
Diversity and Abundance of Soil Animals as Influenced by Long-Term Fertilization in Grey Desert Soil, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description, Climate Characteristics and Soil Properties
2.2. Experimental Design

| Treatment | Sheep manure (kg ha−1) | Straw (kg ha−1) | N (kg ha−1) | P2O5 (kg ha−1) | K2O (kg ha−1) |
|---|---|---|---|---|---|
| CK | 0 | 0 | 0 | 0 | 0 |
| CK0 | 0 | 0 | 0 | 0 | 0 |
| N | 0 | 0 | 241.5 | 0 | 0 |
| NP | 0 | 0 | 241.5 | 138.0 | 0 |
| NK | 0 | 0 | 241.5 | 0 | 58.5 |
| PK | 0 | 0 | 0 | 138.0 | 58.5 |
| NPK | 0 | 0 | 241.5 | 138.0 | 58.5 |
| M | 60,000 | 0 | 0 | 0 | 0 |
| S | 0 | 4500–9000 | 0 | 0 | 0 |
| SNPK | 0 | 4500–9000 | 216.7 | 116.6 | 52.0 |
| MNPK | 30,000 | 0 | 84.9 | 51.4 | 12.4 |
| 1.5 MNPK | 60,000 | 0 | 151.8 | 90.4 | 19.0 |
2.3. Sampling Methods
2.4. Data Analysis
3. Results
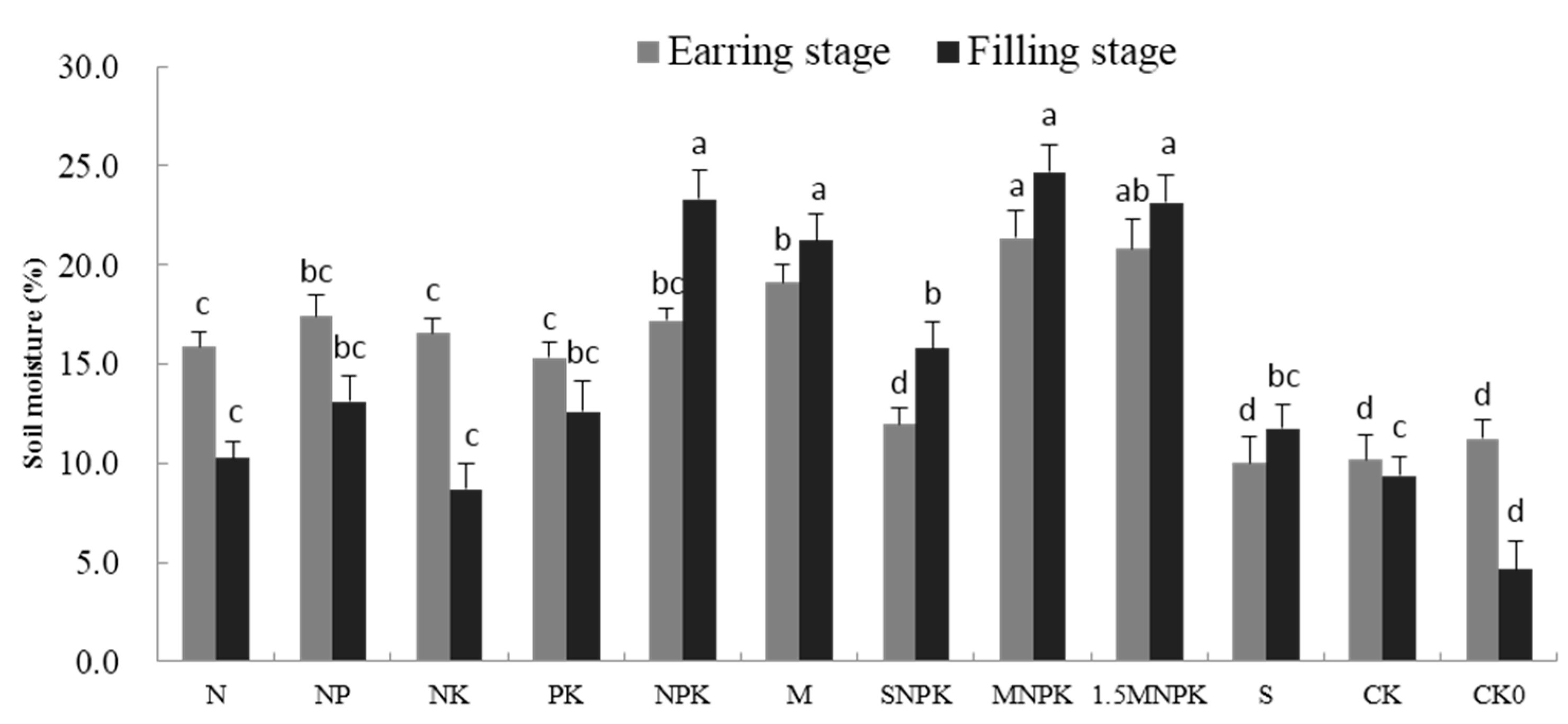
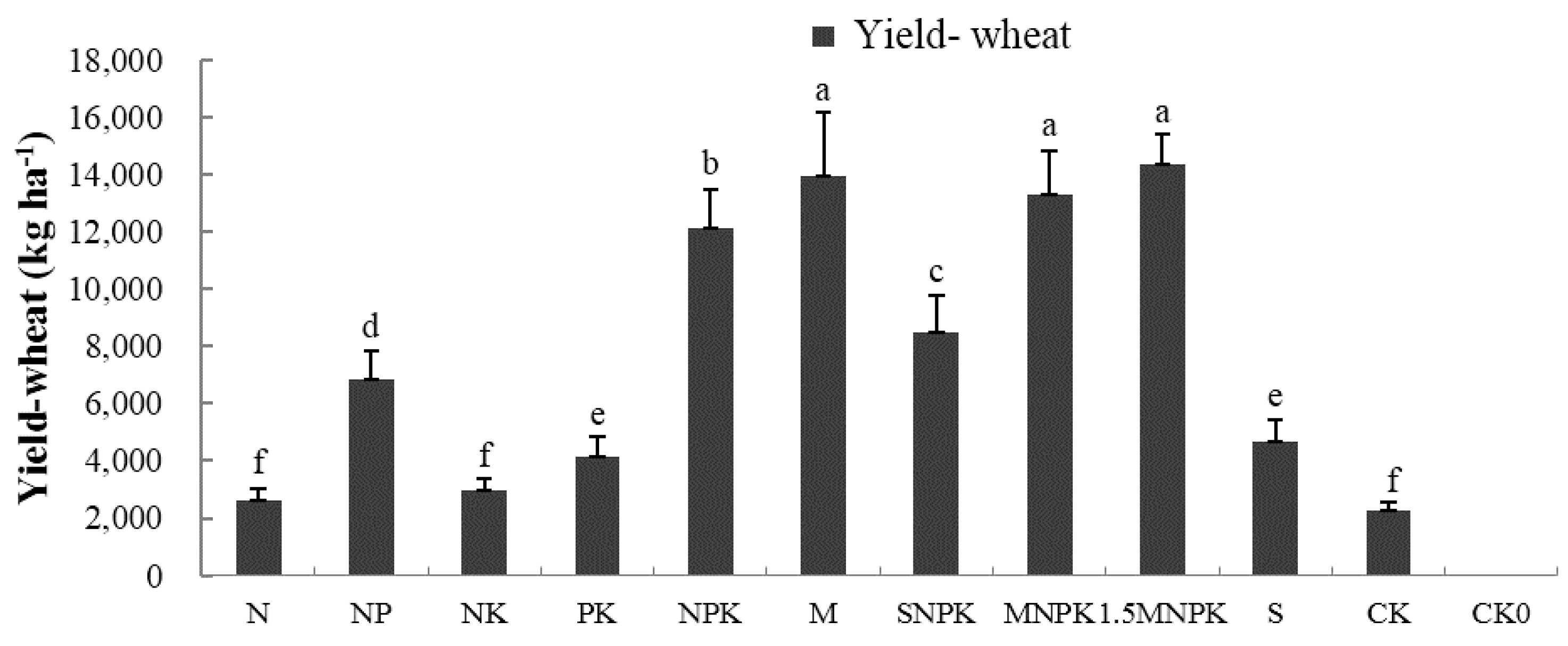
3.1. The Impact of Fertilization on the Soil Moisture and Crop Yield


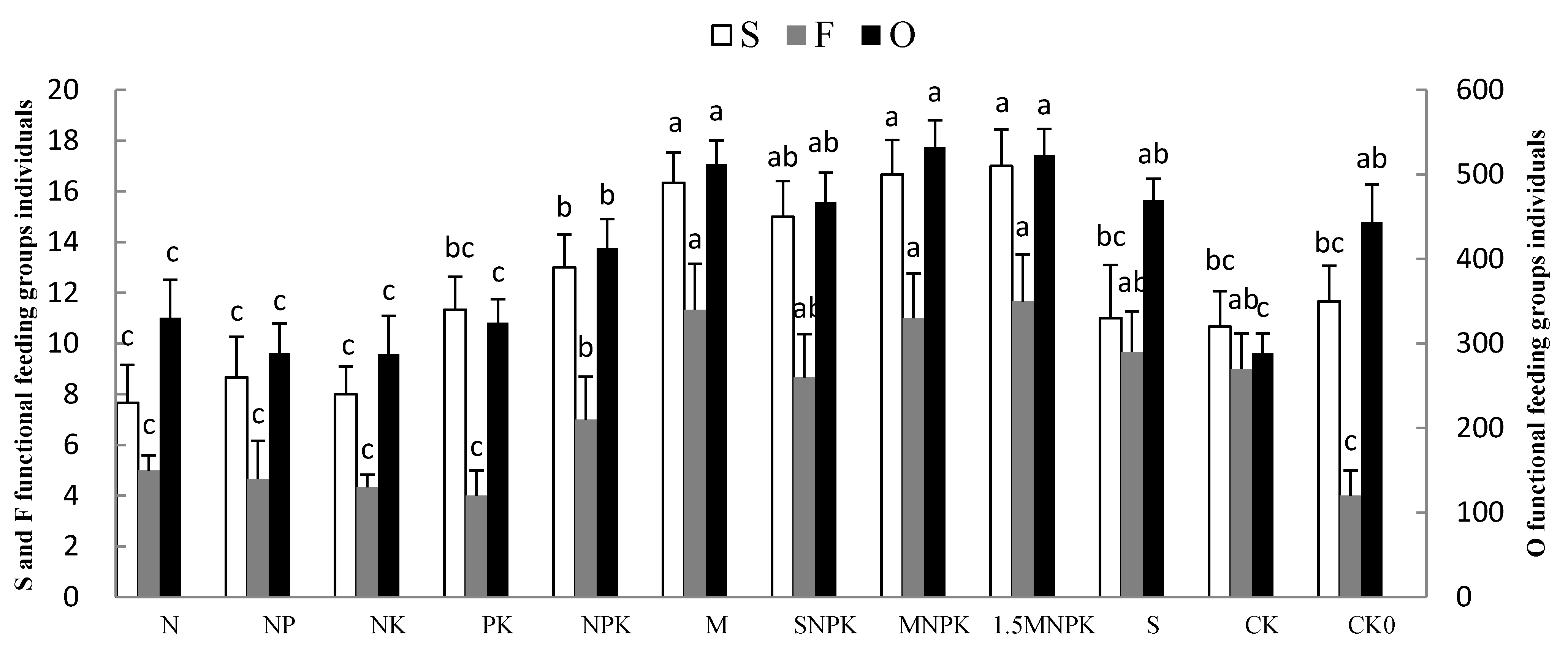
3.2. The Impact of Fertilization on Soil Fauna Community Structure and Diversity

| Taxa | Size | N | NP | NK | PK | NPK | M | SNPK | MNPK | 1.5 MNPK | S | CK | CK0 | Pet. | Deg. | Gu. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nematodes | Meso/micro | 678 | 641 | 631 | 663 | 973 | 1235 | 1125 | 1295 | 1256 | 1133 | 644 | 497 | 60.55 | *** | O |
| Oribatida | Meso/micro | 253 | 157 | 162 | 250 | 163 | 183 | 170 | 178 | 182 | 168 | 146 | 236 | 12.64 | *** | O |
| Prostigmata | Meso/micro | 6 | 2 | 1 | 2 | 5 | 2 | 7 | 8 | 3 | 2 | 0.21 | O | |||
| Mesostigmata | Meso/micro | 58 | 53 | 44 | 47 | 122 | 134 | 88 | 143 | 152 | 38 | 37 | 46 | 5.41 | ** | O |
| Collembola | ||||||||||||||||
| Onychiuridae | Meso/micro | 11 | 13 | 14 | 11 | 38 | 49 | 38 | 52 | 50 | 42 | 13 | 452 | 4.40 | ** | O |
| Entomobryidae | Meso/micro | 5 | 8 | 7 | 4 | 17 | 18 | 22 | 26 | 34 | 13 | 11 | 87 | 1.42 | ** | O |
| Thysanoptera | ||||||||||||||||
