
Soil Seed Bank and Plant Community Development in Passive Restoration of Degraded Sandy Grasslands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Site
2.2. Sampling Design and Procedure
2.3. Data Analysis
3. Results
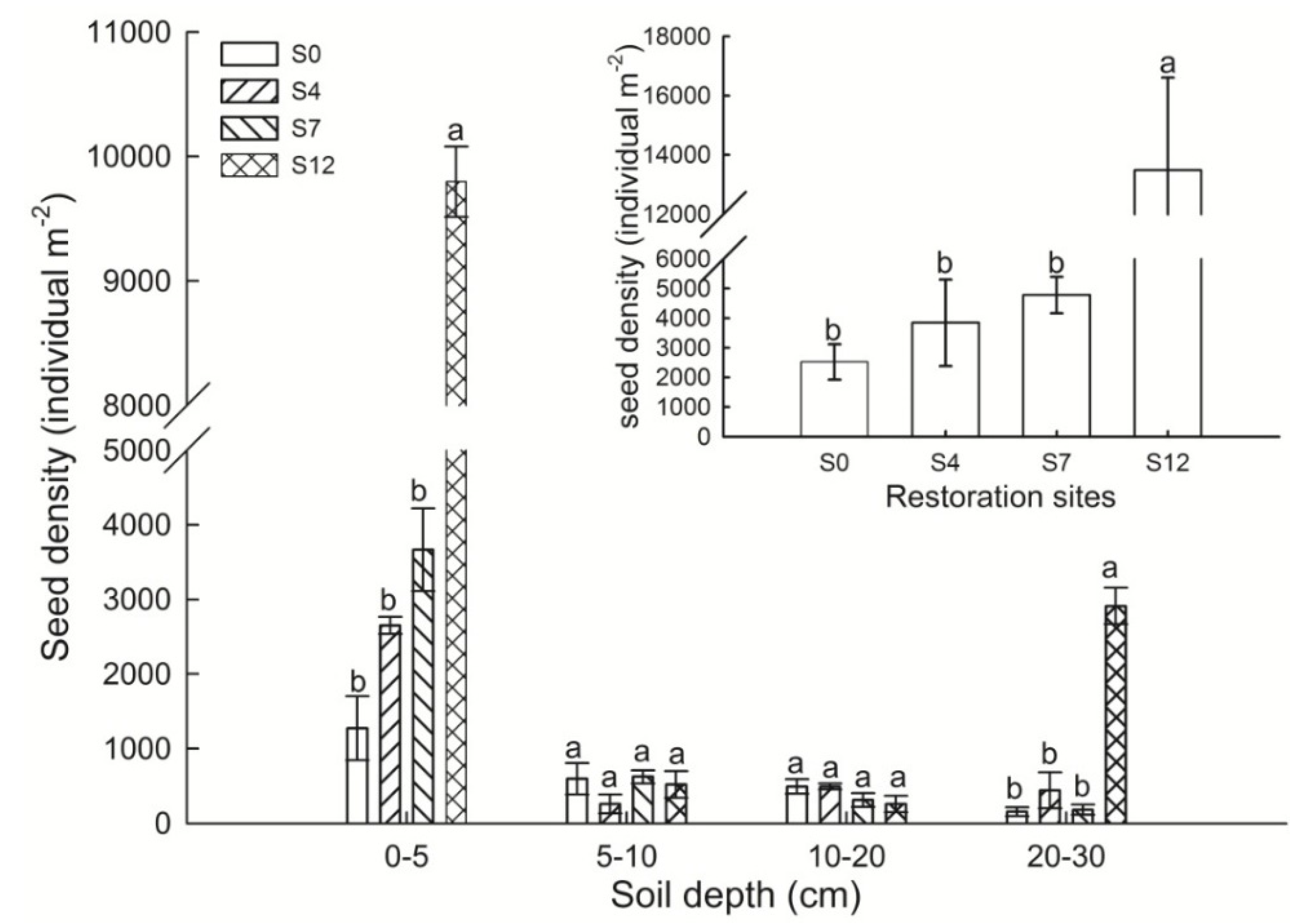
3.1. Soil Seed Bank Recovery
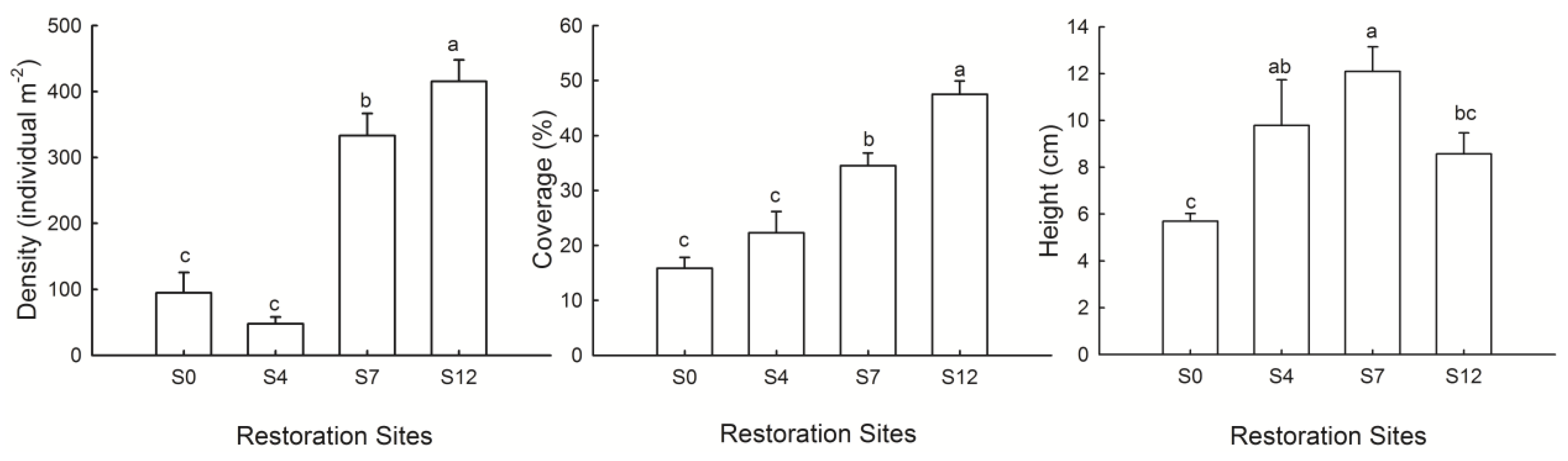
3.2. Plant Recovery
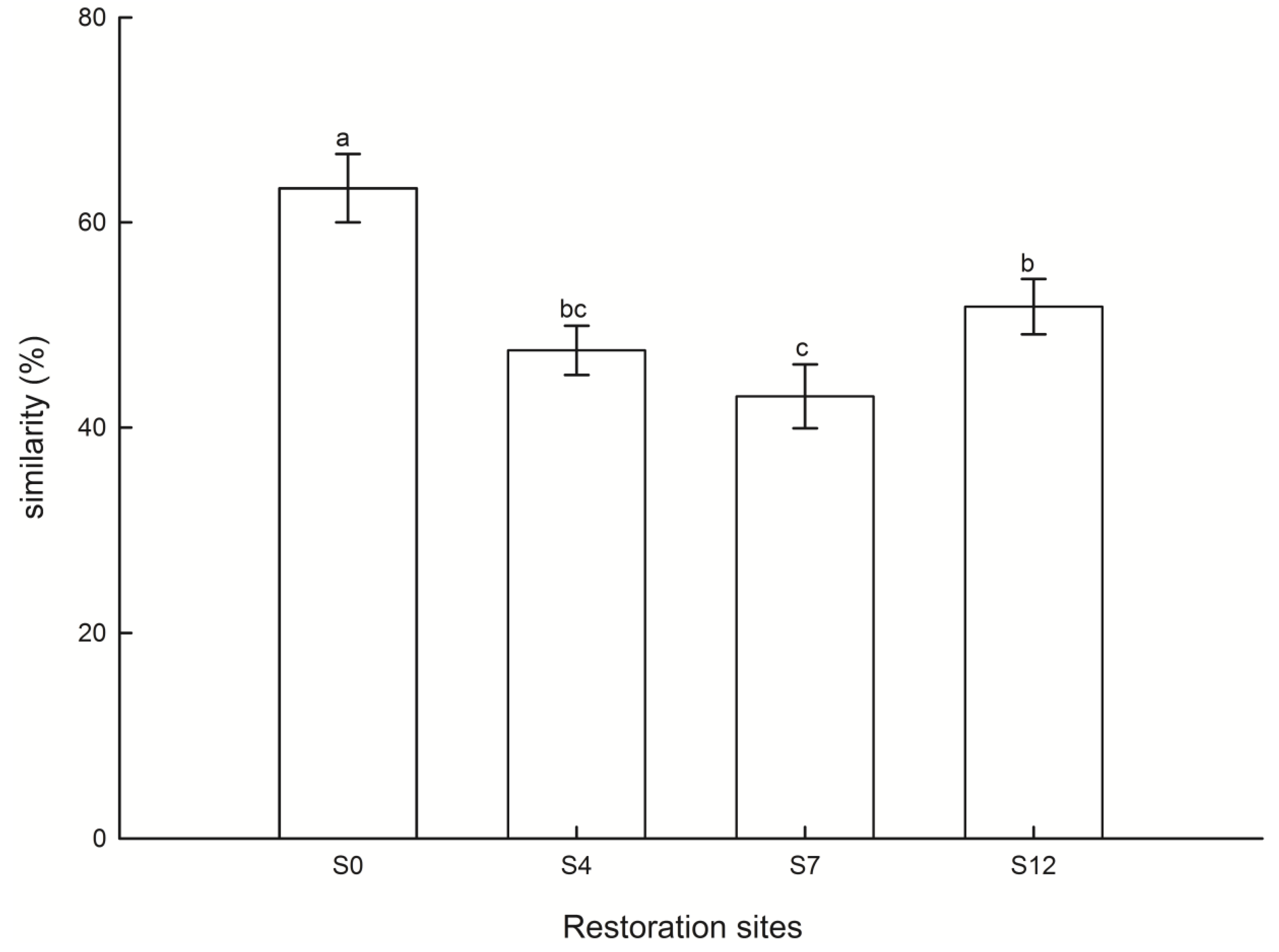
3.3. Restoration Effects on IS Values
4. Discussion
4.1. Soil Seed Bank Recovery
4.2. Plant Recovery
4.3. Similarity in Species Composition between the Soil Seed Bank and Vegetation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhao, L.P.; Su, J.S.; Wu, G.L.; Gilet, F. Long-term effects of grazing exclusion on aboveground and belowground plant species diversity in a steppe of the Loess Plateau, China. Plant Ecol. Evol. 2011, 144, 313–320. [Google Scholar] [CrossRef]
- Li, X.L.; Gao, J.; Brierley, G.; Qiao, Y.M.; Zhang, J.; Yang, Y.W. Rangeland degradation on the Qinghai-Tibet Plateau: Implications for rehabilitation. Land Degrad. Dev. 2013, 24, 72–80. [Google Scholar] [CrossRef]
- Ding, Y.J.; Mu, M.; Zhang, J.Y.; Jiang, T.; Zhang, T.J.; Wang, C.Y.; Wu, L.X.; Ye, B.S.; Bao, M.Z.; Zhang, S.Q. Impacts of Climate Change on the Environment, Economy, and Society of China. Climate and Environmental Change in China: 1951–2012; Springer: Berlin, Germany, 2015. [Google Scholar]
- Cao, C.Y.; Jiang, D.M.; Teng, X.H.; Jiang, Y.; Liang, W.J.; Cui, Z.B. Soil chemical and microbiological properties along a chronosequence of Caragana microphylla Lam. plantations in the Horqin sandy land of Northeast China. Appl. Soil Ecol. 2008, 40, 78–85. [Google Scholar] [CrossRef]
- Wu, G.L.; Li, W.; Zhao, L.P.; Shi, Z.H. Artificial management improves soil moisture, C, N and P in an alpine sandy meadow of western China. Pedosphere 2011, 21, 407–412. [Google Scholar] [CrossRef]
