
The Effects of Food Waste on Wildlife and Humans

1
School of Life and Environmental Sciences, Centre for Integrative Ecology, Deakin University (Burwood Campus), Geelong, VIC 3125, Australia
2
School of Life and Environmental Sciences, The University of Sydney, Sydney, NSW 2006, Australia
3
Global Trophic Cascades Program, Department of Forest Ecosystems and Society, Oregon State University, Corvallis, OR 97331, USA
4
School of Environmental and Forest Sciences, University of Washington, Seattle, WA 98195, USA
*
Author to whom correspondence should be addressed.
Sustainability 2017, 9(7), 1269; https://0-doi-org.brum.beds.ac.uk/10.3390/su9071269
Submission received: 28 April 2017
/
Revised: 29 June 2017
/
Accepted: 7 July 2017
/
Published: 19 July 2017
(This article belongs to the Special Issue Reducing Agricultural and Food Waste: Implications for Biodiversity and the Environment)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:A reduction in the loss and waste of human food is a global issue for addressing poverty and hunger in poorer nations, and for reducing the environmental footprint of the agriculture sector. An emerging issue, however, is that food wasted by humans is often accessible to wildlife, affecting wildlife ecology and behaviour, as well as ecological processes and community dynamics. Here we highlight the extent of such impacts, drawing on examples from mammalian predators and other taxonomic groups. We then develop two conceptual models. The first shows how wildlife access to food waste can exacerbate human-wildlife conflicts. The second highlights that when food waste is removed, the effects on wildlife and ecosystem processes should be monitored. The conceptual models are important when considering that large quantities of food waste are intentionally and unintentionally provided to wildlife around the world. We conclude there is an urgent need to change the way people currently manage the food we produce.
1. Introduction
Every year, 1.3 billion tonnes of food produced for human consumption, a third of total production, is lost or wasted [1]. These are shameful statistics given that globally roughly 1 in 8 people are lacking access to sufficient food [2], while land conversion to agriculture is arguably the single greatest threat to global biodiversity [3]. Decreasing food waste while feeding an expanding human population remains an ongoing challenge and has been the subject of extensive research [4]. An emerging issue, however, is that food wasted by humans is often accessible to wildlife [5,6]. For example, unwanted fruit, vegetables, and grain crops may be raided, or animals may access poorly secured food stores. Other forms of food waste include large quantities of carcasses that are rejected or left to rot in fields, and food that is simply dumped in rubbish tips or discarded after harvest. In Africa and Europe for instance, over 10 million tonnes of carcasses of slaughtered cattle are rejected and discarded each year, while in Australia and the USA 3–4 million tonnes of edible food is discarded in rubbish dumps annually [6]. Similarly, 6.8 million tonnes of fish are discarded after capture each year, or 8% of the total global catch [7].
