Mass Spectrometry-Based Metabolomics of Agave Sap (Agave salmiana) after Its Inoculation with Microorganisms Isolated from Agave Sap Concentrate Selected to Enhance Anticancer Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Microorganism Isolation from Agave Sap Concentrate
2.3. Experimental Strategy for the Selection of Microorganisms that Enhanced Agave Sap Anticancer Activity
2.4. Saponin-Rich Extracts Preparation
2.5. Biological Assays
2.6. Microorganism Identification
2.7. Metabolomics and Saponins Analysis by Mass Spectrometry
2.8. Data Analysis
3. Results and Discussion
3.1. Microorganisms Isolation, Screening, and Identification
3.2. Metabolomics
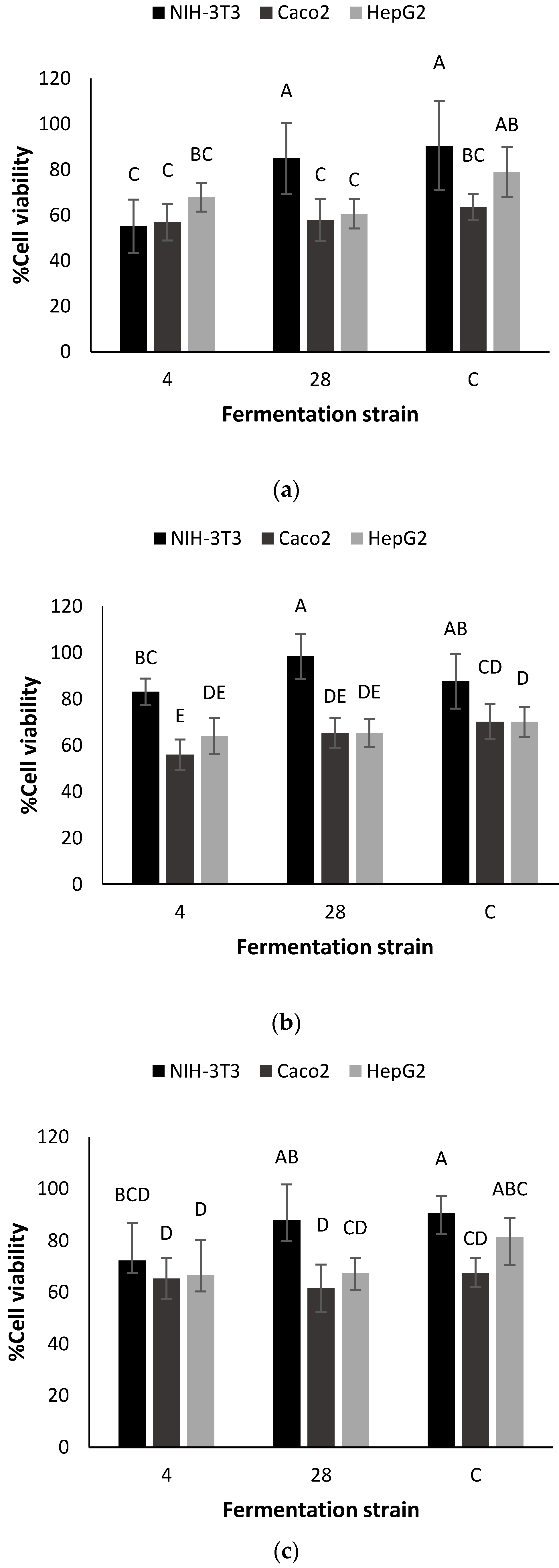
3.3. Saponins Analysis and Effects on Cancer Cell Viability
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lappe-Oliveras, P.; Moreno-Terrazas, R.; Arrizón-Gaviño, J.; Herrera-Suárez, T.; García-Mendoza, A.; Gschaedler-Mathis, A. Yeasts associated with the production of Mexican alcoholic nondistilled and distilled Agave beverages. FEMS Yeast Res. 2008, 8, 1037–1052. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.; López Soto, D.R.; Velázquez Gutiérrez, J.E.; Giles-Gómez, M.; Bolívar, F.; López-Munguía, A. Pulque, a traditional Mexican alcoholic fermented beverage: Historical, microbiological, and technical aspects. Front. Microbiol. 2016, 7, 1026. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Uribe, J.A.; Serna-Saldívar, S. Agave Syrup Extract Having Anticancer Activity. U.S. Patent 8,470,858, 25 June 2013. [Google Scholar]
- Chen, P.-Y.; Chen, C.-H.; Kuo, C.-C.; Lee, T.-H.; Kuo, Y.-H.; Lee, C.-K. Cytotoxic steroidal saponins from Agave Sisalana. Plant. Med. 2011, 77, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Santos-Zea, L.; Fajardo-Ramírez, O.R.; Romo-López, I.; Gutiérrez-Uribe, J.A. Fast centrifugal partition chromatography fractionation of concentrated agave (Agave salmiana) sap to obtain saponins with apoptotic effect on colon cancer cells. Plant Foods Hum. Nutr. 2016, 71, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Santos-Zea, L.; Rosas-Pérez, A.M.; Leal-Díaz, A.M.; Gutiérrez-Uribe, J.A. Variability in saponin content, cancer antiproliferative activity and physicochemical properties of concentrated agave sap. J. Food Sci. 2016, 81, H2069–H2075. [Google Scholar] [CrossRef] [PubMed]