- Jeddi, K.; Chaieb, M. Restoring degraded arid Mediterranean areas with exotic tree species: Influence of an age sequence of Acacia salicina on soil and vegetation dynamics. Flora 2012, 207, 693–700. [Google Scholar] [CrossRef]
- Miao, R.H.; Jiang, D.M.; Musa, A.; Zhou, Q.L.; Guo, M.X.; Wang, Y.C. Effectiveness of shrub planting and grazing exclusion on degraded sandy grassland restoration in Horqin sandy land in Inner Mongolia. Ecol. Eng. 2015, 74, 164–173. [Google Scholar] [CrossRef]
- Ren, H.; Du, W.B.; Wang, J.; Yu, Z.Y.; Guo, Q.F. The natural restoration of degraded rangeland ecosystem in Heshan hill land. Acta Ecol. Sin. 2007, 27, 3593–3600. [Google Scholar]
- Wu, G.L.; Liu, Z.H.; Zhang, L.; Chen, J.M.; Hu, T.M. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China. Plant Soil 2010, 332, 331–337. [Google Scholar] [CrossRef]
- Zahawi, W.A.; Reid, J.L.; Holl, K.D. Hidden costs of passive restoration. Restor. Ecol. 2014, 22, 284–287. [Google Scholar] [CrossRef]
- Bradshaw, A. The use of natural processes in reclamation—Advantages and difficulties. Landsc. Urban Plan 2000, 51, 89–100. [Google Scholar] [CrossRef]
- Prach, K.; Hobbs, R.J. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Wang, Z.J.; Wang, S.X.; Pan, Z.B.; Guo, Y.Z.; Chen, Y.Y. Influence of different recover measures on species structure and variety in Mu Us sandy land. Chin. J. Ecol. 2005, 24, 464–466. [Google Scholar]
- Liu, M.Z.; Jiang, G.M.; Yu, S.L.; Li, Y.G.; Li, G. The role of soil seed banks in natural restoration of the degraded Hunshandak Sandlands, Northern China. Restor. Ecol. 2009, 17, 127–136. [Google Scholar] [CrossRef]
- Shang, Z.H.; Yang, S.H.; Wang, Y.L.; Shi, J.J.; Ding, L.M.; Long, R.J. Soil seed bank and its relation with above-ground vegetation along the degraded gradients of alpine meadow. Ecol. Eng. 2016, 90, 268–277. [Google Scholar] [CrossRef]
- Jeddi, K.; Chaieb, M. Changes in soil properties and vegetation following livestock grazing exclusion in degraded arid environments of South Tunisia. Flora 2010, 205, 184–189. [Google Scholar] [CrossRef]