| Phlaeothripidaee | Macro | 11 | 2 | 15 | 7 | 1 | 19 | 5 | 27 | 21 | 7 | 19 | 0.75 | S | ||
| Hymenoptera | ||||||||||||||||
| Formicidae | Macro | 45 | 47 | 49 | 46 | 48 | 52 | 46 | 45 | 46 | 53 | 51 | 58 | 3.29 | ** | O |
| Formicinae | Macro | 13 | 5 | 5 | 5 | 2 | 23 | 1 | 18 | 2 | 11 | 4 | 0.50 | O | ||
| Homoptera | ||||||||||||||||
| Aphididae | Macro | 25 | 15 | 6 | 8 | 16 | 27 | 45 | 11 | 15 | 5 | 21 | 11 | 1.15 | ** | Ph |
| Coleoptera | ||||||||||||||||
| Nitidulidae | Macro | 1 | 1 | 3 | 1 | 2 | 1 | 0.05 | S | |||||||
| Nitidulidae larva | 5 | 10 | 3 | 0.10 | ||||||||||||
| Staphylinidae | Macro | 23 | 26 | 24 | 34 | 39 | 49 | 45 | 50 | 51 | 33 | 32 | 35 | 2.48 | ** | S |
| Staphylinidae larva | 2 | 0.01 | ||||||||||||||
| Scaphidiidae | Macro | 15 | 14 | 13 | 12 | 21 | 34 | 26 | 33 | 35 | 29 | 27 | 12 | 1.52 | ** | F |
| Scarabaeidae | Macro | 1 | 2 | 1 | 2 | 3 | 2 | 3 | 1 | 1 | 0.09 | Ph | ||||
| Lucanidae | Macro | 1 | 8 | 1 | 3 | 3 | 1 | 0.10 | S | |||||||
| Carabidae | Macro | 6 | 2 | 2 | 2 | 3 | 2 | 4 | 2 | 5 | 2 | 6 | 1 | 0.21 | Pr | |
| Cicindelidae | Macro | 1 | 0.01 | Pr | ||||||||||||
| Silphidae | Macro | 1 | 1 | 3 | 1 | 2 | 3 | 1 | 0.07 | Ca | ||||||
| Coccinellidae | Macro | 4 | 1 | 1 | 6 | 6 | 2 | 6 | 3 | 1 | 0.17 | Pr/Ph | ||||
| Diptera | ||||||||||||||||
| Dolichopodidae | Macro | 6 | 1 | 1 | 17 | 2 | 1 | 2 | 1 | 1 | 0.18 | Pr/S | ||||
| Orthoptera | ||||||||||||||||
| Catantopidae | Macro | 23 | 3 | 2 | 1 | 1 | 1 | 0.17 | Ph | |||||||
| Gryllidae | Macro | 1 | 1 | 1 | 2 | 3 | 0.04 | Ph | ||||||||
| Isopoda | ||||||||||||||||
| Armadillidiidae | Macro | 2 | 1 | 1 | 0.02 | O | ||||||||||
| Araneae | ||||||||||||||||
| Lycosidae | Macro | 12 | 17 | 9 | 37 | 14 | 13 | 20 | 79 | 19 | 16 | 22 | 12 | 1.52 | ** | Pr |
| Zoridae | Macro | 5 | 6 | 8 | 2 | 1 | 4 | 2 | 10 | 0.21 | Pr | |||||
| Oonopidae | Macro | 20 | 3 | 87 | 3 | 52 | 52 | 21 | 120 | 7 | 7 | 5 | 2.12 | ** | Pr | |
| Heteropodidae | Macro | 2 | 1 | 3 | 2 | 7 | 1 | 1 | 2 | 2 | 0.12 | Pr | ||||