- Leal-Díaz, A.M.; Santos-Zea, L.; Martínez-Escobedo, H.C.; Guajardo-Flores, D.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O. Effect of Agave americana and Agave salmiana ripeness on saponin content from aguamiel (agave sap). J. Agric. Food Chem. 2015, 63, 3924–3930. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Xu, G. Mass-spectrometry-based microbial metabolomics: Recent developments and applications. Anal. Bioanal. Chem. 2015, 407, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Metabolomics: Applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Settachaimongkon, S.; Nout, M.J.R.; Antunes Fernandes, E.C.; Hettinga, K.A.; Vervoort, J.M.; van Hooijdonk, T.C.M.; Zwietering, M.H.; Smid, E.J.; van Valenberg, H.J.F. Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp. bulgaricus on the metabolite profile of set-yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.; Giles-Gómez, M.; Hernández, G.; Córdova-Aguilar, M.S.; López-Munguía, A.; Gosset, G.; Bolívar, F. Analysis of bacterial community during the fermentation of pulque, a traditional Mexican alcoholic beverage, using a polyphasic approach. Int. J. Food Microbiol. 2008, 124, 126–134. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, M.; Qi, Y.; Dong, Y.; He, X.; Li, J.; Liu, X.; Sun, L. Differential responses of soil microbial community to four-decade long grazing and cultivation in a semi-arid grassland. Sustainability 2017, 9, 128. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Moreno-García, B.E.; Gutiérrez-Uribe, J.A.; Aráiz-Hernández, D.; Alvarez, M.M.; Serna-Saldivar, S.O. Induction of apoptosis in colon cancer cells treated with isorhamnetin glycosides from Opuntia ficus-indica pads. Plant Foods Hum. Nutr. 2014, 69, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.; Rodríguez, M.E.; Martínez, A.; López-Munguía, A.; Bolívar, F.; Gosset, G. Characterization of bacterial diversity in pulque, a traditional Mexican alcoholic fermented beverage, as determined by 16S rDNA analysis. FEMS Microbiol. Lett. 2004, 235, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular volutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.J.; Calle, J.M.; Simonet, A.M.; Guerra, J.O.; Stochmal, A.; Macías, F.A. Bioactive steroidal saponins from Agave offoyana flowers. Phytochemistry 2013, 95, 298–307. [Google Scholar] [CrossRef] [PubMed]
- METLIN: A metabolite Mass Spectral Database. Available online: http://metlin.scripps.edu (accesed on 1 May 2017).