- Li, Y.Y.; Dong, S.K.; Wen, L.; Wang, X.X.; Wu, Y. Soil seed banks in degraded and revegetated grasslands in the alpine region of the Qinghai-Tibetan Plateau. Ecol. Eng. 2012, 49, 77–83. [Google Scholar] [CrossRef]
- Meissner, R.A.; Facelli, J.M. Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrublands of South Australia. J. Arid Environ. 1999, 42, 117–128. [Google Scholar] [CrossRef]
- Russi, L.; Cocks, P.S.; Roberts, E.H. Seed bank dynamics in a Mediterranean grassland. J. Appl. Ecol. 1992, 29, 763–771. [Google Scholar] [CrossRef]
- Ma, M.J.; Zhou, X.H.; Du, G.Z. Role of soil seed bank along a disturbance gradient in alpine meadow on the Tibet plateau. Flora 2010, 205, 128–134. [Google Scholar] [CrossRef]
- Ungar, I.A.; Woodell, S.R.J. Similarity of seed banks to aboveground vegetation in grazed and ungrazed salt marsh communities on the Gower Peninsula, South Wales. Int. J. Plant Sci. 1996, 157, 746–749. [Google Scholar] [CrossRef]
- Zhao, L.P.; Cheng, J.M.; Su, J.S. The role of soil seed bank in vegetation succession under grazing exclusion in Stipa bungeana grasslands on the Loess Plateau. Acta Pratacult. Sin. 2012, 21, 38–44. [Google Scholar]
- Bossuyt, B.; Hermy, M. Seed bank assembly follows vegetation succession in dune slacks. J. Veg. Sci. 2004, 15, 449–456. [Google Scholar] [CrossRef]
- Li, Y.L. Floristic composition of vegetation and the soil seed bank in different types of dunes of Kerqin steppe. Arid Land Res. Manag. 2004, 18, 283–293. [Google Scholar] [CrossRef]
- Kristine, N.H. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar]
- Cao, C.Y.; Jiang, D.M.; Zhu, L.H.; Nan, Y.H. Degradation and diversity changes of meadow grassland in Keerqin Sandy Land. Atca Pratacult. Sin. 2006, 15, 18–26. [Google Scholar]
