
Moving towards a Competitive Fully Enzymatic Biodiesel Process

Abstract
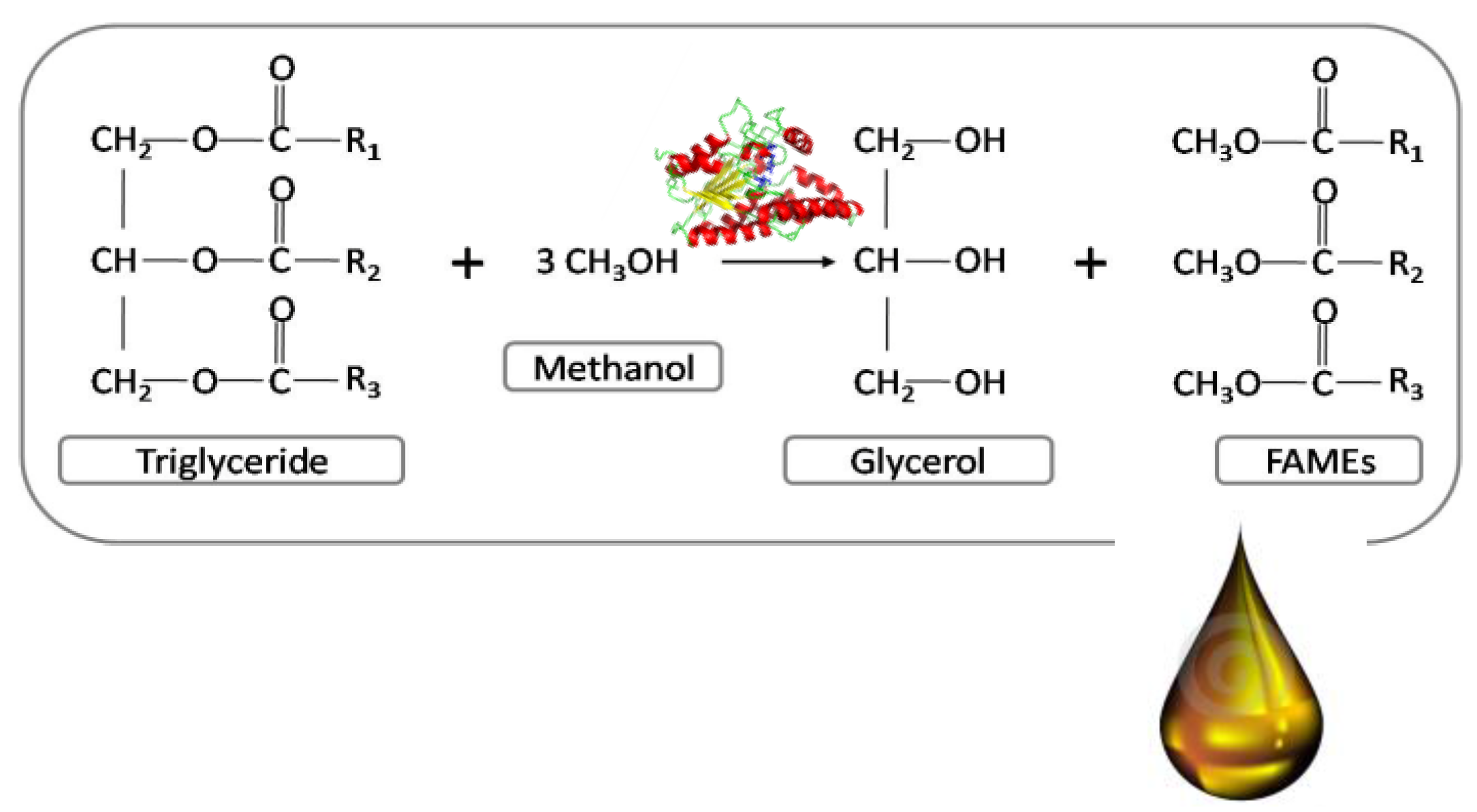
:1. Introduction

2. The Enzymes
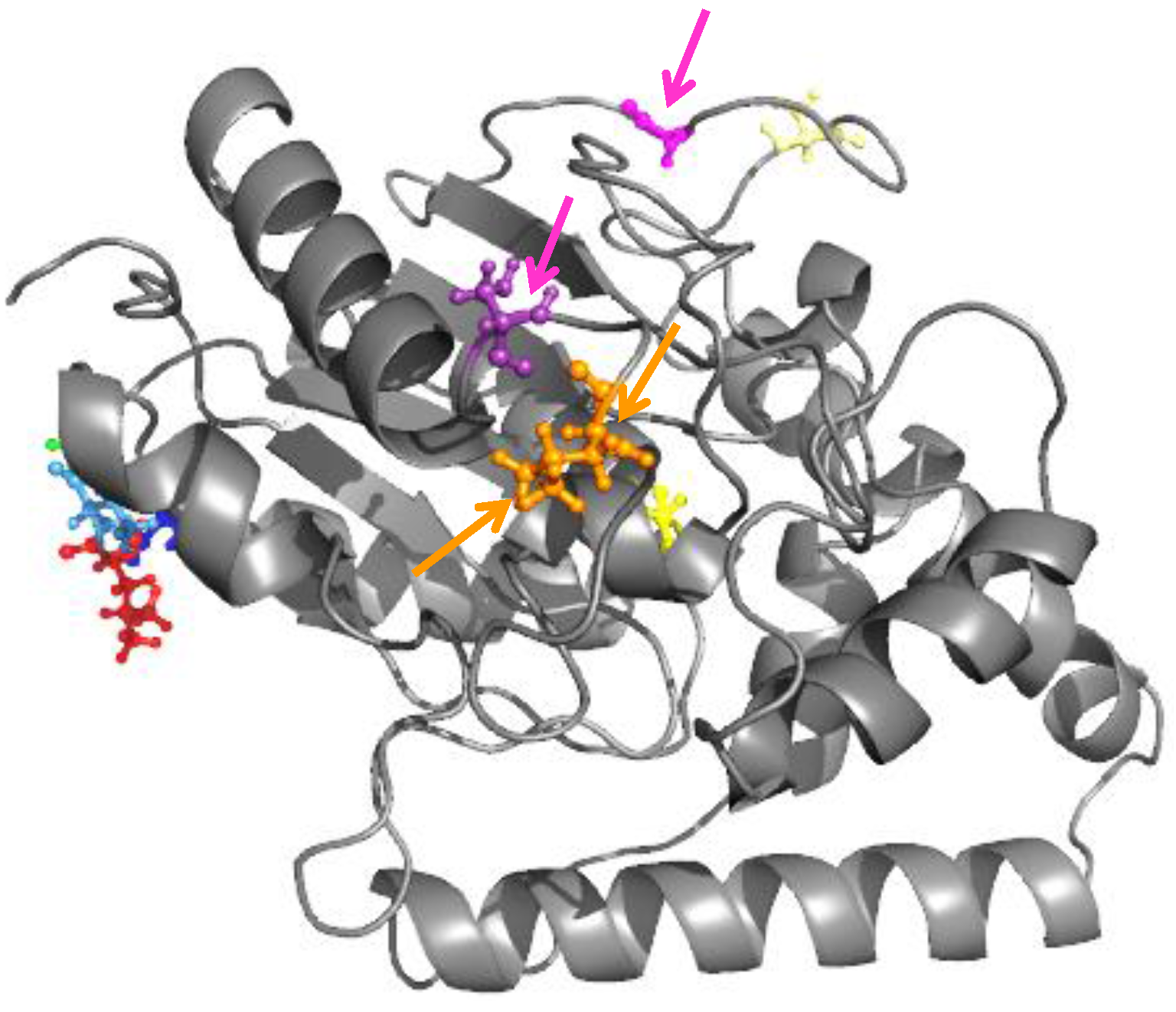
2.1. Improvement of LipC Thermoresistance

2.2. Lipase Immobilization
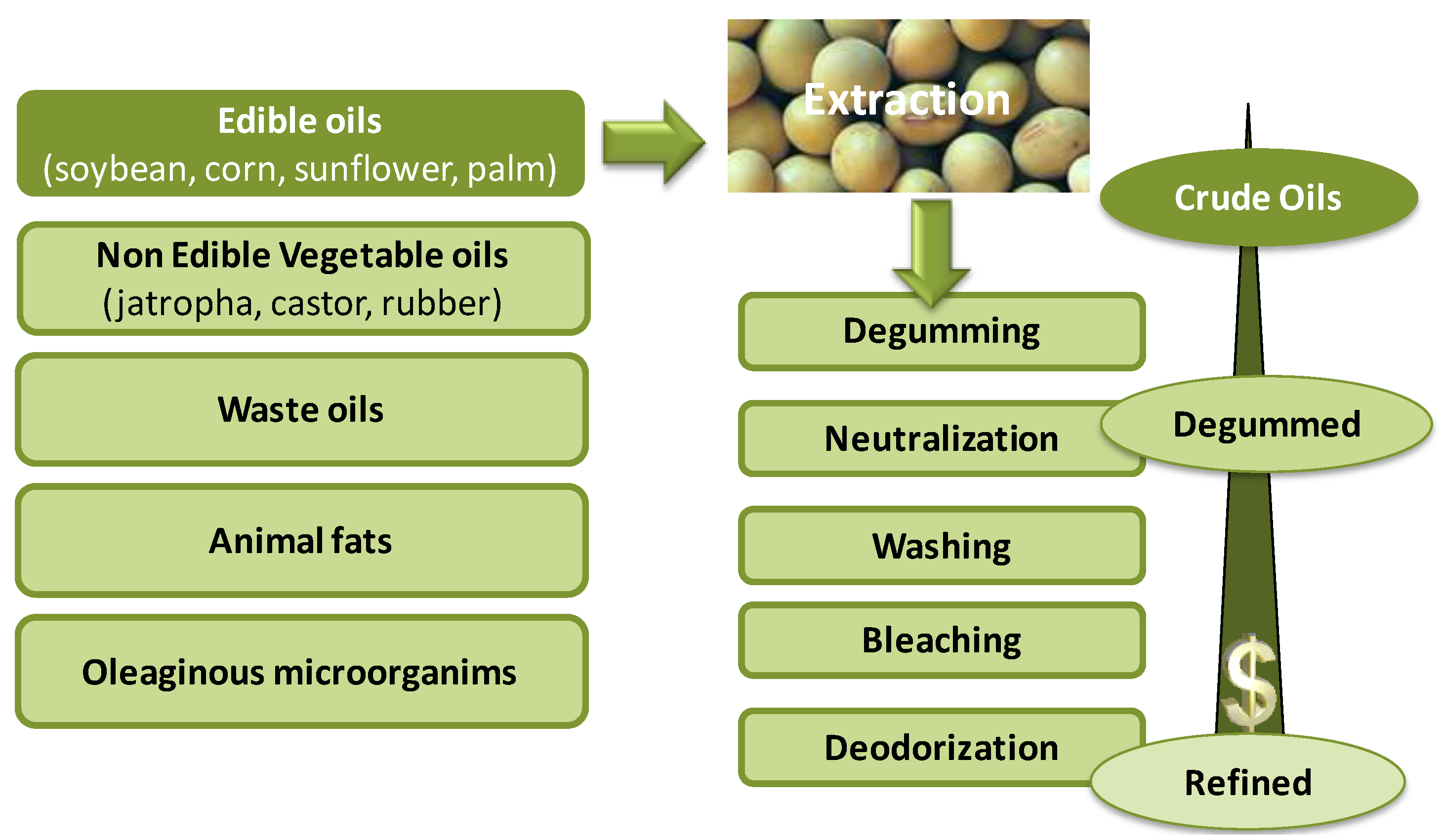
3. The Feedstocks

4. Lipase-Catalyzed Biodiesel Synthesis
4.1. Evaluation of Soluble and Immobilized Lipases
4.2. Testing Soluble Lipase Callera Trans L

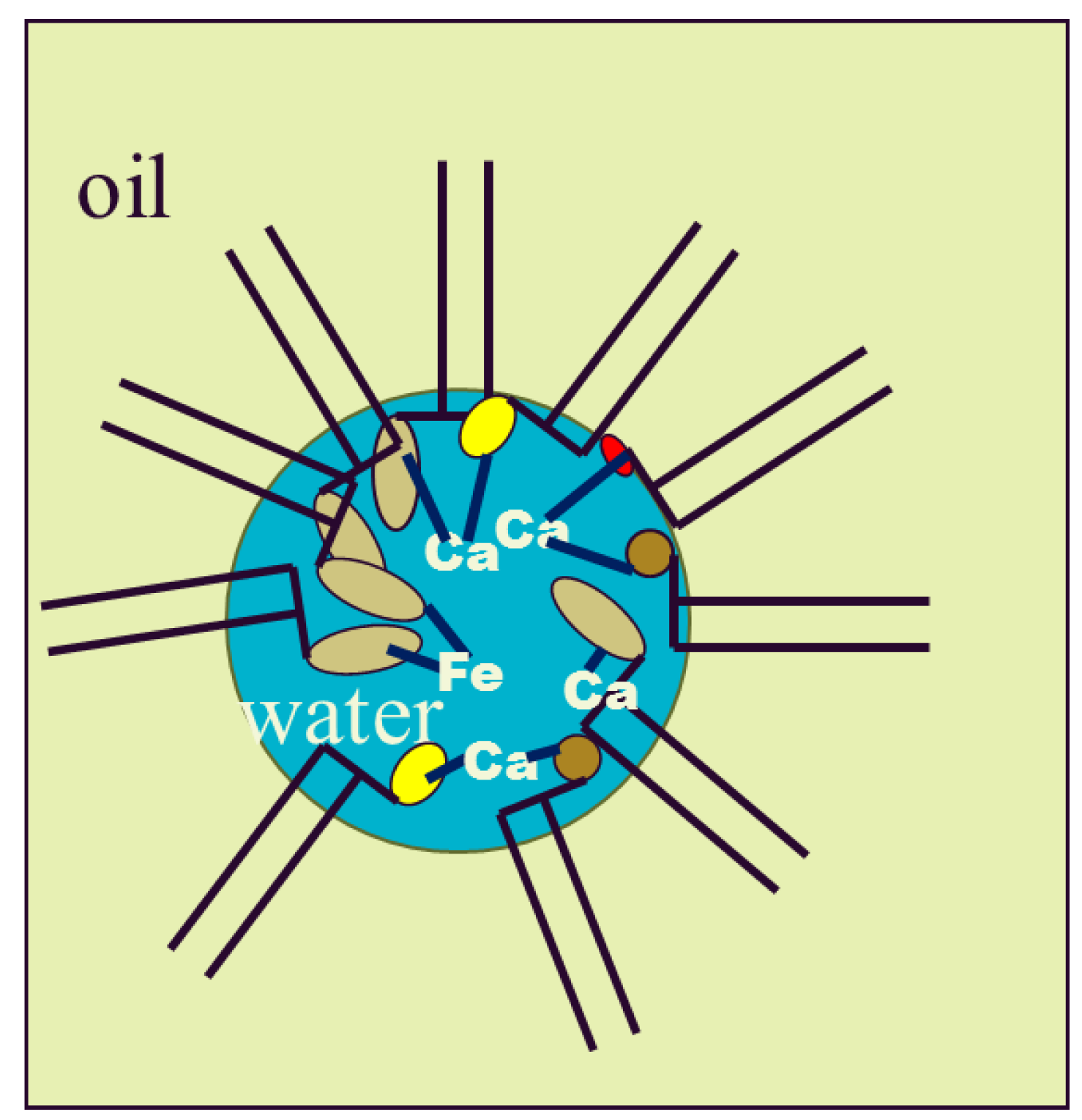
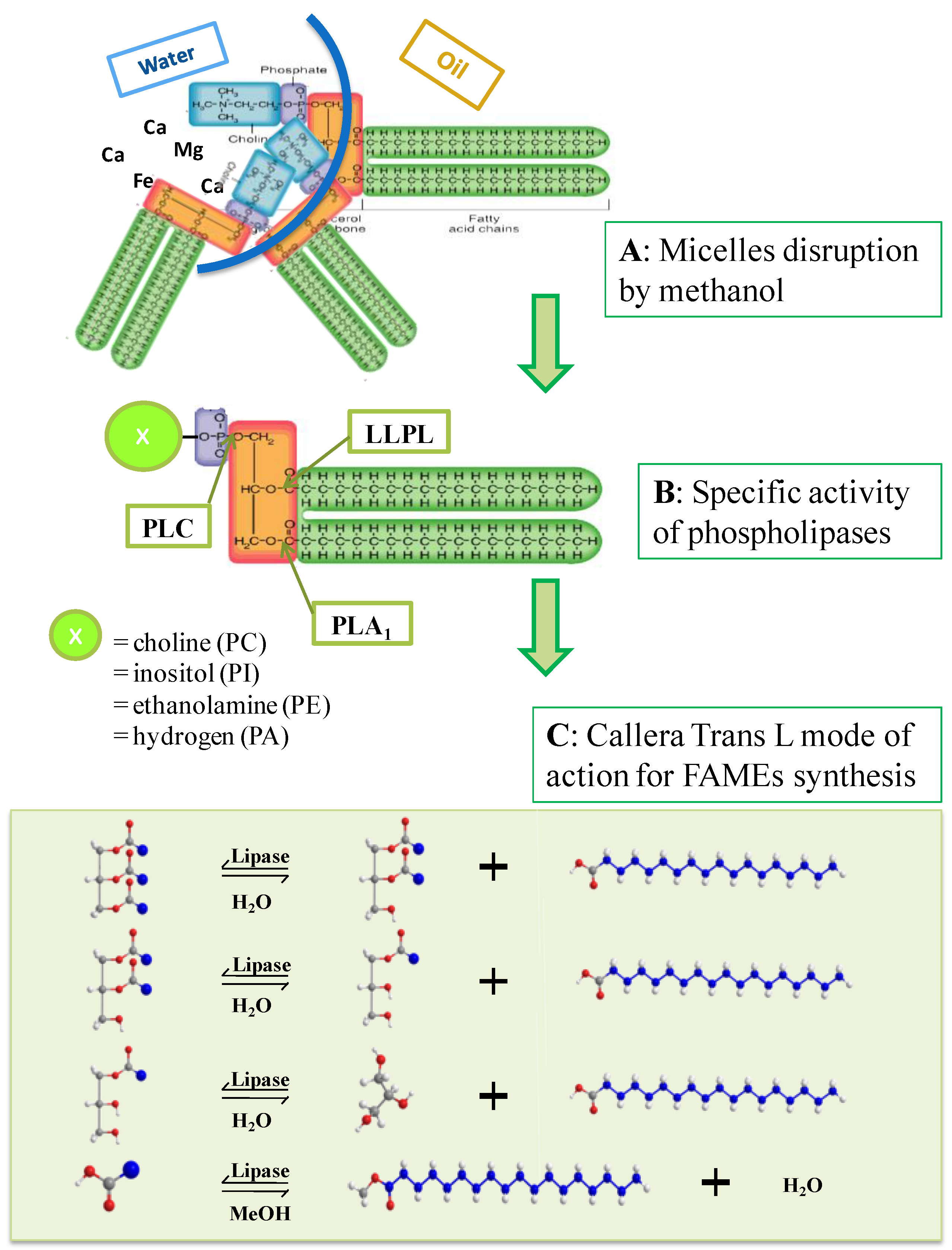
5. Degumming

5.1. Combined Enzymatic Degumming and Transesterification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soybean Oil (1% FFA; 900 ppm P) | Corn Oil (6% FFA; 62 ppm P) | Canola Oil (1% FFA; 250 ppm P) | ||||
|---|---|---|---|---|---|---|
| Enzyme Combination * | FAMEs | P | FAMEs | P | FAMEs | P |
| Callera | 85.2 | 823 | 93.4 | 7.5 | 98.7 | 145 |
| PLA1 + Callera | 98.2 | 8.0 | 94.5 | 7.3 | 98.3 | 142 |
| PLC + Callera | 90.8 | 12.8 | 95.7 | 9.2 | 98.2 | 130 |
| PLA1 + LLPL-2 + Callera | 97.8 | 6.0 | 96.2 | 9.3 | 98.9 | 145 |
| PLA1 + PLC + Callera | 96.6 | 4.6 | 95.5 | 7.5 | 99.1 | 130 |

5.2. Other Feedstocks Evaluation
5.3. Phospholipase Activity in the Combined Process
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Al-Zuhair, S. Production of biodiesel: Possibilities and challenges. Biofuels Bioprod. Biorefin. 2007, 1, 57–66. [Google Scholar] [CrossRef]
- Fjerbaek, L.; Christensen, K.V.; Norddahl, B. A review of the current state of biodiesel production using enzymatic transesterification. Biotechnol. Bioeng. 2009, 102, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Lampert, D. Monitoring chemical interesterification. J. Am. Oil Chem. Soc. 1999, 76, 783–787. [Google Scholar] [CrossRef]
- Shimada, Y.; Watanabe, Y.; Sugihara, A.; Tominaga, Y. Enzymatic alcoholysis for biodiesel fuel production and application of the reaction to oil processing. J. Mol. Catalysis B 2002, 17, 133–142. [Google Scholar] [CrossRef]
