1. Introduction
Eutrophication in man-made reservoirs has received considerable attention over time due to its harmful effects on the aquatic environment and on human and animal health [
1,
2]. The increased probability of algal blooms occurring is of major concern, especially where these blooms are due to (toxic) cyanobacteria species. Under natural conditions in aquatic ecosystems, a balance exists between cyanobacteria and other phytoplankton groups [
3]. However, specific characteristics may allow cyanobacteria to become prevalent. These characteristics are determined by a range of features, including cellular physiology (gas vesicles within cells allow regulation of buoyancy) and physiological response (light and nutrient utilization, for example), cell size, cell structure, and general morphology [
4]. The predominance of cyanobacteria over other species occurs under specific environmental conditions, including optimal light intensity and water temperature, nutrient availability and stability in the water column [
2].
Cyanobacteria harmful algal blooms (CHABs) form an increasing problem globally in all types of water bodies due to mounting eutrophication [
5,
6,
7,
8]. Moreover, they are responsible for a variety of impacts on environmental, economic and social scales [
9]. They greatly impact on zooplankton and fish populations in aquatic ecosystems [
10]. The presence or absence of particular cyanobacteria species may signal the ecological status of a water body; the dominance of cyanobacteria has been particularly useful as an indicator for a high nutrient status [
11]. Thus, monitoring of cyanobacteria blooms in drinking water and reservoirs with secondary uses is necessary [
12]. However, estimating the occurrence of CHABs hot spots in unmeasured locations using traditional sampling methods is very difficult [
12,
13,
14].
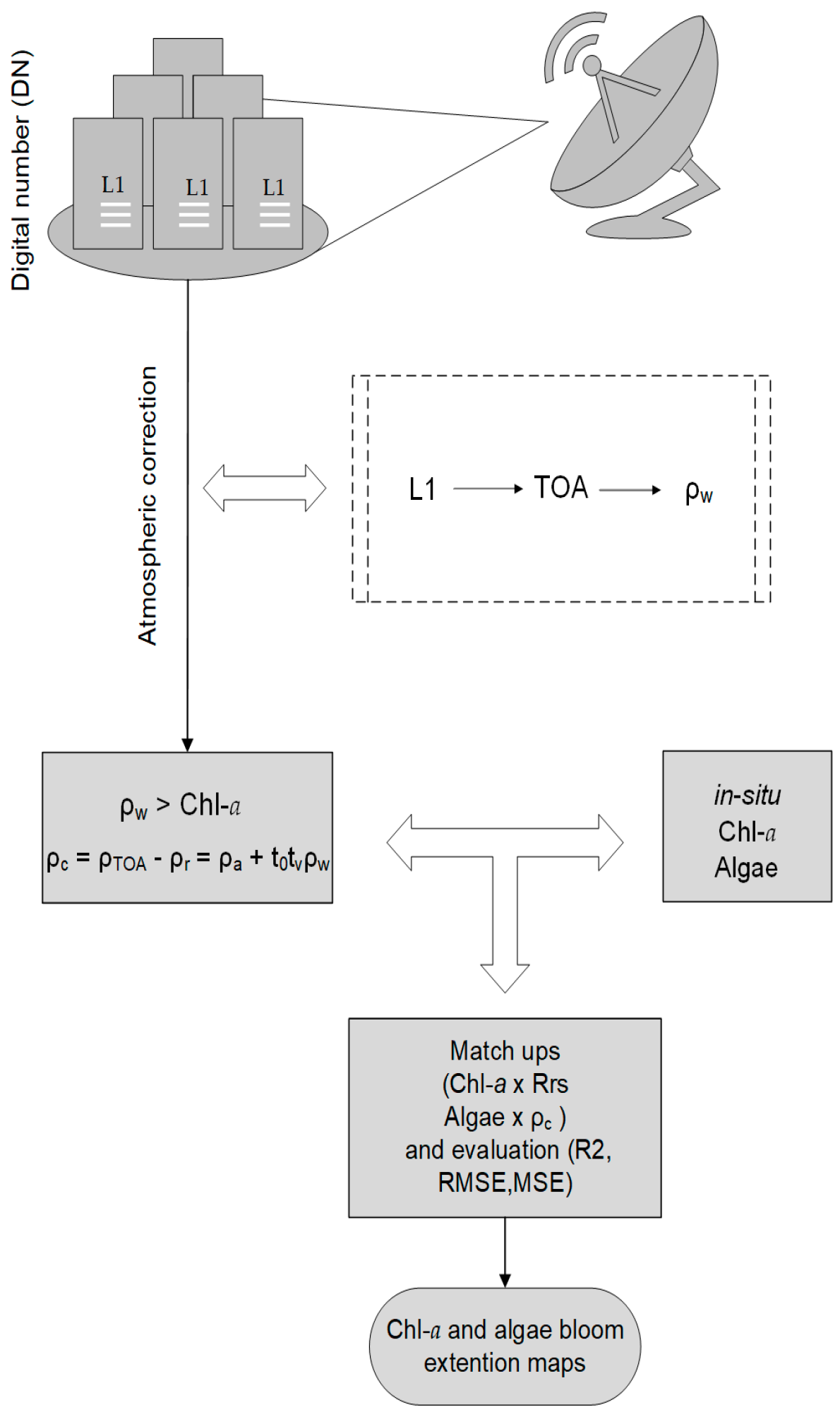
Satellite remote sensing techniques offer suitable tools to integrate large-scale water quality monitoring data [
15,
16] and have been widely used to estimate Chlorophyll-
a [
16,
17,
18,
19,
20] and phycocyanin [
21,
22,
23,
24,
25,
26,
27,
28,
29]. Cyanobacteria blooms have been studied using the bio-optical approach based on retrieval algorithms for phycocyanin, a specific pigment of cyanobacteria. Very few methods use multispectral data, such as Landsat, to estimate cyanobacteria density from atmospherically corrected surface reflectance values, as proposed by [
30,
31,
32]. The main challenge is still the low availability of sensors with suitable spectral bands to retrieve cyanobacteria data using field-based approaches [
33].