| Prodidomidae | Macro | 1 | 1 | 2 | 1 | 1 | 4 | 1 | 5 | 1 | 0.10 | Pr | ||||
| Philodromidae | Macro | 2 | 1 | 1 | 3 | 0.04 | Pr | |||||||||
| Zoropsidae | Macro | 1 | 1 | 1 | 1 | 2 | 1 | 0.04 | Pr | |||||||
| Leptonetidae | Macro | 4 | 1 | 0.03 | Pr | |||||||||||
| Liphistiidae | Macro | 1 | 1 | 2 | 0.02 | Pr | ||||||||||
| Oxyopidae | Macro | 1 | 1 | 1 | 0.02 | Pr | ||||||||||
| Gnaphosidae | Macro | 2 | 3 | 1 | 0.03 | Pr | ||||||||||
| Opiliones | ||||||||||||||||
| Phalangiidae | Macro | 1 | 1 | 0.01 | Pr | |||||||||||
| Hemiptera | ||||||||||||||||
| Ceratocombidaee | Meso/micro | 1 | 1 | 3 | 1 | 1 | 1 | 0.04 | Ph | |||||||
| Diplura | ||||||||||||||||
| Projapygidae | Meso/micro | 1 | 1 | 2 | 0.02 | O | ||||||||||
| Symphyla | ||||||||||||||||
| Scutigerellidae | Meso/micro | 10 | 1 | 1 | 3 | 1 | 1 | 1 | 1 | 3 | 0.12 | S | ||||
| Total group | 19 | 25 | 27 | 29 | 25 | 25 | 28 | 31 | 27 | 24 | 24 | 24 | ||||
| Total Individuals | 1184 | 1054 | 991 | 1261 | 1499 | 1870 | 1778 | 1973 | 2037 | 1587 | 1054 | 1500 | 17788 | |||

| Factors | Nem. | Ony. | Sta. | Pros. | Meso. | Lyco. | Ent. | Oono. | Sca. | TI. | TG. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SM | 0.799 ** | 0.382 ** | 0.749 ** | 0.028 | 0.927 ** | 0.665 * | 0.170 * | −0.062 | 0.613 * | 0.684 * | 0.448 |
| CY-W | 0.855 ** | 0.330 ** | 0.811 ** | −0.104 | 0.946 ** | 0.727 ** | 0.103 * | 0.036 | 0.659 * | 0.757 ** | 0.444 |
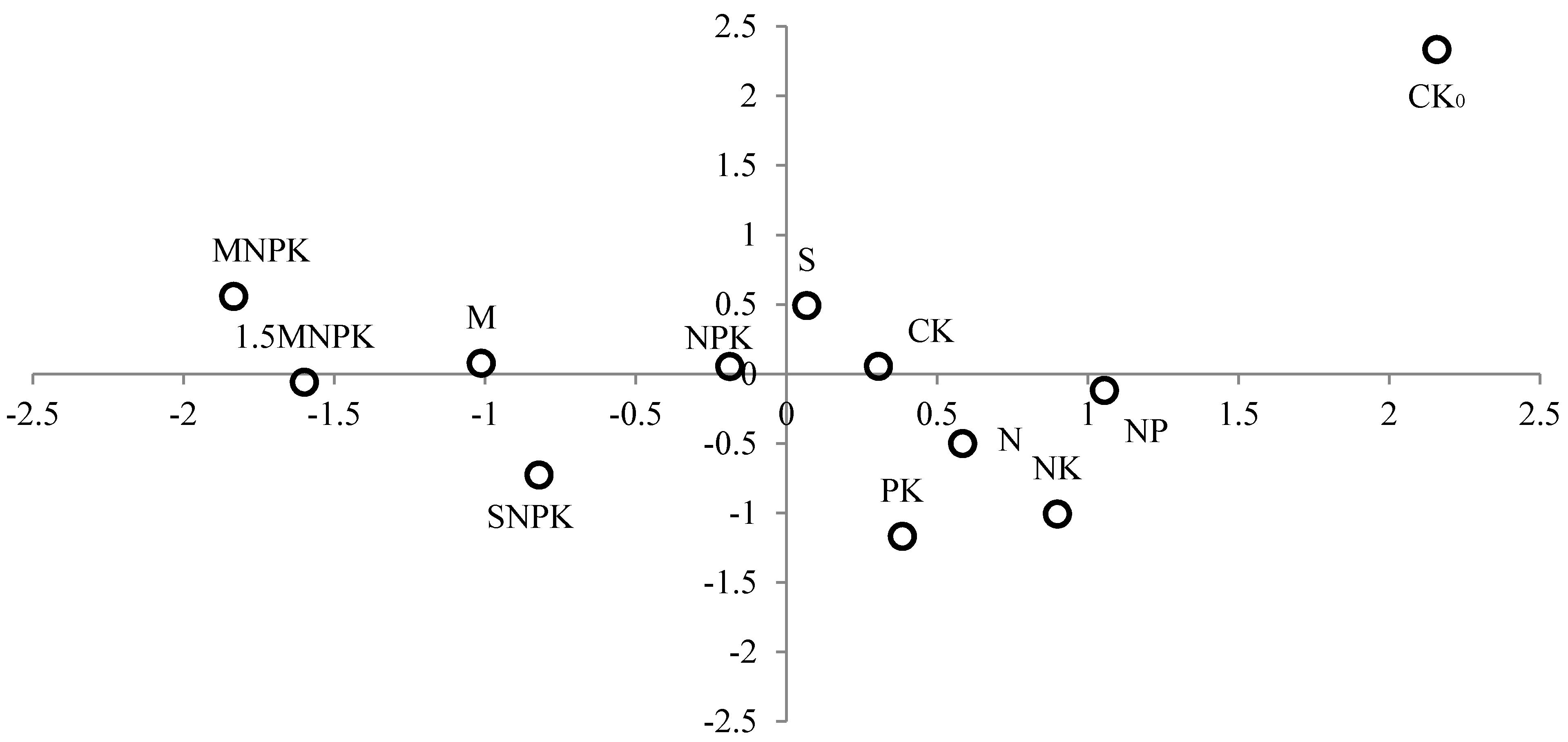
3.3. Soil Fauna Classification and Sorting
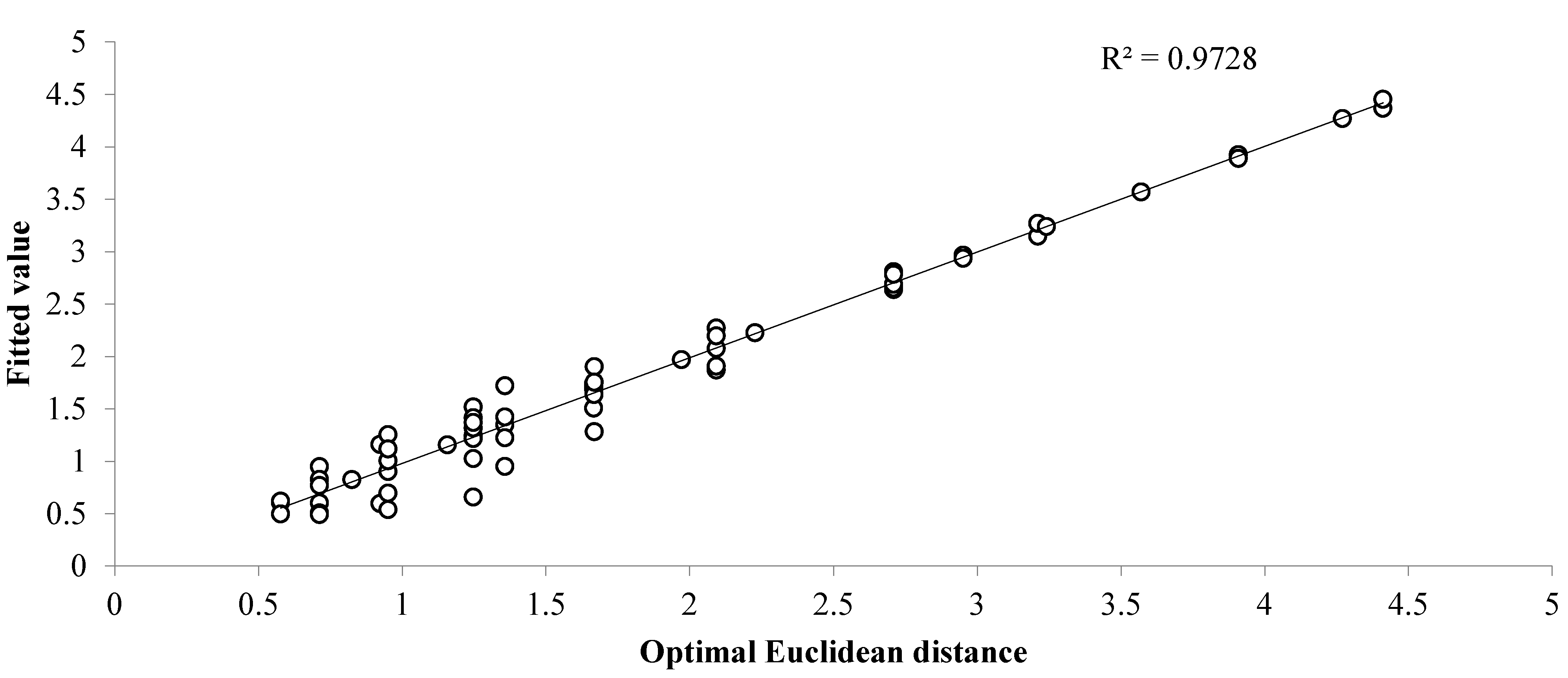
| Treatments | N | NP | NK | PK | NPK | M | SNPK | MNPK | 1.5MNPK | S | CK | CK0 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | 0 | |||||||||||
| NP | 0.923 | 0 | ||||||||||
| NK | 0.951 | 0.577 | 0 | |||||||||
| PK | 0.951 | 0.951 | 1.247 | 0 | ||||||||
| NPK | 1.247 | 0.712 | 0.951 | 1.357 | 0 | |||||||
| M | 2.093 | 1.667 | 2.093 | 2.093 | 0.825 | 0 | ||||||
| SNPK | 1.666 | 1.357 | 1.971 | 1.668 | 0.951 | 0.712 | 0 | |||||
| MNPK | 3.211 | 2.709 | 2.952 | 2.709 | 1.375 | 1.357 | 1.667 | 0 | ||||