- Marchetti, M.; Miguela, V.U.; Errazu, A.F. Possible methods for biodiesel production. Renew. Sustain. Energy Rev. 2007, 11, 1300–1311. [Google Scholar] [CrossRef]
- Marchetti, J.M.; Miguel, V.U.; Errazu, A.F. Techno-economic study of different alternatives for biodiesel production. Fuel Process. Technol. 2008, 89, 740–748. [Google Scholar] [CrossRef]
- Ranganathan, S.V.; Narasimhan, S.L.; Muthukumar, K. An overview of enzymatic production of biodiesel. Bioresour. Technol. 2008, 99, 3975–3981. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, A.E.; Dave, D.; Brooks, M.S.; Budge, S. Production of biodiesel by enzymatic transesterification: Review. Am. J. Biochem. Biotechnol. 2010, 6, 54–76. [Google Scholar] [CrossRef]
- Zhao, X.; Qi, F.; Yuan, C.; Du, W.; Liu, D. Lipase-catalyzed process for biodiesel production: Enzyme immobilization, process simulation and optimization. Renew. Sustain. Energy Rev. 2015, 44, 182–197. [Google Scholar] [CrossRef]
- Szczesna Antczak, M.; Kubiak, A.; Antczak, T.; Bielecki, S. Enzymatic biodiesel synthesis—Key factors affecting efficiency of the process. Renew. Energy 2009, 34, 1185–1194. [Google Scholar]
- Dizge, N.; Aydiner, C.; Imer, D.Y.; Bayramoglu, M.; Tanriseven, A.; Keskinler, B. Biodiesel production from sunflower, soybean, and waste cooking oils by transesterification using lipase immobilized onto a novel microporous polymer. Bioresour. Technol. 2009, 100, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Vyas, A.P.; Verma, J.L.; Subrahmanyam, N. A review on fame production processes. Fuel 2010, 89, 1–9. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Bessler, C.; Srinivas, R.; Krishna, S.H. Optimizing lipases and related enzymes for efficient application. Trends Biotechnol. 2002, 20, 433–437. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Eggert, T. Lipases for biotechnology. Curr. Opin. Biotechnol. 2002, 13, 390–397. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Dijkstra, B.W.; Reetz, M.T. Bacterial biocatalysts: Molecular biology, three-dimensional structures, and biotechnological applications of lipases. Annu. Rev. Microbiol. 1999, 53, 315–351. [Google Scholar] [CrossRef] [PubMed]