The availability of food waste to wildlife can have dramatic effects on ecological communities and humans. This can manifest when wildlife ecology and behaviour is altered by accessing food waste, and when wildlife access to food waste affects other species and ecological processes, which in turn increases conflict between wildlife and humans [5,6]. Despite knowledge of such impacts, however, there has been no attempt to provide a broad conceptual model showing how these effects manifest and interact with each other. To help fill this knowledge gap, we first provide an overview of the extent to which wildlife utilise food waste. We then summarise how wildlife access to food waste can directly alter wildlife ecology and behaviour, indirectly impact ecological communities, and increase human-wildlife conflicts. We then provide two conceptual models. The first summarises the broad impacts of food waste on wildlife and humans. The second highlights what could potentially happen to wildlife, ecosystem processes, and human-wildlife dynamics when food waste is removed or managed properly. We focus primarily on mammalian predators, but also draw on examples from other taxonomic groups. The results have important implications for understanding the mechanisms by which wildlife are affected by the availability of human-provided foods, and provide impetus to change the way people currently manage food waste.
2. Extent of Use
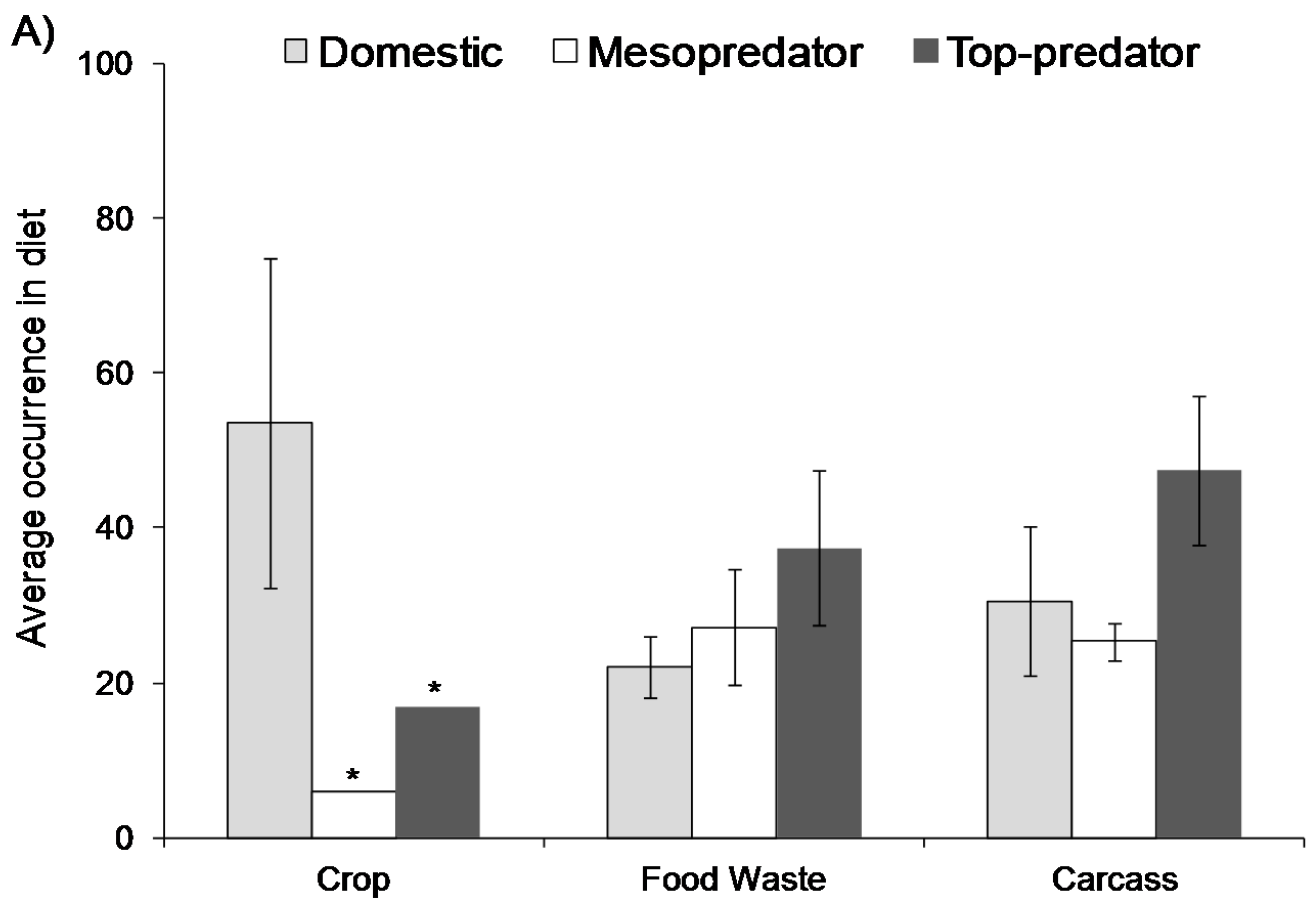
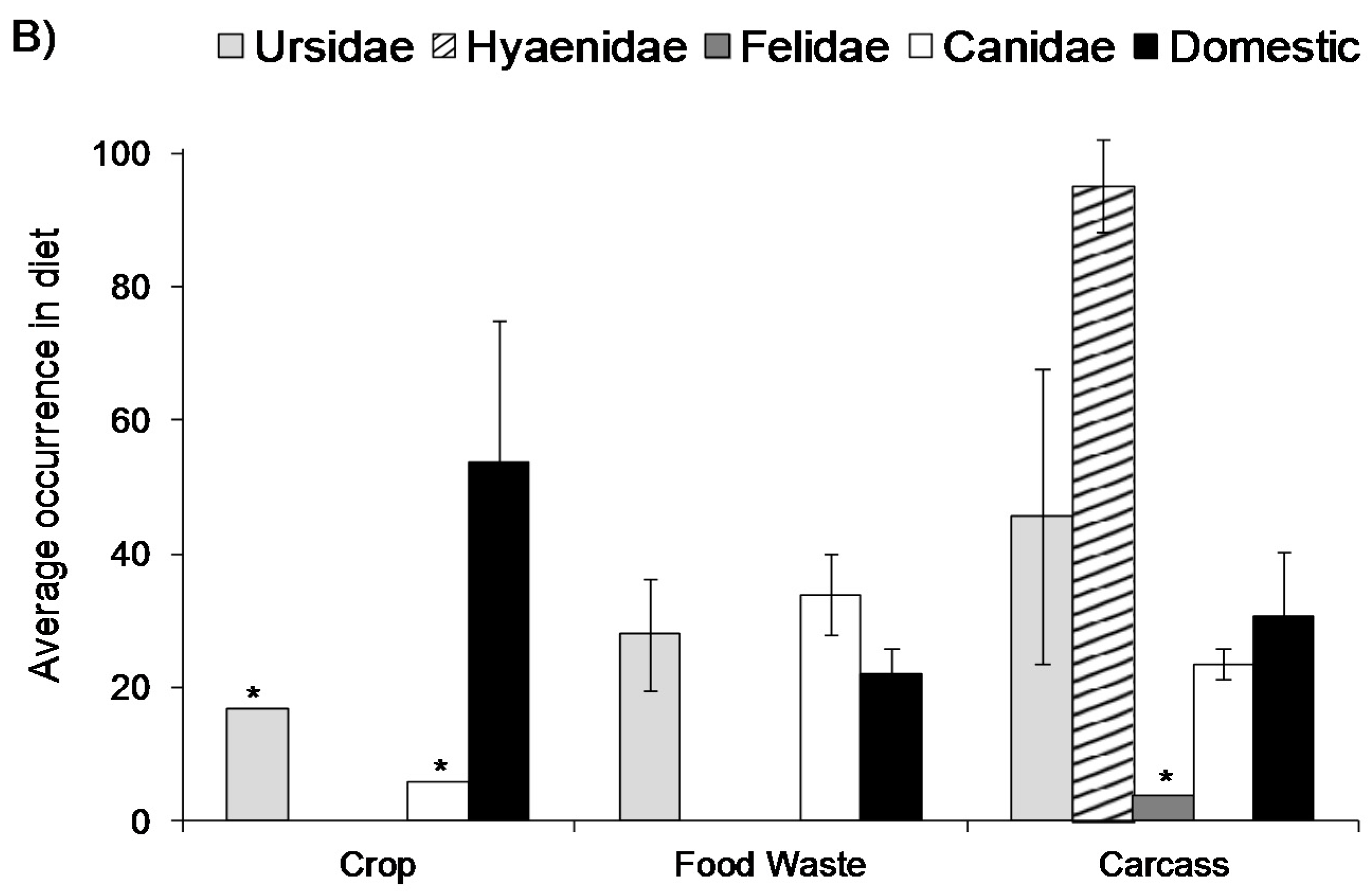
Wildlife may only occasionally utilise food waste, but they can also become completely reliant on this food source. A recent review [5] of human-provided food use by mammalian predators with a body size greater than 1 kg highlights this range of use, including different species groups and the sources of foods they utilise (Figure 1). On average, between 10% and 50% of mammalian predator diets contain food waste, but in some instances it forms >90% [5]. There are several factors that govern the extent to which these species utilise food waste—they may use food waste if wild prey is depleted [8]; the extent of use may relate to dietary preferences, with some predators being more likely to show a preference for some forms of food waste over others [5]; the differential use may reflect habitat quality [9,10], or hunting preferences, with some predators preferring live prey over carcasses or rubbish [5]; skewed use within individual predator species may also reflect social standings, whereby low-ranking individuals and sub-adults become the most frequent users of food waste because they cannot access natural kills, or they avoid kills to reduce intraspecific competition [8]. Alternatively, high-ranking individuals may dominate areas with high food waste which provide high caloric diets [11]. The commonality in all these cases, however, is that mammalian predators can easily become attracted to human-provided foods. The same applies to other taxonomic groups. For example, discards from the fisheries industry in the North Sea alone is estimated to support between 2.5 and 3.5 million sea birds [12].