| 1.5MNPK | 2.709 | 2.227 | 2.709 | 2.093 | 1.247 | 0.712 | 1.247 | 1.247 | 0 | |||
| S | 1.666 | 0.951 | 0.923 | 1.666 | 0.712 | 1.157 | 1.666 | 1.668 | 1.668 | 0 | ||
| CK | 1.247 | 0.577 | 0.712 | 1.357 | 0.712 | 1.247 | 1.247 | 2.093 | 2.093 | 0.577 | 0 | |
| CK0 | 3.570 | 3.241 | 2.709 | 3.908 | 3.211 | 3.908 | 4.271 | 4.411 | 4.411 | 2.709 | 2.952 | 0 |

4. Discussion
4.1. The Link between Soil Fauna and Different Fertilization Regimes
4.2. The Link between Soil Fauna and Habit Types under Different Fertilization Regimes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eekeren, N.; Bommele, L.; Bloem, J.; Schouten, T.; Rutgers, M.; Goede, R.; Reheul, D.; Brussaard, L. Soil biological quality after 36 years of ley-arable cropping, permanent grassland and permanent arable cropping. Appl. Soil Ecol. 2008, 40, 432–446. [Google Scholar] [CrossRef]
- Xin, W.D.; Yin, X.Q.; Song, B. Contribution of soil fauna to litter decomposition in Songnen sandy lands in northeastern China. J. Arid Environ. 2012, 77, 90–95. [Google Scholar] [CrossRef]
- Kiba, D.I.; Zongo, N.A.; Lompo, F.; Jansa, J.; Compaore, E.; Sedogo, P.M.; Frossard, E. The diversity of fertilization practices affects soil and crop quality in urban vegetable sites of Burkina Faso. Eur. J. Agron. 2012, 38, 12–21. [Google Scholar] [CrossRef]
- Murray, P.J.; Cook, R.; Currie, A.F.; Dawson, L.A.; Gange, A.C.; Grayston, S.J.; Treonis, A.M. Interactions between fertilizer addition, plants and the soil environment: Implications for soil fauna structure and diversity. Appl. Soil Ecol. 2006, 33, 199–207. [Google Scholar] [CrossRef]
- Ayuke, F.O.; Brussaard, L.; Vanlauwe, B.; Six, J.; Lelei, D.K.; Kibunja, C.N.; Pulleman, M.M. Soil fertility management: Impacts on soil macrofauna, soil aggregation and soil organic matter allocation. Appl. Soil Ecol. 2011, 48, 53–62. [Google Scholar] [CrossRef]