- Klibanov, A.M. Asymmetric transformations catalyzed by enzymes in organic-solvents. Acc. Chem. Res. 1990, 23, 114–120. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Reetz, M.T. Directed evolution as a means to create enantioselective enzymes. Abstr. Pap. Am. Chem. Soc. 2002. [Google Scholar] [CrossRef]
- Fillat, A.; Romea, P.; Urpi, F.; Javier Pastor, F.I.; Diaz, P. Improving enantioselectivity towards tertiary alcohols using mutants of Bacillus sp bp-7 esterase estbp7 holding a rare ggg(x)-oxyanion hole. Appl. Microbiol. Biotechnol. 2014, 98, 4479–4490. [Google Scholar] [CrossRef] [PubMed]
- Bassegoda, A.; Fillat, A.; Pastor, F.I.J.; Diaz, P. Special rhodococcus sp cr-53 esterase est4 contains a ggg(a)x-oxyanion hole conferring activity for the kinetic resolution of tertiary alcohols. Appl. Microbiol. Biotechnol. 2013, 97, 8559–8568. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.S.; Vieira, N.S.; Cunha, A.S.; Silva, A.M.; Zachia Ayub, M.A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Combi-lipase for heterogeneous substrates: A new approach for hydrolysis of soybean oil using mixtures of biocatalysts. RSC Adv. 2014, 4, 6863–6868. [Google Scholar] [CrossRef]
- Poppe, J.K.; Matte, C.R.; Ruaro Peralba, M.D.C.; Fernandez-Lafuente, R.; Rodrigues, R.C.; Zachia Ayub, M.A. Optimization of ethyl ester production from olive and palm oils using mixtures of immobilized lipases. Appl. Catal. A 2015, 490, 50–56. [Google Scholar] [CrossRef]
