CrossRef]
- Li, M.; Zhang, M.; Wang, S.; Cheng, L.; Han, M.; Yang, L. Effect of short-term drought stress on Platycodon grandiflorum physiological ecology and its saponin content. Chin. Pat. Med. 2021, 43, 2910–2916. [Google Scholar]
- Jia, X.; Sun, C.; Li, G.; Li, G.; Chen, G. Effects of progressive drought stress on the physiology, antioxidative enzymes and secondary metabolites of Radix Astragali. Acta Physiol. Plant. 2015, 37, 262. [Google Scholar] [CrossRef]
- Yadav, R.K.; Sangwan, R.S.; Sabir, F.; Srivastava, A.K.; Sangwan, N.S. Effect of prolonged water stress on specialized secondary metabolites, peltate glandular trichomes, and pathway gene expression in Artemisia annua L. Plant Physiol. Biochem. 2014, 74, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Zahir, A.; Abbasi, B.H.; Adil, M.; Anjum, S.; Zia, M. Synergistic effects of drought stress and photoperiods on phenology and secondary metabolism of Silybum marianum. Appl. Biochem. Biotechnol. 2014, 174, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, M.; Rosenthal, G.A. Herbivores: Their Interactions with Secondary Plant Metabolites; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Harborne, J.B. Introduction to Ecological Biochemistry; Academic Press: London, UK, 2014. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Prior to Withholding Water (0 d) | After Withholding Water (AW) | After Rewatering (AR) | Autumn Period (AP) |
|---|---|---|---|---|
| (4 September 2019) (1 Day) | (4 September 2019–19 September 2019) (15 Days) | (19 September 2019–30 September 2019) (12 Days) | (1 October 2019–20 October 2019) (20 Days) | |
| Control group (CK) | 176 mL | 176 mL | 176 mL | 100 mL |
| Drought group (DG) | The saturated soil water content (33%) | — | 176 mL | 100 mL |
| Collection Time | 5 September 2019 | 19 September 2019 | 22 September 2019 | 20 October 2019 |
|---|---|---|---|---|
| Control group (CK) | (0 d) | Day 15 (Day 15 after withholding water) | Day 18 (Day 3 after rewatering) | Autumn harvest period of Platycodon grandiflorum medicinal materials |
| Drought group (DG) |
| Standard | Regression Equation | R2 | Linear Range (µg·mL−1) |
|---|---|---|---|
| Platycodin D (PD) | logY = 0.6115 logX − 3.3246 | 0.9999 | 5~200 |
| deapioplatycodin (DPD) | logY = 0.6004 logX − 3.3411 | 0.9999 | 5~200 |
| Platycodin D3 (PD3) | logY = 0.5969 logX − 3.2691 | 0.9998 | 5~200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Zhang, M.; Cheng, L.; Yang, L.; Han, M. Changes in the Platycodin Content and Physiological Characteristics during the Fruiting Stage of Platycodon grandiflorum under Drought Stress. Sustainability 2022, 14, 6285. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106285
Li M, Zhang M, Cheng L, Yang L, Han M. Changes in the Platycodin Content and Physiological Characteristics during the Fruiting Stage of Platycodon grandiflorum under Drought Stress. Sustainability. 2022; 14(10):6285. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106285
Chicago/Turabian StyleLi, Min, Meng Zhang, Lin Cheng, Limin Yang, and Mei Han. 2022. "Changes in the Platycodin Content and Physiological Characteristics during the Fruiting Stage of Platycodon grandiflorum under Drought Stress" Sustainability 14, no. 10: 6285. https://0-doi-org.brum.beds.ac.uk/10.3390/su14106285