The Operational Land Imager (OLI) sensor onboard the Landsat-8 satellite has shown potential regarding application in studies on aquatic environments [
34]. OLI images were used by [
35] to estimate cyanobacteria density by means of empirical models and [
36] emphasized the value of OLI images based on the blue to green spectral region for assessing waters with a low to medium amount of biomass of blue-green algae.
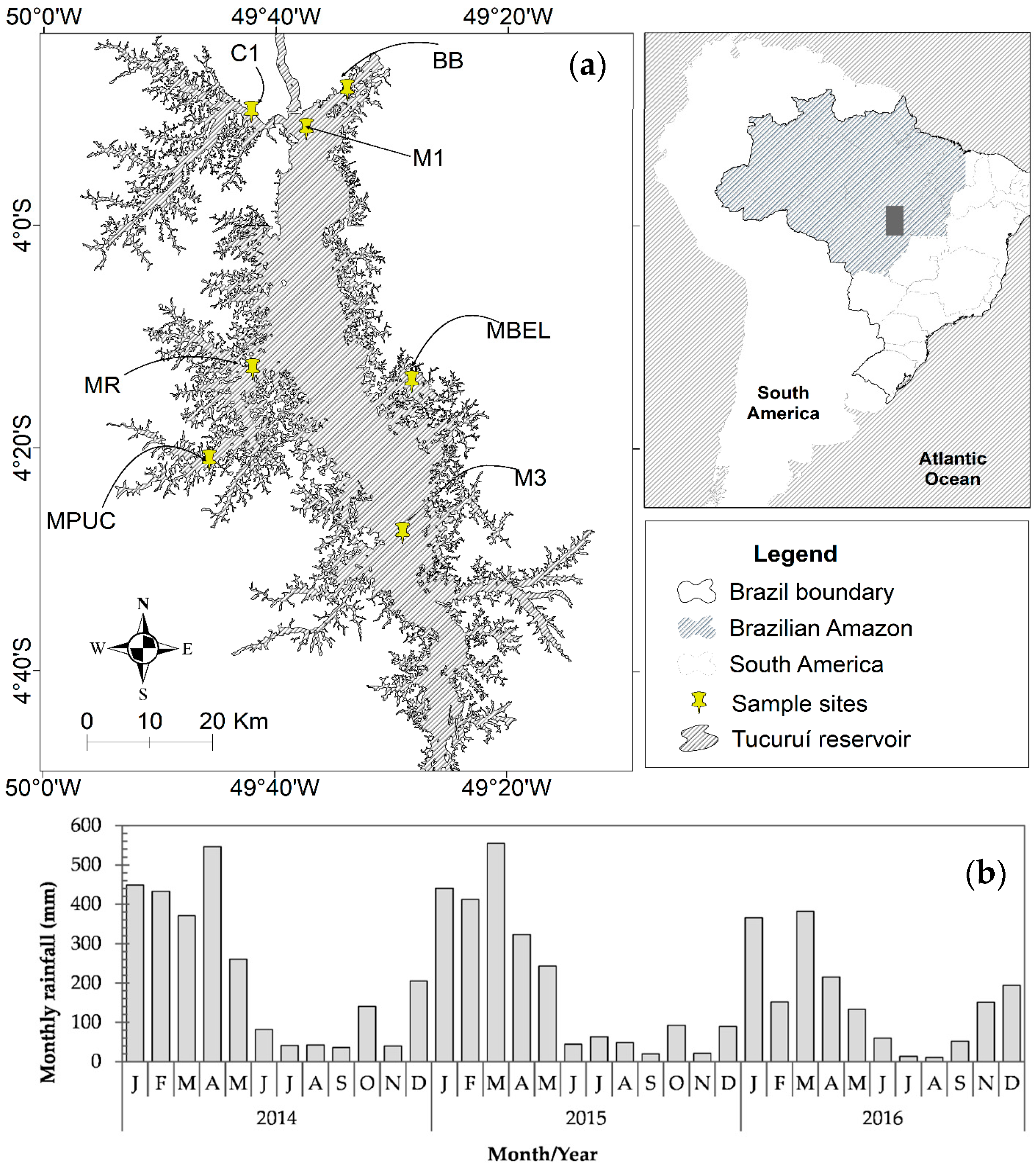
The study of CHABs in the Tucuruí hydroelectric reservoir (THR) in Brazil is of great importance, due to the multiple uses of its water by the local population. However, the reservoir has only been mentioned in a few studies [
37,
38] concerning phytoplankton density and never specifically regarding CHABs. Monitoring cyanobacteria density with satellite images has not been undertaken at all. Here we propose a multidisciplinary approach that aims to assess algal bloom extent and which includes ecological and optical studies, in a spatial and temporal context as supported by [
39]. The first approach was based on the water limnology of the THR, consisting of physico-chemical parameters and phytoplankton studies. The latter was based on the categorization of phytoplankton according to their species’ survival strategy regarding different environmental changes. This categorization consists of a list of 31 functional phytoplankton groups, represented by alphanumeric terms, established by [
40]. The use of this classification has as main objective to detect patterns in the phytoplankton dynamic and distribution, as well as relate them to environmental changes. The second approach was based on the algal bloom extent and Chlorophyll-
a estimated from OLI data.
The main goal of this study was to investigate if the combination between water limnology and satellite imagery is a suitable approach to identify CHAB extent in the Tucuruí hydroelectric reservoir. This motivation was based on the characteristics of the THR water conditions (with low/medium Chlorophyll-
a) and using the literature of [
32,