- Li, Q.; Jiang, Y.; Liang, W.J.; Lou, Y.L.; Zhang, E.P.; Liang, C.H. Long-term effect of fertility management on the soil nematode community in vegetable production under greenhouse conditions. Appl. Soil Ecol. 2010, 46, 111–118. [Google Scholar] [CrossRef]
- Williams, A.; Börjesson, G.; Hedlund, K. The effects of 55 years of different inorganic fertilizer regimes on soil properties and microbial community composition. Soil Biol. Biochem. 2013, 67, 41–46. [Google Scholar] [CrossRef]
- Coleman, D.C.; Whitman, W.B. Linking species richness, biodiversity and ecosystem function in soil systems. Pedobiologia 2005, 49, 479–497. [Google Scholar] [CrossRef]
- Fu, S.L.; Zou, X.M.; Coleman, D. Highlights and perspectives of soil biology and ecology research in China. Soil Biol. Biochem. 2009, 41, 868–876. [Google Scholar] [CrossRef]
- Kautz, T.; Fando, C.L.; Ellmer, F. Abundance and biodiversity of soil microarthropods as influenced by different types of organic manure in a long-term field experiment in Central Spain. Appl. Soil Ecol. 2006, 33, 278–285. [Google Scholar] [CrossRef]
- Froseth, R.B.; Bakken, A.K.; Bleken, M.A.; Riley, H.; Pommeresche, R.; Kristensen, K.T.; Hansen, S. Effects of green manure herbage management and its digestate from biogas production on barley yield, N recovery, soil structure and earthworm populations. Eur. J. Agron. 2014, 52, 90–102. [Google Scholar] [CrossRef]
- Blanchart, E.; Villenave, C.; Viallatoux, A.; Barthes, B.; Girardin, C.; Azontonde, A.; Feller, C. Long-term effect of a legume cover crop (Mucuna pruriens var. utilis) on the communities of soil macrofauna and nematofauna, under maize cultivation, in southern Benin. Eur. J. Soil Biol. 2006, 42, S136–S144. [Google Scholar]
- Lindberg, N.; Persson, T. Effects of long-term nutrient fertilization and irrigation on the microarthropod community in a boreal Norway spruce stand. For. Ecol. Manag. 2004, 188, 125–135. [Google Scholar] [CrossRef]
- Cao, Z.P.; Han, X.M.; Hu, C.; Chen, J.; Zhang, D.P.; Steinberger, Y. Changes in the abundance and structure of a soil mite (Acari) community under long-term organic and chemical fertilizer treatments. Appl. Soil Ecol. 2011, 49, 131–138. [Google Scholar] [CrossRef]