3. Direct Effects on Wildlife Ecology and Behaviour
In addition to changes in dietary preferences, the distribution and quantity of food waste is likely to influence the carrying capacity, behaviour, and habitat use of wildlife. The Resource Dispersion Hypothesis (RDH) [13] provides an obvious theoretical basis for predicting some of the likely outcomes, at least with respect to territory size and group size. The RDH predicts that the spatial dispersion of food patches determines territory size, whereas patch richness dictates group size. Thus, where there is an abundant food source at a focal location, wildlife may congregate and focus their daily activities around this food source and have larger group sizes. Case studies on Australian dingoes (Canis dingo) and North American bears (Ursus spp.) provide ideal examples to highlight such effects. In the case of dingoes, access to large quantities of food scraps at a waste facility resulted in decreased home-ranges and movements, larger group sizes, increased rates of inbreeding, and changes to their sociality and habitat use [14,15,16,17]. Moreover, the population of subsidised dingoes was a genetically distinct cluster, possibly because of founder effects [15]. In the case of bears, the closure of dumps in Yellowstone National Park in the 1970s led to rapidly increased grizzly bear (Ursus arctos horribilis) mortality and a more than fivefold increase in home range areas [18]. Similar effects have been found for black bears (Ursus americanus), but higher fecundity recorded in urban areas (where they ate food waste) was counter-balanced by high human-caused mortality [19]. High bear mortality by vehicle collisions has been specifically linked with bears’ attraction to garbage [20]. Collision with trains is also a leading cause of grizzly bear mortality in some areas of Canada, where they are attracted to railway lines by grain spilled from freight trains [21]. Such changes to bear mortality are akin to an ecological trap, especially when the overall fitness of a population utilising food waste is lower than those in other available habitats. Thus, while areas containing food waste may provide food for bears, the positive effects may be negated by increased mortality. Changes in bear behaviour and demographics have also been observed in urban areas with food waste, but the extent of such changes are linked to the availability of natural foods, habitat quality, and proximity to preferred habitat patches [9,10,22].
4. Indirect Effects on Other Species and Ecological Communities
The predictability of food waste as a resource can trigger population increases of opportunistic species, in turn altering predator-predator and predator-prey dynamics [5,6]. For example, abundant food supplies can change the interactions between individuals, including bears tolerating other bears around rubbish dumps [23]. The presence of free-ranging domestic dogs in association with garbage dumps determines the presence of other species including maned wolves (Chrysocyon brachyurus), giant anteaters (Myrmecophaga tridactyla), and threatened pudu (Pudu puda) [24,25], with these species either avoiding areas with domestic dogs or being eradicated through predation. The removal of an anthropogenic food source can alter predator-prey dynamics, as in the case of spotted hyenas (Crocuta crocuta) who increased predation on domestic donkeys during Christian fasting periods when food waste was limited [26]. Increasing food waste availability can have a similar effect; for instance, overfishing and increasing waste in landfills have changed the resource base of coastal food webs, and generalist seabirds like western gulls (Larus occidentalis) have responded by shifting their diet to human trash, but at the same time, they increased predation pressure on a threatened species of fish (steelhead, Oncorhynchus mykiss) [27]. More broadly, food provisioning by humans can alter ecosystem functions provided by opportunistic herbivores who act as seed dispersers, as in the case of human food provisioning of macaques (Macaca mulatta) which caused shorter seed dispersal ranges and dispersal into human-modified areas that were not conducive to seed germination [28].
Increased interactions between species at food waste sources may lead to increased hybridisation both among wild species and between wild canids and their domestic relatives [15]. For example, some species of canids can interbreed and produce fertile offspring, including coyotes (Canis latrans), grey wolves (Canis lupus), and domestic dogs (Canis familiaris). The coyote is one species that has thrived in human-modified environments, expanding their distribution and abundance across North America in the absence of the grey wolf [29]. Where grey wolf populations are small and isolated, coyotes and wolves interbreed such that there are now distinct hybrid zones [30]. As in the case of dingoes [15], access to abundant food waste at focal locations could increase contact between species that can interbreed [31]. Similarly, in fragmented human-modified habitats, native species are more likely to come into contact with their domestic relatives. Such hybridisation can result in phenotypic and behavioural changes to offspring [32]. More broadly, hybridisation with domestic relatives is considered a major threat to the conservation of species like Ethiopian wolves (Canis simensis) [33], Australian dingoes [34], and Scottish wildcats (Felis silvestris) [35].
5. Other Impacts to Wildlife Health
Increased interaction between wild and domestic animals can facilitate the spread of disease and pathogens between wild animals, livestock, other domestic animals, and sometimes humans. Increased risk of disease transmission (e.g., rabies, canine distemper virus) due to increased densities of free-ranging domestic dogs could threaten species like African wild dogs and Ethiopian wolves [36,37]. High densities of wildlife can also increase disease occurrence within species, and disease-related mortality has been found to be higher in urban areas for raccoons (Procyon lotor) [38]. In scavenging food waste, animals are at risk of consuming plastic and other non-digestible waste and, while the consumption of plastic by marine wildlife is well-studied [39], it is less so for terrestrial species. In the maned wolf, however, non-digestible anthropogenic waste (e.g., plastic) has been found to comprise 14.1% of all diet items [40] and to be present in up to 40% of scats [41]. Similar studies on dingoes indicate that the probability of a scat sample containing rubbish is relatively high at 17% [14]. Elephants (Elaphus maximus) have been observed feeding at garbage dumps in India [42] and Sri Lanka [43] and, while little is known about the consequences of this behaviour on elephant health, plastic waste is spread via their dung and may then be consumed by other wildlife (T. Thekaekara, The Shola Trust pers comm.).