36]. The application of satellite remote sensing techniques may help to compensate for the limited spatial dimension of traditional in situ methods. It permits acquisition of necessary information at different spatial and temporal scales, allowing a more complete analysis of aquatic ecosystems, and it is a functional analysis in synoptic order. We also aim to map Chlorophyll-
a on a spatial and temporal scale, as it is closely related to cyanobacteria occurrence. The outcomes of this study aim to support water management regarding CHABs impact in the Tucuruí hydroelectric reservoir.
4. Discussion
Water quality monitoring in reservoirs is an important approach to understanding the complexity of this environment. These environments are classified as ecosystems with permanent disturbance on a vertical and horizontal scale [
64] and their water quality is greatly influenced by human interference.
In the case of the THR, little human interference affects its water quality, due to minimal occupation near its edges and insignificant changes in land use and cover, as is described by [
65]. When the THR is full, the main source of nutrient loading is rainfall, while in the emptying phase it is the water level, which releases nutrients from the bottom [
66]. The THR is characterized by areas with different limnological patterns as a result of its morphometry, and areas which were not deforested prior to the flooding [
43,
67].
This study proposes a multidisciplinary approach to assess the feasibility for the OLI sensor to monitor algal blooms in the THR. To this effect, an ecological and optical approach in both spatial and temporal scale is applied. An understanding of the ecological characteristics of reservoirs, including bio-physical and chemical features, is important for their water management. Biological studies are important to assess uses of water in reservoirs due to their close relation to the effects of algal blooms. Enhanced phytoplankton growth is a major concern for policy and management particularly when the reservoir is used for recreation, aquaculture or potable supplies [
68].