- Ruiz, C.; Pastor, F.I.J.; Diaz, P. Isolation of lipid- and polysaccharide-degrading micro-organisms from subtropical forest soil, and analysis of lipolytic strain bacillus sp cr-179. Lett. Appl. Microbiol. 2005, 40, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Bassegoda, A.; Pastor, F.I.J.; Diaz, P. Rhodococcus sp strain cr-53 lipr, the first member of a new bacterial lipase family (family x) displaying an unusual y-type oxyanion hole, similar to the Candida antarctica lipase clan. Appl. Environ. Microbiol. 2012, 78, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Bommarius, A.; Bommarius-Riebel, B. Fundamentals of Biocatalysis; Wiley-VCH: Weinheim, Germany, 2005. [Google Scholar]
- Lutz, S.; Patrick, W.M. Novel methods for directed evolution of enzymes: Quality, not quantity. Curr. Opin. Biotechnol. 2004, 15, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Bornscheuer, U.T.; Pohl, M. Improved biocatalysts by directed evolution and rational protein design. Curr. Opin. Chem. Biol. 2001, 5, 137–143. [Google Scholar] [CrossRef]
- Arnold, G. Methods in molecular biology. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2003. [Google Scholar]
- Reetz, M.T.; Hobenreich, H.; Soni, P.; Fernandez, L. A genetic selection system for evolving enantioselectivity of enzymes. Chem. Commun. 2008, 5502–5504. [Google Scholar] [CrossRef] [PubMed]
- Bassegoda, A.; Cesarini, S.; Diaz, P. Lipase improvement: Goals and strategies: Minireview. Comput. Struct. Biotechnol. J. 2012, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bofill, C.; Prim, N.; Mormeneo, M.; Manresa, A.; Javier Pastor, F.I.; Diaz, P. Differential behaviour of pseudomonas sp 42a2 lipc, a lipase showing greater versatility than its counterpart lipa. Biochimie 2010, 92, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Reetz, M.T.; Carballeira, J.D. Iterative saturation mutagenesis (ISM) for rapid directed evolution of functional enzymes. Nat. Protoc. 