6. Human-Wildlife Conflict
Wildlife access to food waste can create conflict with humans via indirect and direct feeding. In northern India, abundant sources of human food waste resulted in higher densities of free-ranging domestic dogs who also relied on small livestock as a food source during winter [44]. Predation on livestock by free-ranging dogs can exacerbate human-wildlife conflict where the predator species cannot be identified and native predators are instead blamed [45]. Also, where predation by free-ranging dogs is high, farmers may be less tolerant of predation by protected predators [44]. Extensive use of human-provided foods by wildlife could also lead to increased interactions between humans and wildlife, and to wildlife becoming more habituated to humans. Close encounters between wildlife and humans can stem from direct feeding [46], but also if wildlife encounter humans while seeking food waste. For example, at least one person has reportedly been killed by an elephant that was seeking out a garbage dump [43], and maned wolves feeding on garbage in a Brazilian national park commonly attack camping tents [47]. Such close encounters probably reflect the increased habituation of wildlife towards humans following prolonged periods of access to food waste including direct and/or indirect feeding. Asian lions (Panthera leo persica) in India, for instance, have little fear of humans nowadays, to the extent that tourists can make close observations without a vehicle [48]. This habituation follows periods where humans provided these lions directly with food (in the 1980s, tour guides led goats into the forest and tethered them to stakes for the lions to eat) but this could lead to violent encounters between lions and tourists in the future [31]. Ecotourism and the associated direct feeding of wildlife has led to human fatalities, such as a habituated dingo killing a boy on Fraser Island in Australia in 2001 [49], and habituated grizzly bears killing at least eight people between 1967 and 1986 in several national parks in North America [50].
Anthropogenic food waste as a resource for animal use is typically associated with urban areas, thus creating increased opportunities for animals to impact human livelihoods. Urbanisation is also driving evolution of a range of taxa through habitat modifications, novel disturbances, and social interactions [51] which are largely linked with the provision of anthropogenic food resources. Increases in wildlife or introduced species in urban areas can create a public nuisance. The population of introduced house crows (Corvus splendens) in Singapore increased more than 30 times over 15–16 years, mostly due to their exploitation of anthropogenic food resources [52]. Similarly, rock pigeons (Columba livia) thrive in