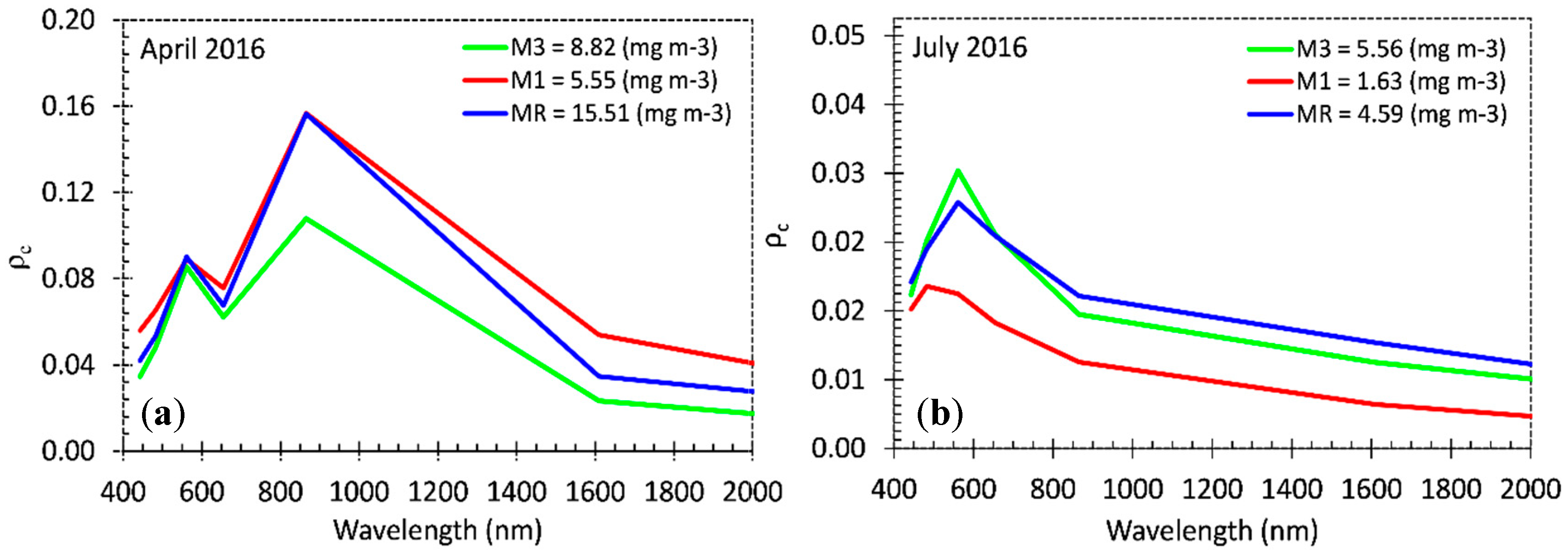
Our results show that Chl-
a concentrations were higher in April (full reservoir and at the river zone) at 15.51 mg m
−3 than in July at 4.10 mg m
−3. Significant variation (
p < 0.05) of Chl-
a concentration was found between months but not in spatial scale. In the full phase of the reservoir, Chl-
a tends to increase at certain sites near dendritic and edge areas. In April, Chl-
a concentrations in the water were in the range of eutrophic, while in July the range reached oligotrophic to mesotrophic values. Da Costa Lobato et al. [
42] showed similar results in the emptying phase of the reservoir (June to August), classifying this reservoir as mesotrophic