2007, 2, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Siddiqui, K.S.; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Radivojac, P.; Obradovic, Z.; Smith, D.K.; Zhu, G.; Vucetic, S.; Brown, C.J.; Lawson, J.D.; Dunker, A.K. Protein flexibility and intrinsic disorder. Protein Sci. 2004, 13, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Reetz, M.T.; Soni, P.; Fernandez, L.; Gumulya, Y.; Carballeira, J.D. Increasing the stability of an enzyme toward hostile organic solvents by directed evolution based on iterative saturation mutagenesis using the b-fit method. Chem. Commun. 2010, 46, 8657–8658. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Lang, D.A.; Liebeton, K.; Jaeger, K.E.; Dijkstra, B.M. Crystal structure of Pseudomonas aeruginosa lipase in the open conformation—The prototype for family i.1 of bacterial lipases. J. Biol. Chem. 2000, 275, 31219–31225. [Google Scholar] [CrossRef] [PubMed]
- Daniel, R.M.; Dunn, R.V.; Finney, J.L.; Smith, J.C. The role of dynamics in enzyme activity. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Feller, G. Protein stability and enzyme activity at extreme biological temperatures. J. Phys. Condes. Matter 2010, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, P. Immobilisation and application of lipases in organic media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef] [PubMed]
- Tischer, W.; Kasche, V. Immobilized enzymes: Crystals or carriers? Trends Biotechnol. 1999, 17, 326–335. [Google Scholar] [CrossRef]
- Bezerra, C.S.; de Farias Lemos, C.M.G.; de Sousa, M.; Barros Goncalves, L.R. Enzyme immobilization onto renewable polymeric matrixes: Past, present, and future trends. J. Appl. Polym. Sci. 2015, 132. [Google Scholar] [CrossRef]
- Ferrer, M.; Plou, F.J.; Fuentes, G.; Cruces, M.A.; Andersen, L.; Kirk, O.; Christensen, M.; Ballesteros, A. Effect of the immobilization method of lipase from Thermomyces lanuginosus on sucrose acylation. Biocatal. Biotransform. 2002, 20, 63–71. [Google Scholar] [CrossRef]
- Bezbradica, D.I.; Mateo, C.; Guisan, J.M. Novel support for enzyme immobilization prepared by chemical activation with cysteine and glutaraldehyde. J. Mol. Catal. B Enzym. 2014, 102, 218–224. [Google Scholar] [CrossRef]