- Yoo, S.-H.; Weon, H.-Y.; Noh, H.-J.; Hong, S.-B.; Lee, C.-M.; Kim, B.-Y.; Kwon, S.-W.; Go, S.-J. Roseomonas aerilata sp. nov., isolated from an air sample. Int. J. Syst. Evol. Microbiol. 2008, 58, 1482–1485. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.; Santo, C.E.; Grass, G.; Chung, A.P.; Morais, P.V. Roseomonas pecuniae sp. nov., isolated from the surface of a copper-alloy coin. Int. J. Syst. Evol. Microbiol. 2011, 61, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, M.; Notaguchi, E.; Sato, A.; Yoshioka, M.; Hasegawa, A.; Kagami, T.; Narita, T.; Yamashita, M.; Sei, K.; Soda, S.; et al. Characterization of Pseudomonas stutzeri NT-I capable of removing soluble selenium from the/aqueous phase under aerobic conditions. J. Biosci. Bioeng. 2011, 112, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, X.; Xin, Y.; Xue, S.; Zhang, W. Purification and characterization of a dehalogenase from Pseudomonas stutzeri DEH130 isolated from the marine sponge Hymeniacidon perlevis. World J. Microbiol. Biotechnol. 2013, 29, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L.A.; Uchino, M.; Falsen, E.; Lysenko, A.M.; Zhukova, N.V.; Mikhailov, V.V. Pseudomonas xanthomarina sp. nov., a novel bacterium isolated from marine ascidian. J. Gen. Appl. Microbiol. 2005, 51, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Ghanem, M.; Gadda, G. Cloning, sequence analysis, and purification of choline oxidase from Arthrobacter globiformis: A bacterial enzyme involved in osmotic stress tolerance. Arch. Biochem. Biophys. 2004, 421, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, G.; Salminen, S.; O’Brien, J.; Ouwenhand, A.; Holzapfel, W.; Shortt, C.; Fonden, R.; Miller, G.D.; Donohue, D.; Playne, M.; et al. Inventory of microorganisms with a documented history of use in food. Bull. Int. Dairy Fed. 2002, 377, 10–19. [Google Scholar]
- Roes, M.; Goodwin, C.M.; Meyers, P.R. Gordonia lacunae sp. nov., isolated from an estuary. Syst. Appl. Microbiol. 2008, 31, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.H.; Sun, W.J.; Yu, B.; Cui, F.J.; Qian, J.Y.; Liu, J.Z.; Wang, L.; Qi, X.H.; Wei, H. Continuous conversion of rice starch hydrolysate to 2-keto-d-gluconic acid by Arthrobacter globiformis C224. Biotechnol. Bioprocess Eng. 2013, 18, 709–714. [Google Scholar] [CrossRef]
- Takaichi, S.; Maoka, T.; Akimoto, N.; Carmona, M.L.; Yamaoka, Y. Carotenoids in a Corynebacterineae, Gordonia terrae AIST-1: Carotenoid glucosyl mycoloyl esters. Biosci. Biotechnol. Biochem. 2008, 72, 2615–2622. [Google Scholar] [CrossRef] [PubMed]
- Canela, N.; Herrero, P.; Mariné, S.; Nadal, P.; Ras, M.R.; Rodríguez, M.Á.; Arola, L. Analytical methods in sphingolipidomics: Quantitative and profiling approaches in food analysis. J. Chromatogr. A 2016, 1428, 16–38. [Google Scholar] [CrossRef] [PubMed]