with few oligotrophic sites. Low trophic levels in this reservoir could be related to its stabilization process, which is a positive factor for the maintenance of the phytoplankton diversity and biomass [
2].
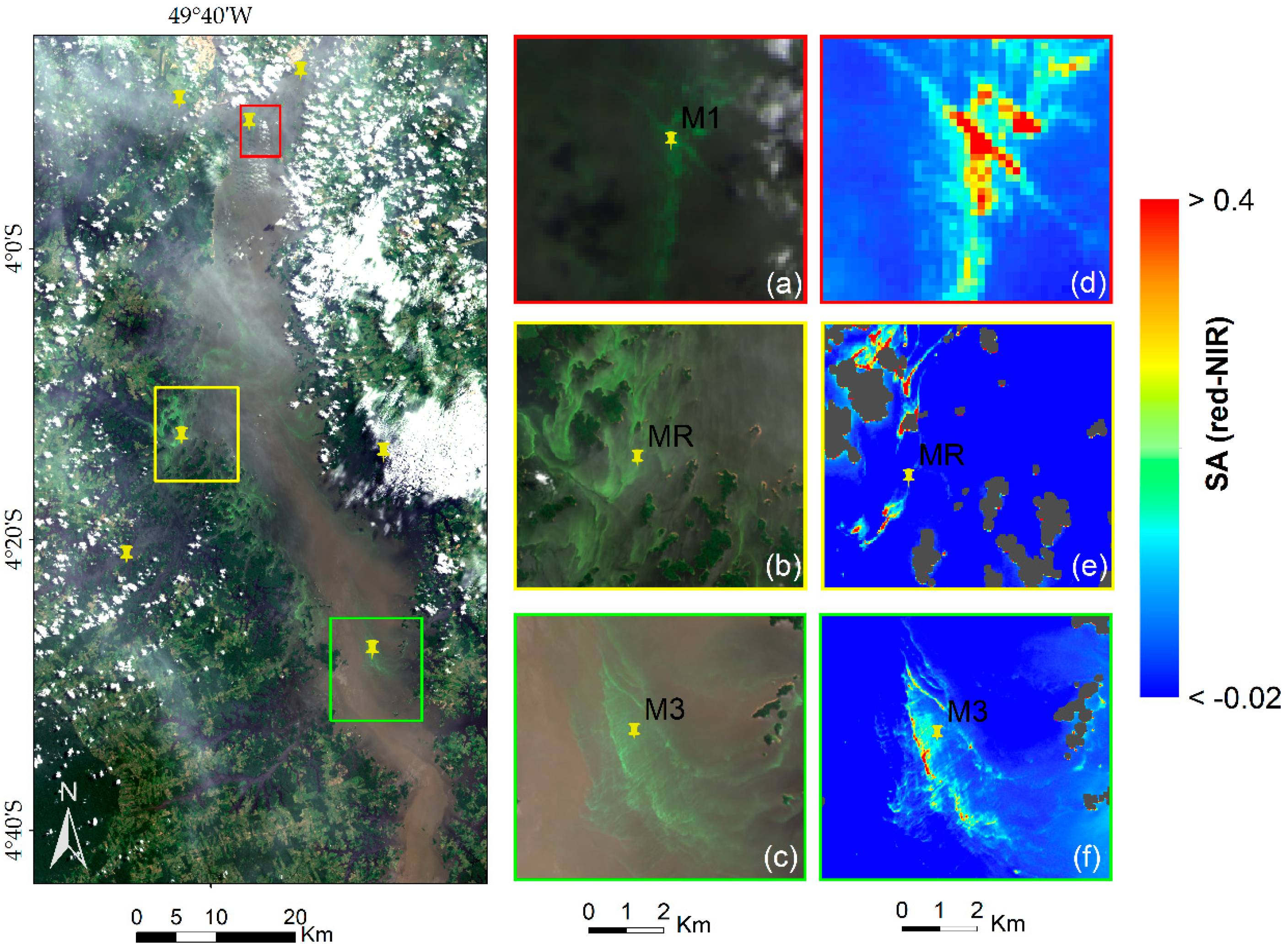
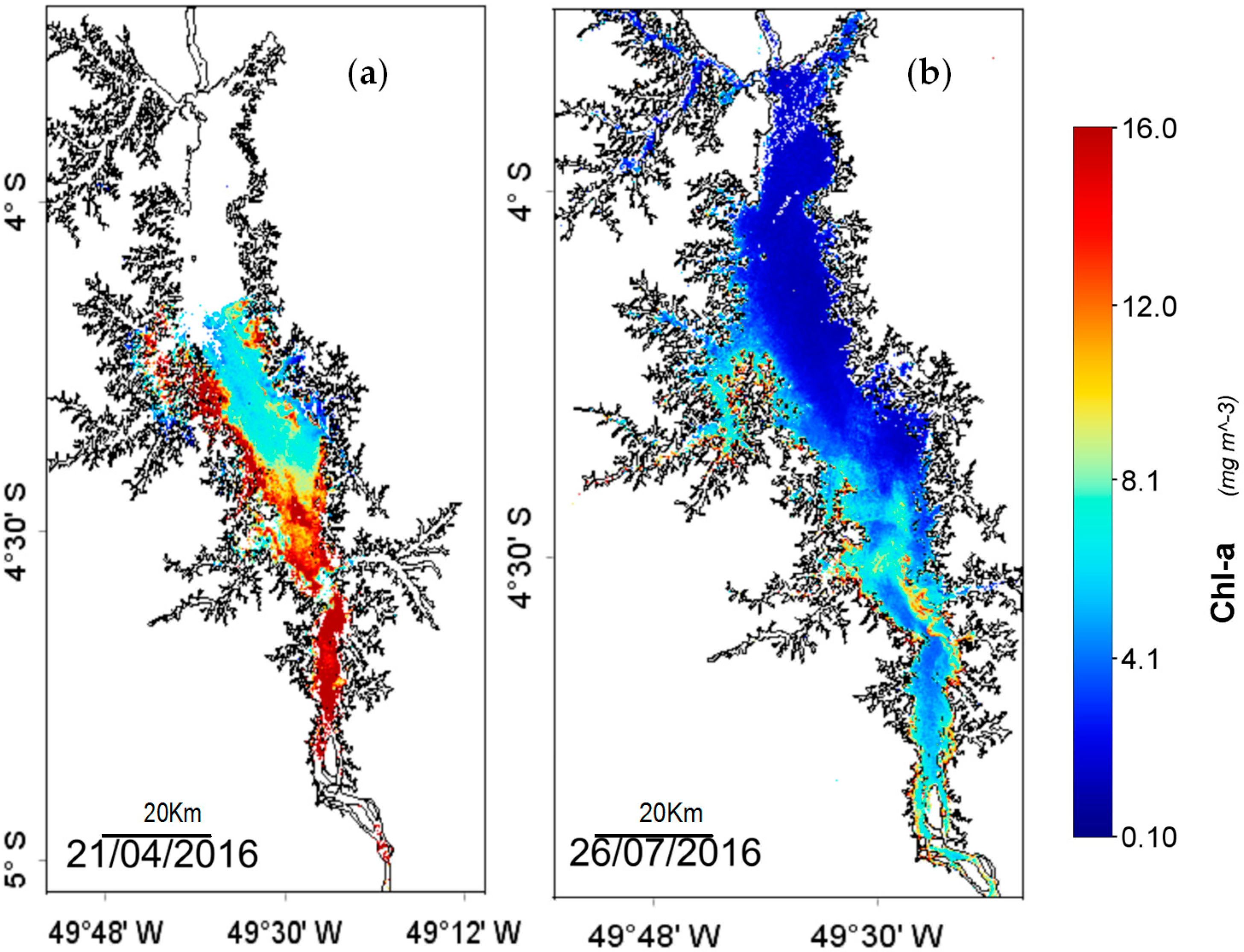
The OBPG algorithm applied to this study area showed to be able to represent the spatial dynamic of the THR in favorable weather conditions for Chl-
a. The SA
red-NIR algorithm could identify the algal bloom extent even in thin cloud conditions (
Figure 4d). The algorithm attached higher values of SA
red-NIR to areas with higher nutrients in calm weather conditions, as preferred by cyanobacteria [
11].