- Godoy, C.A.; de las Rivas, B.; Guisan, J.M. Site-directing an intense multipoint covalent attachment (MCA) of mutants of the geobacillus thermocatenulatus lipase 2 (btl2): Genetic and chemical amination plus immobilization on a tailor-made support. Process Biochem. 2014, 49, 1324–1331. [Google Scholar] [CrossRef]
- Jesionowski, T.; Zdarta, J.; Krajewska, B. Enzyme immobilization by adsorption: A review. Adsorption 2014, 20, 801–821. [Google Scholar] [CrossRef]
- Cesarini, S.; Bofill, C.; Pastor, F.I.J.; Reetz, M.T.; Diaz, P. A thermostable variant of P. aeruginosa cold-adapted lipc obtained by rational design and saturation mutagenesis. Process Biochem. 2012, 47, 2064–2071. [Google Scholar] [CrossRef]
- Gangoiti, J.; Santos, M.; Llama, M.J.; Serra, J.L. Production of chiral (r)-3-hydroxyoctanoic acid monomers, catalyzed by Pseudomonas fluorescens gk13 poly(3-hydroxyoctanoic acid) depolymerase. Appl. Environ. Microbiol. 2010, 76, 3554–3560. [Google Scholar] [CrossRef] [PubMed]
- Al-Duri, B.; Goddard, R.; Bosley, J. Characterisation of a novel support for biocatalysis in supercritical carbon dioxide. J. Mol. Catal. B Enzym. 2001, 11, 825–834. [Google Scholar] [CrossRef]
- Chang, S.F.; Chang, S.W.; Yen, Y.H.; Shieh, C.J. Optimum immobilization of Candida rugosa lipase on celite by rsm. Appl. Clay Sci. 2007, 37, 67–73. [Google Scholar] [CrossRef]
- Hama, S.; Kondo, A. Enzymatic biodiesel production: An overview of potential feedstocks and process development. Bioresour. Technol. 2013, 135, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.R. Influence of blending canola, palm, soybean, and sunflower oil methyl esters on fuel properties of biodiesel. Energy Fuels 2008, 22, 4301–4306. [Google Scholar] [CrossRef]
- Haas, M.J.; McAloon, A.J.; Yee, W.C.; Foglia, T.A. A process model to estimate biodiesel production costs. Bioresour. Technol. 2006, 97, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Encinar, J.M.; González, J.F.; Rodríguez-Reinares, A. Ethanolysis of used frying oil: Biodiesel preparation and characterization. Fuel Process. Technol. 2007, 88, 513–522. [Google Scholar] [CrossRef]