almost every major city in the world, spreading pathogens to humans and domestic animals and fouling buildings with their excrement to the cost of hundreds of thousands and even millions of pounds per year in the UK [53]. An increased abundance of gulls (Larus spp.) in association with garbage dumps has also been linked with aviation bird strikes [54], which not only cause expensive damage to aircraft but have resulted in 231 human fatalities over an 83-year period [55].
7. Conceptual Models
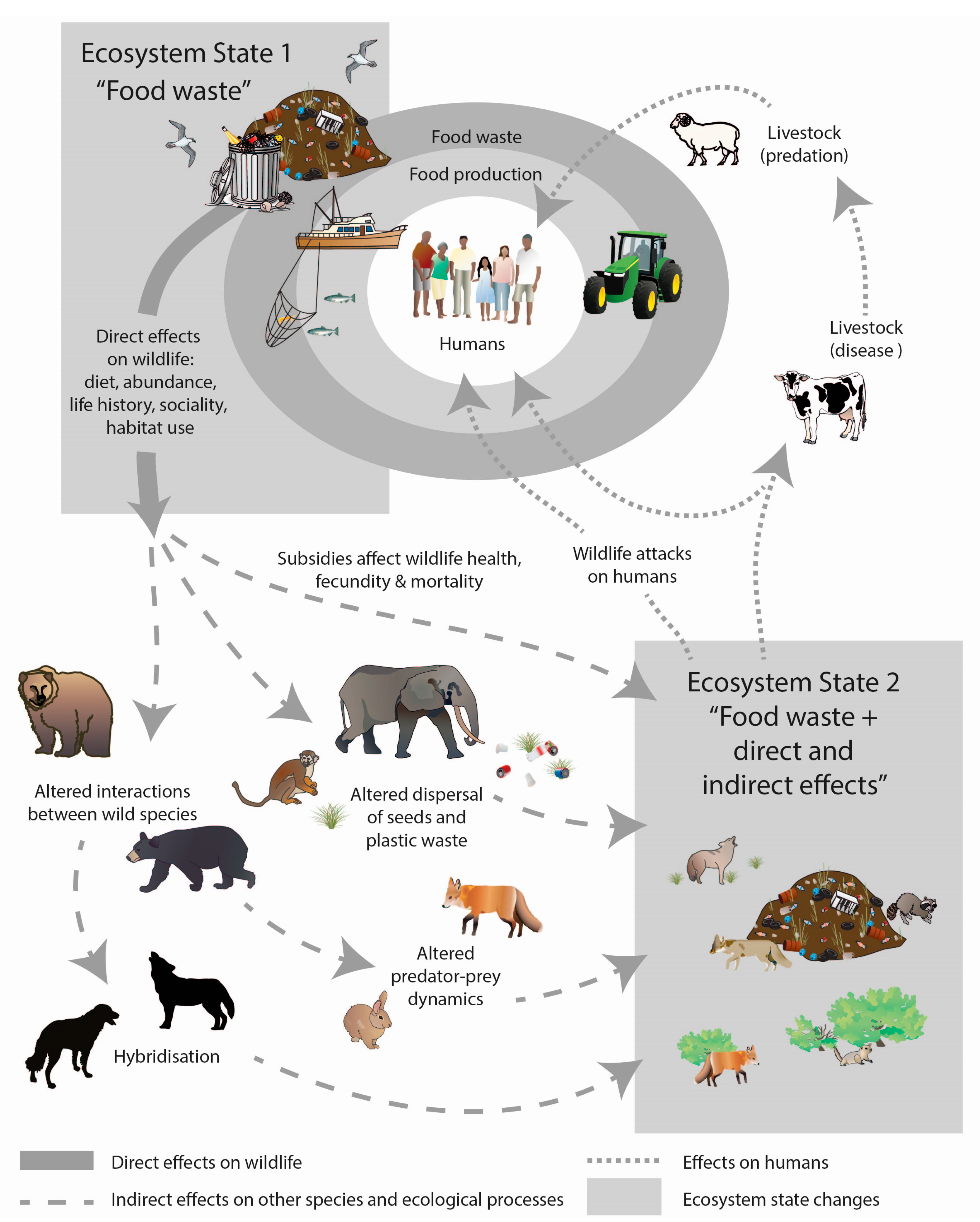
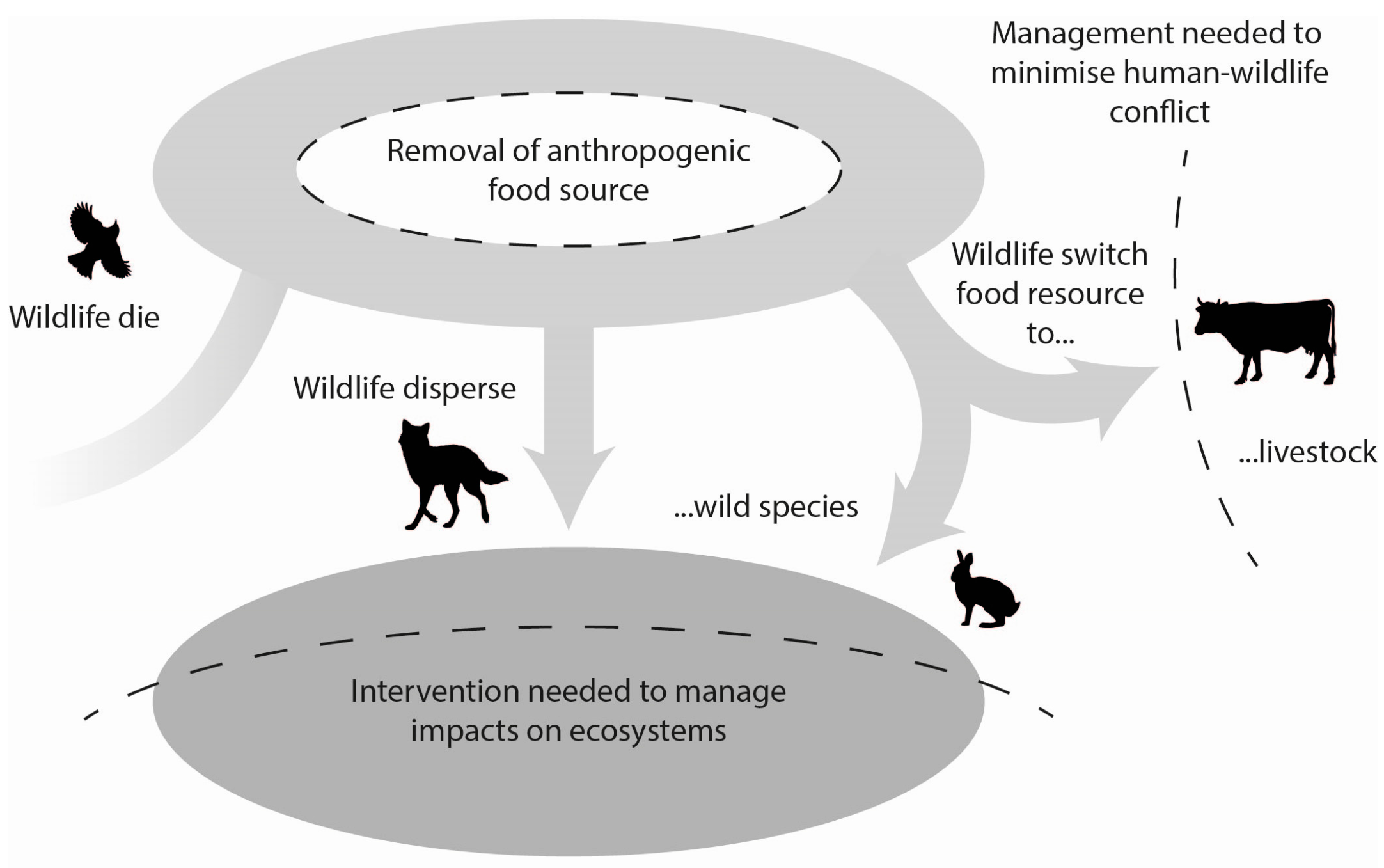
The direct and indirect effects of food waste on wildlife and humans are likely to manifest over multi-generations and across multiple ecosystems. This is depicted in Figure 2, which shows how an ecosystem with food waste (Ecosystem State 1) can directly affect wildlife ecology and behaviour, and indirectly affect other species. Over multiple generations, such impacts are likely to further alter ecosystems and the processes that structure ecological communities (Ecosystem State 2). There may also be impacts on humans and human livelihoods, especially via disease transmission, predation on livestock, and attacks on humans by wildlife. The outstanding question is what happens to wildlife, ecosystem processes, and human-wildlife dynamics when food waste is removed or managed properly by humans. With respect to the wildlife that currently rely on human-provided foods there are several possible outcomes including (1) they die of starvation; (2) they disperse and live elsewhere; (3) they switch their dietary preferences to other prey; and/or (4) they die due to conflict with humans searching for alternative foods. These scenarios are depicted in Figure 3 along with the consequences to ecosystem processes. Studies that track what happens to wildlife when food waste is removed would provide insights into such possibilities. Such an experiment became possible with the closure of dumps in Yellowstone National Park in the 1970s [18]. Experimental manipulation of food waste is also possible [56]. The results of these experiments would help inform the best management strategies to adopt when food waste is removed. It may, for instance, require direct intervention by directly reducing the abundance of some species to protect others (Figure 3). Until such studies are completed, the flow on effects of removing abundant human-provided food sources will be unclear [57]. However, if the food source remains, we do know that the consequences for wildlife, ecosystem processes and human livelihoods are mostly negative (Figure 2), so it may be pertinent to remove and properly manage agricultural and food waste even if the knock-on effects are unclear.
Acknowledgments
This work was not funded by any specific grants.