We observed a negative correlation between Chl-
a and cyanobacteria density; a result that was also observed by [
37] in earlier years. In developing a trophic state index for tropical/subtropical reservoirs, De Souza Cunha et al. [
69] noted poor correlation between Chl-
a concentration and phytoplankton abundance. The main reason for such a poor correlation between Chl-
a and phytoplankton abundance is a shift occurring between phytoplankton groups, which contributes to the total Chlorophyll-
a content. This shift was reported by [
70] studying algae in the Arctic, where authors reported an increase in small phytoplankton cells over larger cells, while Chl-
a concentrations did not change.
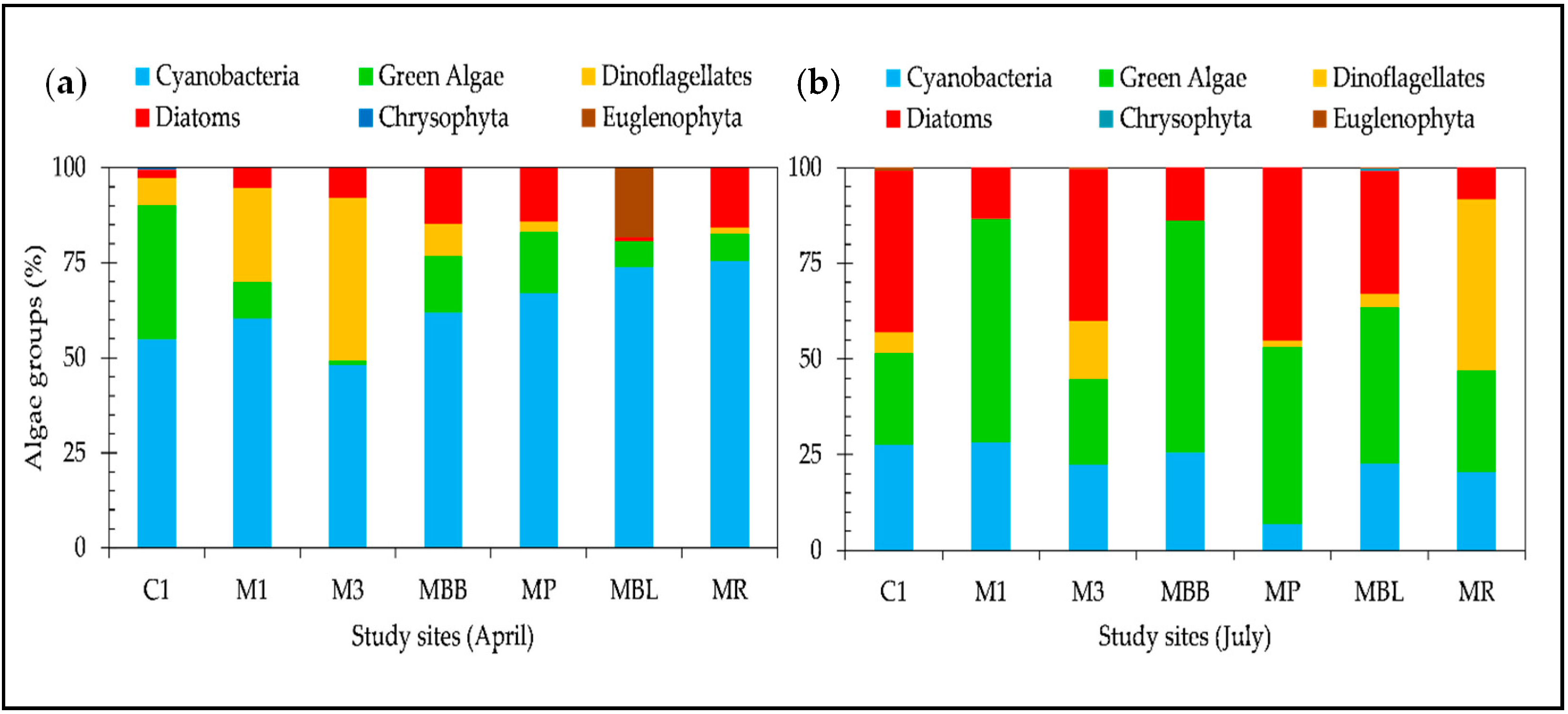
We also observed an increase in phytoplankton abundance in the lake zone (sites M1, C1 and BB), although the Chl-
a concentration did not show significant variation between sites. Kasprzak et al. [
71] reported shifts within the phytoplankton community related to the nutrient load during eutrophication periods. They reported a replacement of small flagellates by green algae. This may be caused by different requirements among phytoplankton species for nutrients. Due to this shifting in the phytoplankton group, caution is necessary when using Chl-
a as an estimator of phytoplankton biomass [
72]. Dinoflagellates and some cyanobacteria species can actually have relatively low Chlorophyll-
a content per cell biovolume [
73,
74]. Certain cyanobacteria species, such as
Dolichospermum circinalis and
Microcystis aeruginosa, present protoplasm that reflects in the green region; and sometimes these species appear to be dark or brown in color with their gas-filled vesicles scattering light in the blue region [
6].
Our results show an increase in density of the
Nostocales order represented by the
Raphidiopsis curvata species in the cyanobacteria group from April to July of 2016 (see
Figures S1 and S2). We also observed an increase in nutrients such as phosphate between April and July, accompanied by an increase in silicate. Species of the
Nostocales order favor turbid waters with a high phosphor concentration [
75]. They are able to fix atmospheric nitrogen in low, combined inorganic sources, opening up opportunities for them to grow in low Chl-