- Yan, Q.L.; Liu, Z.M.; Zhu, J.J.; Luo, Y.M.; Wang, H.M.; Jiang, D.M. Structure, pattern and mechanisms of formation of seed banks in sand dune systems in northeastern Inner Mongolia, China. Plant Soil 2005, 277, 175–184. [Google Scholar]
- Funes, G.; Basconcelo, S.; Diaz, S.; Cabido, M. Edaphic patchiness influences grassland regeneration from the soil seed-bank in mountain grasslands of central Argentina. Austral Ecol. 2001, 26, 205–212. [Google Scholar] [CrossRef]
- Levassor, C.; Ortega, M.; Peco, B. Seed bank dynamics of Mediterranean pastures subjected to mechanical disturbance. J. Veg. Sci. 1990, 1, 339–344. [Google Scholar] [CrossRef]
- Sacande, M.; Berrahmouni, N. Community participation and ecological criteria for selecting species and restoring natural capital with native species in the Sahel. Restor. Ecol. 2016. [Google Scholar] [CrossRef]
- Tolvanen, A.; Laine, K. The alpine soil seed standing vegetation bank in relation to field seedlings and in subarctic Finland. Arct. Antarct. Alp. Res. 2004, 36, 229–238. [Google Scholar]
- Bruno, B. Dispersal-limited species—A challenge for ecological restoration. Basic Appl. Ecol. 2014, 15, 559–564. [Google Scholar]
- Amiaud, B.; Touzard, B. The relationships between soil seed bank, aboveground vegetation and disturbances in old embanked marshlands of Western France. Flora 2004, 199, 25–35. [Google Scholar] [CrossRef]
- González-Alday, J.; Marrs, R.H.; Martínez-Ruiz, C. Soil seed bank formation during early revegetation after hydroseeding in reclaimed coal wastes. Ecol. Eng. 2009, 35, 1062–1069. [Google Scholar] [CrossRef]
- Matus, G.; Papp, M.; Tothmeresz, B. Impact of management on vegetation dynamics and seed bank formation of inland dune grassland in Hungary. Flora 2005, 200, 296–306. [Google Scholar] [CrossRef]
- Tessema, Z.K.; de Boer, W.F.; Baars, R.M.T.; Prins, H.H.T. Influence of grazing on soil seed banks determines the restoration potential of aboveground vegetation in a semi-arid savanna of Ethiopia. Biotropica 2012, 44, 211–219. [Google Scholar] [CrossRef]