- Aida, K.; Kinoshita, M.; Susgawara, T.; Ono, J.; Miyazawa, T.; Ohnishi, M. Apoptosis Inducement by Plant and Fungus Sphingoid Bases in Human Colon Cancer Cells. J. Oleo Sci. 2004, 53, 503–510. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Suggested Compounds | Accurate Mass | m/z |
|---|---|---|---|
| 1 | Dehydro-phytosphingosine | 315.27 | 316.28 (M + H)+, 338.25 (M + Na)+ |
| 2 | Magueyoside A | 1194.48 | 445.29 (M-3Hex-2Pen + H)+, 607.34 (M-2Hex-2Pen + H)+, 769.35 (M-1Hex-2Pen + H)+, 1217.51 (M + Na)+ |
| Magueyoside B | 1062.44 | 445.29 (M-3Hex-1Pen + H)+, 607.34 (M-2Hex-1Pen + H)+, 769.36 (M-1Hex-1Pen + H)+, 1085.48 (M + Na)+ | |
| 3 | Kammogenin | 444.28 | 445.29 (M + H)+, 467.25 (M + Na)+, 483.23 (M + K)+, 911.52 (2M + Na)+ |
| 4 | Kammogenin glycoside 1 (KG1) | 1092.44 | 445.29 (M-4Hex + H)+, 607.34 (M-3Hex + H)+, 769.36 (M-2Hex + H)+, 931.40 (M-1Hex + H)+, 1115.51 (M + Na)+ |
| Kammogenin glycoside 2 (KG2) | 1224.48 | 445.29 (M-4Hex-1Pen + H)+, 607.34 (M-3Hex-1Pen + H)+, 769.36 (M-2Hex-1Pen + H)+, 1247.47 (M + Na)+ |
| Strain | Fermentation Time (h) | MagA + MagB § | KG1 + KG2 ⌷ |
|---|---|---|---|
| (µg PE/mg Extract) | |||
| A. globiformis | 48 | 2.99 ± 0.68 a | 2.05 ± 0.30 a,b |
| 96 | 2.73 ± 0.58 a | 1.81 ± 0.32 a,b | |
| 192 | 2.33 ± 0.71 a | 1.71 ± 0.57 a,b | |
| Gordonia sp. | 48 | 2.71 ± 0.29 a | 2.04 ± 0.21 a,b |
| 96 | 3.09 ± 0.65 a | 2.43 ± 0.67 a,b | |
| 192 | 2.70 ± 1.01 a | 2.80 ± 0.71 a | |
| Control | 48 | 3.56 ± 0.05 a | 2.51 ± 0.11 a,b |
| 96 | 2.57 ± 0.39 a | 1.84 ± 0.20 a,b | |
| 192 | 1.70 ± 0.59 a | 1.27 ± 0.41 b | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa, L.M.; Santos-Zea, L.; Escalante, A.; Gutiérrez-Uribe, J.A. Mass Spectrometry-Based Metabolomics of Agave Sap (Agave salmiana) after Its Inoculation with Microorganisms Isolated from Agave Sap Concentrate Selected to Enhance Anticancer Activity. Sustainability 2017, 9, 2095. https://0-doi-org.brum.beds.ac.uk/10.3390/su9112095
Figueroa LM, Santos-Zea L, Escalante A, Gutiérrez-Uribe JA. Mass Spectrometry-Based Metabolomics of Agave Sap (Agave salmiana) after Its Inoculation with Microorganisms Isolated from Agave Sap Concentrate Selected to Enhance Anticancer Activity. Sustainability. 2017; 9(11):2095. https://0-doi-org.brum.beds.ac.uk/10.3390/su9112095
Chicago/Turabian StyleFigueroa, Luis M., Liliana Santos-Zea, Adelfo Escalante, and Janet A. Gutiérrez-Uribe. 2017. "Mass Spectrometry-Based Metabolomics of Agave Sap (Agave salmiana) after Its Inoculation with Microorganisms Isolated from Agave Sap Concentrate Selected to Enhance Anticancer Activity" Sustainability 9, no. 11: 2095. https://0-doi-org.brum.beds.ac.uk/10.3390/su9112095