a waters classified as oligotrophic [
76]. The increase in turbidity in the THR during its emptying phase is due to the increase in diatoms, which could explain the high silicate values in July [
46].
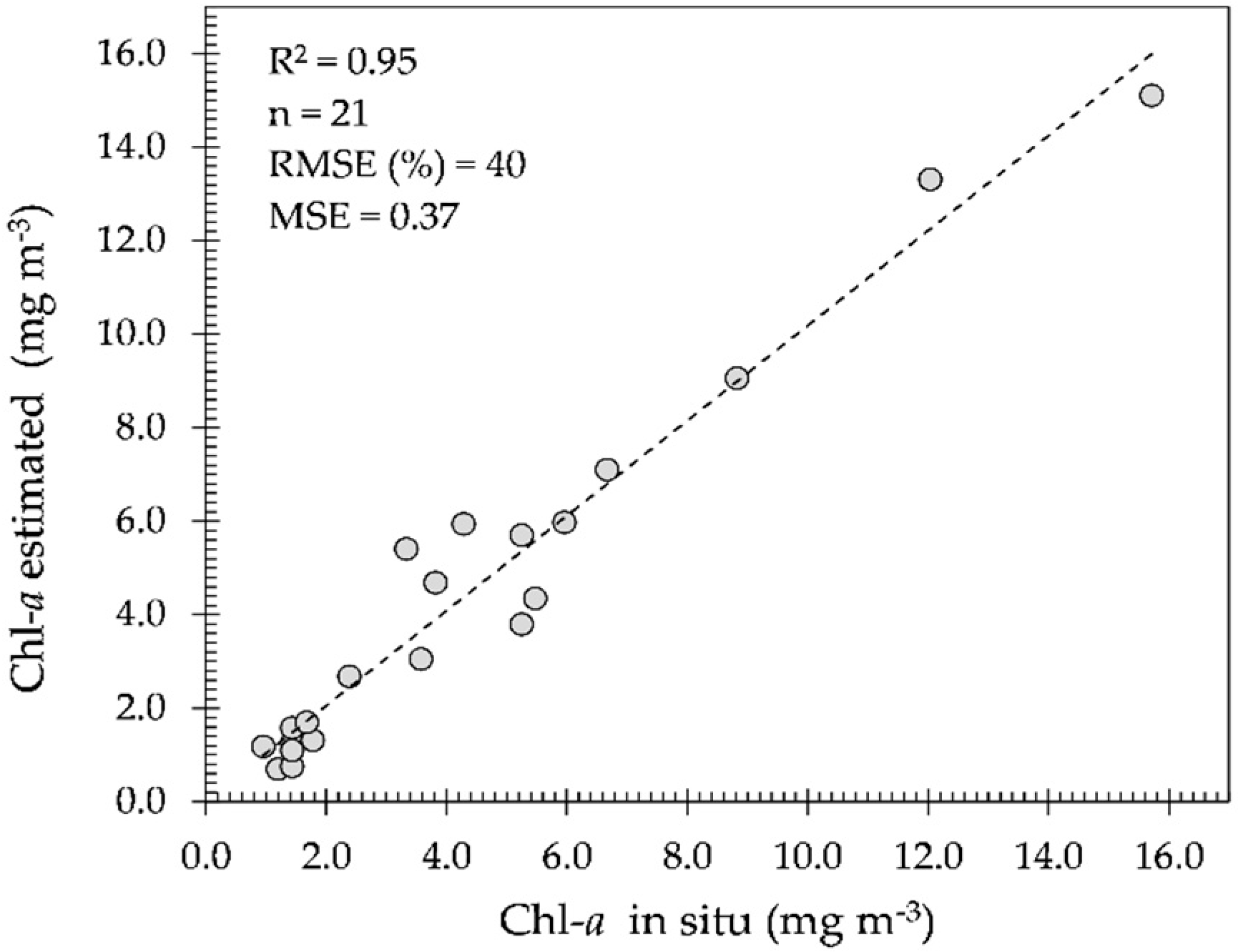
In spite of this study area being located in a cloudy region and investigated in the wet season (
Figure 7a), the OBPG algorithm made a fair estimation of the Chl-
a concentrations during the cloud-free period with low glint effect, corroborated with field measurements as shown in
Figure 6. However, when the glint effect is high (August to September), the estimation becomes very poor. In the
supplementary material, the OBPG algorithm is seen to be applied in cloud-free and low glint conditions (
Figure S1). In this case, it is evident that the THR seasonally alternates between being classified as eutrophic (in April
Figure 7a—full reservoir) and as meso-oligotrophic (in July
Figure 7b;
Figure S1—emptying reservoir).
In April, M1, MR and M3 were classified as severe bloom areas based on the SA
red-NIR algorithm. The main causes were significant variation on pH levels and changes in weather conditions in temporal scale [
2]. The pH level, in many aquatic systems, plays an important role regulating algal abundance and distribution as showed by [
77]. Moreover, a decrease in cyanobacteria under acidified water conditions has been showed by [
78,
79]. We believe that it is very unlikely that the OLI sensor is capable of monitoring algal bloom in July (emptying phase) in this study area on a temporal scale. The main reason being the turbulent environment, which favors diatom biomass increase, consequently increasing turbidity and benefitting species adapted to high turbidity, such as those of the
Nostocales order. Additionally, filamentous cyanobacteria, as the
Notocales, may be abundant but rarely form scums in turbulent waters [
80]. Turbidity increase is due to the increase in phytoplankton biomass. Scum is conductive to high reflectance in the red and NIR bands as outlined by [
66] and it allows for flagging of the algal bloom extent; however, if no scum is formed, algal bloom will not be detectable using the OLI sensor.
5. Conclusions