- Sun, J.H.; Wang, Y.R.; Zeng, Y.J. Characteristics of the soil seed banks in degenerated grasslands under grazing and grazing suspension. Acta Bot. Boreali Occident. Sin. 2005, 25, 2035–2042. [Google Scholar]
- Tong, C.; Feng, X.; Zhang, Y.M.; Zhong, Y.K. Soil seed banks in different grazing exclusion restoring succession stages in the Xiligole degraded steppe. Acta Ecol. Sin. 2008, 28, 1991–2002. [Google Scholar]
- Zhang, J.Y.; Wang, Y.; Zhao, X.; Zhang, T. Grassland recovery by protection from grazing in a semi-arid sandy region of northern China. N. Z. J. Agric. Res. 2005, 48, 277–284. [Google Scholar] [CrossRef]
- Bai, F.; Settles, A.M. Imprinting in plants as a mechanism to generate seed phenotypic diversity. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lugo, S.; de Nascimento, L.; Mellado, M.; Bermejo, L.A.; Arevalo, J.R. Vegetation change and chemical soil composition after 4 years of goat grazing exclusion in a Canary Islands pasture. Agric. Ecosyst. Environ. 2009, 132, 276–282. [Google Scholar] [CrossRef]
- Gomaa, N.H. Soil seed bank in different habitats of the Eastern Desert of Egypt. Saudi J. Biol. Sci. 2012, 19, 211–220. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Family | Soil Seed Bank | Vegetation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| S0 | S4 | S7 | S12 | S0 | S4 | S7 | S12 | ||
| Aristida adscensionis | Poaceae | − | + | + | + | + | + | + | + |
| Artemisia arenaria | Compositae | − | − | − | − | + | + | − | − |
| Atraphaxis bracteata | Polygonaceae | − | + | − | − | − | − | − | − |
| Agriophyllum squarrosum | Amaranthaceae | + | + | + | + | + | + | − | − |
| Bassia dasyphylla | Amaranthaceae | − | − | − | + | + | − | + | + |
| Chenopodium acuminatum | Amaranthaceae | − | − | − | − | − | − | + | + |
| Corispermum candelabrum | Amaranthaceae | + | − | + | + | + | + | + | + |
| Cuscuta chinensis | Convolvulaceae | − | − | − | − | − | + | + | + |