Author Contributions
Thomas M. Newsome and Lily M. van Eeden conceived and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Food and Agriculture Organization of the United Nations (FAO). Global Food Losses and Food Waste—Extent, Causes and Prevention; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- The World Bank. Food Price Watch, February 2014: Prices Decline at a Slower Pace; Focus on Food Loss and Waste. Available online: http://www.worldbank.org/en/topic/poverty/publication/food-price-watch-february-2014 (accessed on 11 March 2014).
- Machovina, B.; Feeley, K.J. Taking a bite out of biodiversity. Science 2014, 343, 838. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Newsome, T.M.; Dellinger, J.A.; Pavey, C.R.; Ripple, W.J.; Shores, C.R.; Wirsing, A.J.; Dickman, C.R. The ecological effects of providing resource subsidies to predators. Glob. Ecol. Biogeogr. 2015, 24, 1–11. [Google Scholar] [CrossRef]
- Oro, D.; Genovart, M.; Tavecchia, G.; Fowler, M.S.; Martínez-Abraín, A. Ecological and evolutionary implications of food subsidies from humans. Ecol. Lett. 2013, 16, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, K. Discards in the World’s Marine Fisheries. An Update; FAO Fisheries Technical Paper, No. 470; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005. [Google Scholar]
- Kolowski, J.M.; Holekamp, K.E. Effects of an open refuse pit on space use patterns of spotted hyenas. Afr. J. Ecol. 2007, 46, 341–349. [Google Scholar] [CrossRef]
- Johnson, H.E.; Breck, S.W.; Baruch-Mordo, S.; Lewis, D.L.; Lackey, C.W.; Wilson, K.R.; Broderick, J.; Mao, J.S.; Beckmann, J.P. Shifting perceptions of risk and reward: Dynamic selection for human development by black bears in the western United States. Biol. Conserv. 2015, 187, 164–172. [Google Scholar] [CrossRef]
- Baruch-Mordo, S.; Wilson, K.R.; Lewis, D.L.; Broderick, J.; Mao, J.S.; Breck, S.W. Stochasticity in natural forage production affects use of urban areas by black bears: Implications to management of human-bear conflicts. PLoS ONE 2014, 9, e85122. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.P.; Berger, J. Rapid ecological and behavioural changes in carnivores: The responses of black bears (Ursus americanus) to altered food. J. Zool. 2003, 261, 207–212. [Google Scholar] [CrossRef]
- Camphuysen, C.J.; Calvo, B.; Durinck, J.; Ensor, K.; Follestad, A.; Furness, R.W.; Garthe, S.; Leaper, G.; Skov, H.; Tasker, M.L.; et al. Consumption of Discards by Seabirds in the North Sea; Final Report EC DG XIV Research Contract BIOECO/93/10; Netherlands Institute for Sea Research: Texel, The Netherlands, 1995. [Google Scholar]
- Macdonald, D.W. The ecology of carnivore social behaviour. Nature 1983, 301, 379–384. [Google Scholar] [CrossRef]
- Newsome, T.M.; Ballard, G.-A.; Fleming, P.J.S.; Ven, R.; Story, G.L.; Dickman, C.R. Human-resource subsidies alter the dietary preferences of a mammalian top predator. Oecologia 2014, 175, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Newsome, T.M.; Stephens, D.; Ballard, G.-A.; Dickman, C.R.; Fleming, P.J.S. Genetic profile of dingoes (Canis lupus dingo) and free-roaming domestic dogs (C. l. familiaris) in the Tanami Desert, Australia. Wildl. Res. 2013, 40, 196–206. [Google Scholar] [CrossRef]
- Newsome, T.M.; Ballard, G.-A.; Dickman, C.R.; Fleming, P.J.S.; van de Ven, R. Home range, activity and sociality of a top predator, the dingo: A test of the Resource Dispersion Hypothesis. Ecography 2013, 36, 914–925. [Google Scholar] [CrossRef]
- Newsome, T.M.; Ballard, G.-A.; Dickman, C.R.; Fleming, P.J.S.; Howden, C. Anthropogenic resource subsidies determine space use by Australian arid zone dingoes: An improved resource selection modelling approach. PLoS ONE 2013, 8, e63931. [Google Scholar] [CrossRef] [PubMed]
- Craighead, J.J.; Summer, J.S.; Mitchell, J.A. The Grizzly Bears of Yellowstone: Their Ecology in the Yellowstone Ecosystem, 1959–1992; Island Press: Washington, DC, USA, 1995. [Google Scholar]
- Beckmann, J.P.; Lackey, C.W. Carnivores, urban landscapes, and longitudinal studies: A case history of black bears. Hum.-Wildl. Confl. 2008, 2, 168–174. [Google Scholar]
- Huber, D.; Kusak, J.; Frkovic, A. Traffic kills of brown bears in Gorski Kotar, Croatia. Ursus 1995, 10, 167–171. [Google Scholar]
- Gangadharan, A.; Pollock, S.; Gilhooly, P.; Friesen, A.; Dorsey, B.; St. Clair, C.C. Grain spilled from moving trains create a substantial wildlife attractant in protected areas. Anim. Conserv. 2017, in press. [Google Scholar] [CrossRef]
- Lewis, D.L.; Baruch-Mordo, S.; Wilson, K.R.; Breck, S.W.; Mao, J.S.; Broderick, J. Foraging ecology of black bears in urban environments: Guidance for human-bear conflict mitigation. Ecosphere 2015, 6, Article 141. [Google Scholar] [CrossRef]