- Encinar, J.M.; Sanchez, N.; Martinez, G.; Garcia, L. Study of biodiesel production from animal fats with high free fatty acid content. Bioresour. Technol. 2011, 102, 10907–10914. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Shimada, Y.; Sugihara, A.; Tominaga, Y. Conversion of degummed soybean oil to biodiesel fuel with immobilized Candida antarctica lipase. J. Mol. Catal. B Enzym. 2002, 17, 151–155. [Google Scholar] [CrossRef]
- Cesarini, S.; Diaz, P.; Nielsen, P. Exploring a new, soluble lipase for fames production in water-containing systems using crude soybean oil as a feedstock. Process Biochem. 2013, 48, 484–487. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Brask, J.; Fjerbaek, L. Enzymatic biodiesel production: Technical and economical considerations. Eur. J. Lipid Sci. Technol. 2008, 110, 692–700. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, M.T.; Mohamed, A.R. Homogeneous, heterogeneous and enzymatic catalysis for transesterification of high free fatty acid oil (waste cooking oil) to biodiesel: A review. Biotechnol. Adv. 2010, 28, 500–518. [Google Scholar] [CrossRef] [PubMed]
- Halim, S.F.A.; Kamaruddin, A.H. Catalytic studies of lipase on fame production from waste cooking palm oil in a tert-butanol system. Process Biochem. 2008, 43, 1436–1439. [Google Scholar] [CrossRef]
- Brask, J.; Damstrup, M.L.; Nielsen, P.M.; Holm, H.C.; Maes, J.; de Greyt, W. Combining enzymatic esterification with conventional alkaline transesterification in an integrated biodiesel process. Appl. Biochem. Biotechnol. 2011, 163, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.W.; Abo, M.; Du, W.; Liu, D. Enzymes in biodiesel production. In Proceedings of the World Congress on Industrial Biotechnology and Bioprocessing, Toronto, ON, Canada, 11–14 July 2006.