| Caragana microphylla | Leguminosae | − | − | − | + | + | + | − | + |
| Carduus nutans | Compositae | − | − | − | − | + | + | − | + |
| Cleistogenes squarrosa | Poaceae | − | − | − | − | − | − | + | − |
| Cynanchum thesioides | Apocynaceae | − | − | − | − | + | + | + | + |
| Chloris virgata | Poaceae | − | − | − | − | − | − | + | − |
| Diarthron linifolium | Thymelaeaceae | − | + | − | − | + | + | + | − |
| Digitaria sanguinalis | Poaceae | − | − | − | − | + | + | + | + |
| Echinops gmelinii | Compositae | − | − | − | − | − | + | − | − |
| Euphorbia humifusa | Euphorbiaceae | − | − | − | − | + | + | + | + |
| Eragrostis minor | Poaceae | − | − | − | − | − | − | + | + |
| Hedysarum fruticosum | Leguminosae | − | − | − | − | − | + | − | − |
| Lespedeza davurica | Leguminosae | − | − | − | − | + | + | − | + |
| Lactuca indica | Compositae | − | − | − | − | − | − | − | + |
| Rochelia leiosperma | Boraginaceae | − | − | − | − | − | − | + | − |
| Salsola ruthenica | Amaranthaceae | − | − | − | − | − | − | + | + |
| Setaria viridis | Poaceae | + | + | + | + | + | + | + | + |
| Tribulus terrestris | Zygophyllaceae | − | − | − | − | + | + | − | − |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, R.; Song, Y.; Sun, Z.; Guo, M.; Zhou, Z.; Liu, Y. Soil Seed Bank and Plant Community Development in Passive Restoration of Degraded Sandy Grasslands. Sustainability 2016, 8, 581. https://0-doi-org.brum.beds.ac.uk/10.3390/su8060581
Miao R, Song Y, Sun Z, Guo M, Zhou Z, Liu Y. Soil Seed Bank and Plant Community Development in Passive Restoration of Degraded Sandy Grasslands. Sustainability. 2016; 8(6):581. https://0-doi-org.brum.beds.ac.uk/10.3390/su8060581
Chicago/Turabian StyleMiao, Renhui, Yongheng Song, Zhaolin Sun, Meixia Guo, Zhenxing Zhou, and Yinzhan Liu. 2016. "Soil Seed Bank and Plant Community Development in Passive Restoration of Degraded Sandy Grasslands" Sustainability 8, no. 6: 581. https://0-doi-org.brum.beds.ac.uk/10.3390/su8060581