- Herrero, S. Social behaviour of black bears at a garbage dump in Jasper National Park. In Bears: Their Biology and Management, Proceedings of the 5th International Conference on Bear Research and Management, Madison, WI, USA, 10–13 February 1980; International Association for Bear Research and Management: West Glacier, MT, USA, 1983; Volume 5, pp. 54–70. [Google Scholar]
- Silva-Rodríguez, E.A.; Sieving, K.E. Domestic dogs shape the landscape-scale distribution of a threatened forest ungulate. Biol. Conserv. 2012, 150, 103–110. [Google Scholar] [CrossRef]
- Lacerda, A.C.R.; Tomas, W.M.; Marinho-Filho, J. Domestic dogs as an edge effect in the Brasília National Park, Brazil: Interactions with native mammals. Anim. Conserv. 2009, 12, 477–487. [Google Scholar] [CrossRef]
- Yirga, G.; De Iongh, H.H.; Leirs, H.; Gebrihiwot, K.; Deckers, J.; Bauer, H. Adaptability of large carnivores to changing anthropogenic food sources: Diet change of spotted hyena (Crocuta crocuta) during Christian fasting period in northern Ethiopia. J. Anim. Ecol. 2012, 81, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Osterback, A.-M.K.; Frechette, D.M.; Hayes, S.A.; Shaffer, S.A.; Moore, J.W. Long-term shifts in anthropogenic subsidies to gulls and implications for an imperiled fish. Biol. Conserv. 2015, 191, 606–613. [Google Scholar] [CrossRef]
- Sengupta, A.; McConkey, K.R.; Radhakrishna, S. Primates, provisioning and plants: Impacts of human cultural behaviours on primate ecological functions. PLoS ONE 2015, 10, e0140961. [Google Scholar] [CrossRef] [PubMed]
- Newsome, T.M.; Ripple, W.J. A continental scale trophic cascade from wolves through coyotes to foxes. J. Anim. Ecol. 2015, 84, 49–59. [Google Scholar] [CrossRef] [PubMed]
- VonHoldt, B.M.; Kays, R.; Pollinger, J.P.; Wayne, R.K. Admixture mapping identifies introgressed genomic regions in North American canids. Mol. Ecol. 2016, 25, 2443–2453. [Google Scholar] [CrossRef] [PubMed]
- Newsome, T.M.; Fleming, P.J.S.; Dickman, C.R.; Doherty, T.S.; Ripple, W.J.; Ritchie, E.G.; Wirsing, A.J. Making a new dog? BioScience 2017, 67, 374–381. [Google Scholar] [CrossRef]
- Vilà, C.; Walker, C.; Sundqvist, A.-K.; Flagstad, Ø.; Andersone, Z.; Casulli, A.; Kojola, I.; Valdmann, H.; Halverson, J.; Ellegren, H. Combined use of maternal, paternal and bi-parental genetic markers for the identification of wolf-dog hybrids. Heredity 2003, 90, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gottelli, D.; Sillero-Zubiri, C.; Applebaum, G.D.; Roy, M.S.; Girman, D.J.; Garcia-Moreno, J.; Ostrander, E.A.; Wayne, R.K. Molecular genetics of the most endangered canid: The Ethiopian wolf Canis simensis. Mol. Ecol. 1994, 3, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.; Wilton, A.N.; Fleming, P.J.S.; Berry, O. Death by sex in an Australian icon: A continent-wide survey reveals extensive hybridization between dingoes and domestic dogs. Mol. Ecol. 2015, 24, 5643–5656. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Barratt, E.M.; Gottelli, D.; Kitchener, A.C.; Daniels, M.J.; Pritchard, J.K.; Bruford, M.W. Genetic diversity and introgression in the Scottish wildcat. Mol. Ecol. 2001, 10, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Laurenson, K.; Sillero-Zubiri, C.; Thompson, H.; Shiferaw, F.; Thirgood, S.; Malcolm, J. Disease as a threat to endangered species: Ethiopian wolves, domestic dogs and canine pathogens. Anim. Conserv. 1998, 1, 273–280. [Google Scholar] [CrossRef]
- Woodroffe, R.; Ginsberg, J.R. Conserving the African wild dog Lycaon pictus. I. Diagnosing and treating causes of decline. Oryx 1999, 33, 132–142. [Google Scholar] [CrossRef]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Demographic factors contributing to high raccoon densities in urban landscapes. J. Wildl. Manag. 2003, 67, 324–333. [Google Scholar] [CrossRef]
- Gall, S.C.; Thompson, R.C. The impact of debris on marine life. Mar. Pollut. Bull. 2015, 92, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.A.; Talamoni, S.A. Diet adjustments of maned wolves, Chrysocyon brachyurus (Illiger) (Mammalia, Canidae), subjected to supplemental feeding in a private natural reserve, Southeastern Brazil. Rev. Bras. Zool. 2003, 20, 339–345. [Google Scholar] [CrossRef]
- Aragona, M.; Setz, E.Z.F. Diet of the maned wolf, Chrysocyon brachyurus (Mammalia: Canidae), during wet and dry seasons at Ibitipoca State Park, Brazil. J. Zool. 2001, 254, 131–136. [Google Scholar] [CrossRef]
- Joshi, R. Evaluating the impact of human activities during the Maha–Kumbh 2010 fair on elephants in the Shivalik elephant reserve. Trop. Nat. Hist. 2013, 13, 107–129. [Google Scholar]
- Mendis, R. Elephants Exposed to Rotting Garbage. Available online: http://cea.nsf.ac.lk/bitstream/handle/1/14157/E-2009-Nov-01-Sunday-Leader.pdf?sequence=2&isAllowed=y (accessed on 4 July 2016).