- Wang, B.; Liu, H.; Wang, X.H.; Li, J.M.; Ma, Y.B.; Ma, X.W. Soil phosphorus accumulation model for an arid area of north-western China with 3-year rotation of wheat, maize and cotton. J. Agric. Sci. 2014. [Google Scholar] [CrossRef]
- Lv, J.L.; Liu, X.J.; Liu, H.; Wang, X.H.; Li, K.H.; Tian, C.Y.; Christie, P. Greenhouse gas intensity and net annual global warming potential of cotton cropping systems in an extremely arid region. Nutr. Cycl. Agroecosys. 2014, 98, 15–26. [Google Scholar] [CrossRef]
- Mando, A.; Ouattara, B.; Sédogo, M.; Stroosnijder, L.; Ouattara, K.; Brussaard, L.; Vanlauwe, B. Long-term effect of tillage and manure application on soil organic fractions and crop performance under Sudano-Sahelian conditions. Soil Till Res. 2005, 80, 95–101. [Google Scholar] [CrossRef]
- Ferro, N.D.; Charrier, P.; Morari, F. Dual-scale micro-CT assessment of soil structure in a long-term fertilization experiment. Geoderma 2013, 204–205, 84–93. [Google Scholar] [CrossRef]
- Naveed, M.; Moldrup, P.; Vogel, H.J.; Lamandé, M.; Wildenschild, D.; Tuller, M.; Jonge, L.W. Impact of long-term fertilization practice on soil structure evolution. Geoderma 2014, 217–218, 181–189. [Google Scholar] [CrossRef]
- Brussaard, L.; Pulleman, M.M.; Ouédraogo, E.; Mando, A.; Six, J. Soil fauna and soil function in the fabric of the food web. Pedobiologia 2007, 50, 447–462. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soils Resources; World Soil Resource Report No. 103; FAO: Rome, Italy, 2014. [Google Scholar]
- Lin, Y.H.; Liu, H.; Zhang, S.Q.; Zhang, F.D. The community abundance and diversity of arable soil insect community following different fertilizer treatments in Xinjiang, China. Agric. Sci. China 2008, 7, 210–216. [Google Scholar] [CrossRef]