We used ecological and optical approaches in spatial and temporal contexts to map algal bloom extent in the Tucuruí hydroelectric reservoir (THR). Our main objective was to investigate whether the combination between water limnology and satellite imagery is a suitable approach for monitoring spatial distribution and temporal frequency of algal blooms and establish their potential toxicity in the THR. Despite the fact that the ecological and optical approaches showed both drawbacks and advantages, the overall conclusion is that the OBPG algorithm is suitable for estimating the spatial and temporal variability in Chl-a concentrations. Thus, this algorithm may be applied to this study area, using OLI/L8 imagery in the periods of little cloud cover on a temporal scale and with good understanding of its water limnology.
Interestingly, the OBPG algorithm showed a fair result for this study area, which was not expected due to the use of the blue green ratio. However, the explanation for such a result might be that the oligotrophic to mesotrophic classification between July and September yielded R
rs towards the blue-green region. Additionally, this study area presents low turbidity and color concentrations from July to September as shown in the
supplementary material (Figures S3 and S5).
Despite the limitations of the SAred-NIR algorithm, it showed that it is possible to flag algal bloom occurrence with some a priori knowledge of the study area and availability of limnological and remote sensing data. As the main drawback of this study was using a reduced number of satellite images, we recommend that cyanobacteria or phytoplankton studies in this area ensure that their ecological functioning is carefully considered when attempting to map occurrence using limited satellite imagery. Moreover, the goal of this study was not to quantify algal blooms directly. Instead, this approach meant to search for patterns in space and time based on their ecological preferences for conditions (such as physical, chemical and biological) that would favor algal blooms. Therefore, generated maps based on their probable occurrence would help water management decisions.
In conclusion, further study on the bio-optical properties of Amazonian reservoir waters would be beneficial to local water management in order to understand the water quality issues in these areas.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}