- Home, C.; Pal, R.; Sharma, R.K.; Suryawanshi, K.R.; Bhatnagar, Y.V.; Vanak, A.T. Commensal in conflict: Livestock depredation patterns by free-ranging domestic dogs in the Upper Spiti Landscape, Himachal Pradesh, India. Ambio 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Cozza, K.; Fico, R.; Battistini, M.-L.; Rogers, E. The damage-conservation interface illustrated by predation on domestic livestock in central Italy. Biol. Conserv. 1996, 78, 329–336. [Google Scholar] [CrossRef]
- Orams, M.B. Feeding wildlife as a tourism attraction: A review of issues and impacts. Tour. Manag. 2002, 23, 281–293. [Google Scholar] [CrossRef]
- Almeida Cunha, A. Negative effects of tourism in a Brazilian Atlantic forest National Park. J. Nat. Conserv. 2010, 18, 291–295. [Google Scholar] [CrossRef]
- Packer, C. Is India’s pride actually African? Science 2013, 340, 1289–1290. [Google Scholar] [CrossRef]
- Burns, G.L.; Howard, P. When wildlife tourism goes wrong: A case study of stakeholder and management issues regarding Dingoes on Fraser Island, Australia. Tour. Manag. 2003, 24, 699–712. [Google Scholar] [CrossRef]
- Herrero, S.; Fleck, S. Injury to people inflicted by black, grizzly or polar bears: Recent trends and new insights. In Bears: Their Biology and Management, Proceedings of the 8th International Conference on Bear Research and Management, Victoria, BC, Canada, 20–25 February 1989; International Association for Bear Research and Management: West Glacier, MT, USA, 1990; Volume 8, pp. 25–32. [Google Scholar]
- Alberti, M.; Correa, C.; Marzluff, J.M.; Hendry, A.P.; Palkovacs, E.P.; Gotanda, K.M.; Hunt, V.M.; Apgar, T.M.; Zhou, Y. Global urban signatures of phenotypic change in animal and plant populations. Proc. Natl. Acad. Sci. USA 2017, in press. [Google Scholar] [CrossRef]
- Lim, H.C.; Sodhi, N.S.; Brook, B.W.; Soh, M.C.K. Undesirable aliens: Factors determining the distribution of three invasive bird species in Singapore. J. Trop. Ecol. 2003, 19, 685–695. [Google Scholar] [CrossRef]
- Bevan, R.D.R. The cost of feral pigeon 1990. In British Ornithological Union, Proceedings of a Symposium; ADAS/British Ornithologists’ Union: London, UK, 1990. [Google Scholar]
- Horton, N.; Brough, T.; Rochard, J.B.A. The importance of refuse tips to gulls wintering in an inland area of south-east England. J. Appl. Ecol. 1983, 20, 751–765. [Google Scholar] [CrossRef]
- Thorpe, J. Fatalities and Destroyed Civil Aircraft Due to Bird Strikes, 1912–2002; International Bird Strike Committee: Warsaw, Poland, 2003. [Google Scholar]
- Bino, G.; Dolev, A.; Yosha, D.; Guter, A.; King, R.; Saltz, D.; Kark, S. Abrupt spatial and numerical responses of overabundant foxes to a reduction in anthropogenic resources. J. Appl. Ecol. 2010, 47, 1262–1271. [Google Scholar] [CrossRef]
- Gordon, I.J.; Altwegg, R.; Evans, D.M.; Ewen, J.G.; Johnson, J.A.; Pettorelli, N.; Young, J.K. Reducing agricultural loss and food waste: How will nature fare? Anim. Conserv. 2016, 19, 305–308. [Google Scholar] [CrossRef]
Figure 1.
Average occurrence of food waste in the diet of (A) terrestrial predators > 1 kg in body size split into three functional groups and (B) the same data split into five family groups. The figure is based on data from 83 published studies (±95% confidence intervals) [5]. Frequency of occurrence of prey items in scats was used wherever possible. The asterisk (*) indicates that only one study was available. In (A) domestic species are those that have been purposefully changed by genetic selection by humans (e.g., domestic dogs). Mesopredators are species that suffer from top-down effects (e.g., coyote), whereas top predators are species that suffer little from top-down effects upon reaching maturity (e.g., grey wolf).
Figure 1.
Average occurrence of food waste in the diet of (A) terrestrial predators > 1 kg in body size split into three functional groups and (B) the same data split into five family groups. The figure is based on data from 83 published studies (±95% confidence intervals) [5]. Frequency of occurrence of prey items in scats was used wherever possible. The asterisk (*) indicates that only one study was available. In (A) domestic species are those that have been purposefully changed by genetic selection by humans (e.g., domestic dogs). Mesopredators are species that suffer from top-down effects (e.g., coyote), whereas top predators are species that suffer little from top-down effects upon reaching maturity (e.g., grey wolf).


Figure 2.
Conceptual model showing how wildlife access to food waste can alter wildlife ecology and behaviour, other species, and ecological processes, as well as exacerbate human-wildlife conflicts. The model includes two ecosystem states. In Ecosystem State 1, wildlife use of food waste directly affects wildlife ecology and behaviour (e.g., changes in diet, abundance, life history, sociality, and habitat use). Over multiple generations, these direct impacts are likely to indirectly affect other species and ecological processes, resulting in the formation of Ecosystem State 2. In both ecosystem states there will be impacts on humans and human livelihoods, especially via disease transmission, predation on livestock, and attacks on humans by wildlife.
Figure 2.
Conceptual model showing how wildlife access to food waste can alter wildlife ecology and behaviour, other species, and ecological processes, as well as exacerbate human-wildlife conflicts. The model includes two ecosystem states. In Ecosystem State 1, wildlife use of food waste directly affects wildlife ecology and behaviour (e.g., changes in diet, abundance, life history, sociality, and habitat use). Over multiple generations, these direct impacts are likely to indirectly affect other species and ecological processes, resulting in the formation of Ecosystem State 2. In both ecosystem states there will be impacts on humans and human livelihoods, especially via disease transmission, predation on livestock, and attacks on humans by wildlife.

Figure 3.
Conceptual model showing how the removal of anthropogenic food resources may impact wildlife and ecosystems. For example, wildlife may die of starvation, disperse and live elsewhere, or switch their dietary preferences to other wild prey or livestock, with the latter potentially resulting in increased human-wildlife conflict. Monitoring of such impacts may be necessary when reducing the availability of food waste to wildlife.
Figure 3.
Conceptual model showing how the removal of anthropogenic food resources may impact wildlife and ecosystems. For example, wildlife may die of starvation, disperse and live elsewhere, or switch their dietary preferences to other wild prey or livestock, with the latter potentially resulting in increased human-wildlife conflict. Monitoring of such impacts may be necessary when reducing the availability of food waste to wildlife.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Newsome, T.M.; Van Eeden, L.M. The Effects of Food Waste on Wildlife and Humans. Sustainability 2017, 9, 1269. https://0-doi-org.brum.beds.ac.uk/10.3390/su9071269
AMA Style
Newsome TM, Van Eeden LM. The Effects of Food Waste on Wildlife and Humans. Sustainability. 2017; 9(7):1269. https://0-doi-org.brum.beds.ac.uk/10.3390/su9071269
Chicago/Turabian StyleNewsome, Thomas M., and Lily M. Van Eeden. 2017. "The Effects of Food Waste on Wildlife and Humans" Sustainability 9, no. 7: 1269. https://0-doi-org.brum.beds.ac.uk/10.3390/su9071269
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.