- Lin, Y.H.; Lu, P.; Yang, X.Y.; Zhang, F.D. Soil insect diversity and abundance following different fertilizer treatments on the Loess Plateau of China. J. Integr. Agr. 2013, 12, 1644–1651. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of soil analysis. Part 2: Chemical and Microbiological Properties; American Society of Agronomy/Soil Science Society of America: Madison, WI, USA, 1982. [Google Scholar]
- Yin, W.Y. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 1998. [Google Scholar]
- Zhu, X.Y.; Zhu, B. Diversity and abundance of soil fauna as influenced by long-term fertilization in cropland of purple soil, China. Soil Till Res. 2014, 3321, 1–8. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Interscience: New York, NY, USA, 1969. [Google Scholar]
- Margalef, R. Perspectives in Ecological Theory; The University of Chicago Press: Chicago, IL, USA, 1970; pp. 111–119. [Google Scholar]
- Johnson, T.E.; Hasted, A.; Ristic, R.; Bastian, S.E.P. Multidimensional scaling (MDS), cluster and descriptive analyses provide preliminary insights into Australian Shiraz wine regional characteristics. Food Qual. Preference 2013, 29, 174–185. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- Kirkpatrick, L.A.; Feenay, B.C. A Simple Guide to SPSS for Windows, for Version 12.0; Thomson Wadsworth: Stamford, CT, USA, 2005. [Google Scholar]
- Briar, S.S.; Grewal, P.S.; Somasekhar, N.; Stinner, D.; Miller, S.A. Soil nematode community, organic matter, microbial biomass and nitrogen dynamics in field plots transitioning from conventional to organic management. Appl. Soil Ecol. 2007, 37, 256–266. [Google Scholar] [CrossRef]
- Papatheodorou, E.M.; Kordatos, H.; Kouseras, T.; Monokrousos, N.; Spiroudi, U.M.; Diamantopoulos, J.; Stamou, G.P.; Argyropoulou, M.D. Differential responses of structural and functional aspects of soil microbes and nematodes to abiotic and biotic modifications of the soil environment. Appl. Soil Ecol. 2012, 61, 26–33. [Google Scholar] [CrossRef]
- Li, Q.; Bao, X.L.; Lu, C.Y.; Zhang, X.K.; Zhu, J.K.; Jiang, Y.; Liang, W.J. Soil microbial food web responses to free-air ozone enrichment can depend on the ozone-tolerance of wheat cultivars. Soil Biol. Biochem. 2012, 47, 27–35. [Google Scholar] [CrossRef]