- Cowan, D.; Nielsen, P.M. Enzymatic degumming of edible oils and fats. In Bleaching and Purifying Fats and Oils: Theory and Practice; Patterson, H.B.W., Ed.; AOCS Press: Urbana, IL, USA, 2009; pp. 216–235. [Google Scholar]
- Watanabe, Y.; Nagao, T.; Nishida, Y.; Takagi, Y.; Shimada, Y. Enzymatic production of fatty acid methyl esters by hydrolysis of acid oil followed by esterification. J. Am. Oil Chem. Soc. 2007, 84, 1015–1021. [Google Scholar] [CrossRef]
- Luo, Y.; Zheng, Y.T.; Jiang, Z.B.; Ma, Y.S.; Wei, D.X. A novel psychrophilic lipase from Pseudomonas fluorescens with unique property in chiral resolution and biodiesel production via transesterification. Appl. Microbiol. Biotechnol. 2006, 73, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Cesarini, S.; Infanzon, B.; Pastor, F.I.J.; Diaz, P. Fast and economic immobilization methods described for non-commercial pseudomonas lipases. BMC Biotechnol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, H.; Hama, S.; Tamalampudi, S.; Noda, H. Whole-cell biocatalysts for biodiesel fuel production. Trends Biotechnol. 2008, 26, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Noureddini, H.; Gao, X.; Philkana, R.S. Immobilized Pseudomonas cepacia lipase for biodiesel fuel production from soybean oil. Bioresour. Technol. 2005, 96, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Du, W.; Liu, D.; Ding, F. Lipase-mediated methanolysis of soybean oils for biodiesel production. J. Chem. Technol. Biotechnol. 2008, 83, 71–76. [Google Scholar] [CrossRef]
- Lv, D.; Du, W.; Zhang, G.; Liu, D. Mechanism study on ns81006-mediated methanolysis of triglyceride in oil/water biphasic system for biodiesel production. Process Biochem. 2010, 45, 446–450. [Google Scholar] [CrossRef]
- Chen, X.; Du, W.; Liu, D. Effect of several factors on soluble lipase-mediated biodiesel preparation in the biphasic aqueous-oil systems. World J. Microbiol. Biotechnol. 2008, 24, 2097–2102. [Google Scholar] [CrossRef]
- Tan, T.; Xu, J.; Asano, Y. Biocatalysis in biorefinery: A green and highly efficient way to convert renewables preface. J. Mol. Catal. B 2009, 56, 77–77. [Google Scholar] [CrossRef]
- Nielsen, P.M. Industrial Biofames Production with Lipase Callera Trans L, Proceedings of the 104th AOCS Annual Meeting and Expo, Montréal, QC, Canada, 28 April–1 May 2013.
- Aalrust, E.; Beyer, W.; Ottofrickenstein, H.; Penk, G.; Plainer, H.; Reiner, R. Enzymatic Treatment of Edible Oils. U.S. Patent 5264367, 23 November 1993. [Google Scholar]
- Dijkstra, A. Continuous Removal of Gum Phase from Tri: Glyceride Oil by Mixing Alkali with the Oil. In Proceedings of the World Conference on Oilseed Technology and Utilization, Champaign, IL, USA, 1 November 1993.
- Nieuwenhuyzen, W.V.; Tomás, M.C. Update on vegetable lecithin and phospholipid technologies. Eur. J. Lipid Sci. Technol. 2008, 110, 472–486. [Google Scholar] [CrossRef]
- Clausen, K. Enzymatic oil-degumming by a novel microbial phospholipase. Eur. J. Lipid Sci. Technol. 2001, 103, 333–340. [Google Scholar] [CrossRef]
- Holm, H.C.; Nielsen, P.M.; Christensen, M.W. Production of Fatty Acid Alkyl Esters. U.S. Patent 20080199924 A1, 21 August 2008. [Google Scholar]
- Dayton, C.; Galhardo, F. Enzymatic Degumming Utilizing a Mixture of Pla and Plc Phospholipases. U.S. Patent 20080182322 A1, 31 July 2008. [Google Scholar]
- Cesarini, S.; Haller, R.F.; Diaz, P.; Nielsen, P.M. Combining phospholipases and a liquid lipase for one-step biodiesel production using crude oils. Biotechnol. Biofuels 2014, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinisetty, D.; Moldovan, D.; Devireddy, R. The effect of methanol on lipid bilayers: An atomistic investigation. Ann. Biomed. Eng. 2006, 34, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Daicheng, L.; Fucui, M. Soybean phospholipids. In Recent Trends for Enhancing the Diversity and Quality of Soybean Products; Krezhova, D., Ed.; InTech: Rijeka, Croatia, 2011; Volume 22. [Google Scholar]
- Cowan, D. Chapter 15. Lipases for the Production of Food Components; Wiley-Blackwell: Chichester, UK, 2009; pp. 332–359. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cesarini, S.; Pastor, F.I.J.; Nielsen, P.M.; Diaz, P. Moving towards a Competitive Fully Enzymatic Biodiesel Process. Sustainability 2015, 7, 7884-7903. https://0-doi-org.brum.beds.ac.uk/10.3390/su7067884
Cesarini S, Pastor FIJ, Nielsen PM, Diaz P. Moving towards a Competitive Fully Enzymatic Biodiesel Process. Sustainability. 2015; 7(6):7884-7903. https://0-doi-org.brum.beds.ac.uk/10.3390/su7067884
Chicago/Turabian StyleCesarini, Silvia, F. I. Javier Pastor, Per M. Nielsen, and Pilar Diaz. 2015. "Moving towards a Competitive Fully Enzymatic Biodiesel Process" Sustainability 7, no. 6: 7884-7903. https://0-doi-org.brum.beds.ac.uk/10.3390/su7067884