- Hassall, M.; Adl, S.; Berg, M.; Griffiths, B.; Scheu, S. Soil fauna-microbe interactions: Towards a conceptual framework for research. Eur. J. Soil Biol. 2006, 42, S54–S60. [Google Scholar] [CrossRef]
- Carrillo, Y.; Ball, B.A.; Bradford, M.A.; Jordan, C.F.; Molina, M. Soil fauna alter the effects of litter composition on nitrogen cycling in a mineral soil. Soil Biol. Biochem. 2011, 43, 1440–1449. [Google Scholar] [CrossRef]
- Liu, H.; Lin, Y.H.; Zhang, Y.S.; Tan, X.X.; Wang, X.H. Effects of long-term fertilization on biodiversity and enzyme activity in grey desert soil. Acta Ecol. Sin. 2008, 28, 3898–3904. [Google Scholar]
- Mandal, A.; Patra, A.K.; Singh, D.; Swarup, A.; Masto, R.E. Effect of long-term application of manure and fertilizer on biological and biochemical activities in soil during crop development stages. Bioresour. Technol. 2007, 98, 3585–3592. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Liu, H.; Jia, H.T.; Jiang, P.A. Effects of long-term fertilization on soil organic matter and physical properties in grey desert. Xinjiang Agric. Sci. 2014, 51, 58–65. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Wang, X.; Liusui, Y.; Sun, X.; Zhao, C.; Liu, H. Diversity and Abundance of Soil Animals as Influenced by Long-Term Fertilization in Grey Desert Soil, China. Sustainability 2015, 7, 10837-10853. https://0-doi-org.brum.beds.ac.uk/10.3390/su70810837
Jiang M, Wang X, Liusui Y, Sun X, Zhao C, Liu H. Diversity and Abundance of Soil Animals as Influenced by Long-Term Fertilization in Grey Desert Soil, China. Sustainability. 2015; 7(8):10837-10853. https://0-doi-org.brum.beds.ac.uk/10.3390/su70810837
Chicago/Turabian StyleJiang, Maibo, Xihe Wang, Yunhao Liusui, Xueqing Sun, Chengyi Zhao, and Hua Liu. 2015. "Diversity and Abundance of Soil Animals as Influenced by Long-Term Fertilization in Grey Desert Soil, China" Sustainability 7, no. 8: 10837-10853. https://0-doi-org.brum.beds.ac.uk/10.3390/su70810837