Mapping Invasive Tamarisk (Tamarix): A Comparison of Single-Scene and Time-Series Analyses of Remotely Sensed Data

Abstract
:1. Introduction
2. Methods
2.1. Study Area

2.2. Field Data
2.3. Remotely Sensed Data
2.4. Data Analyses
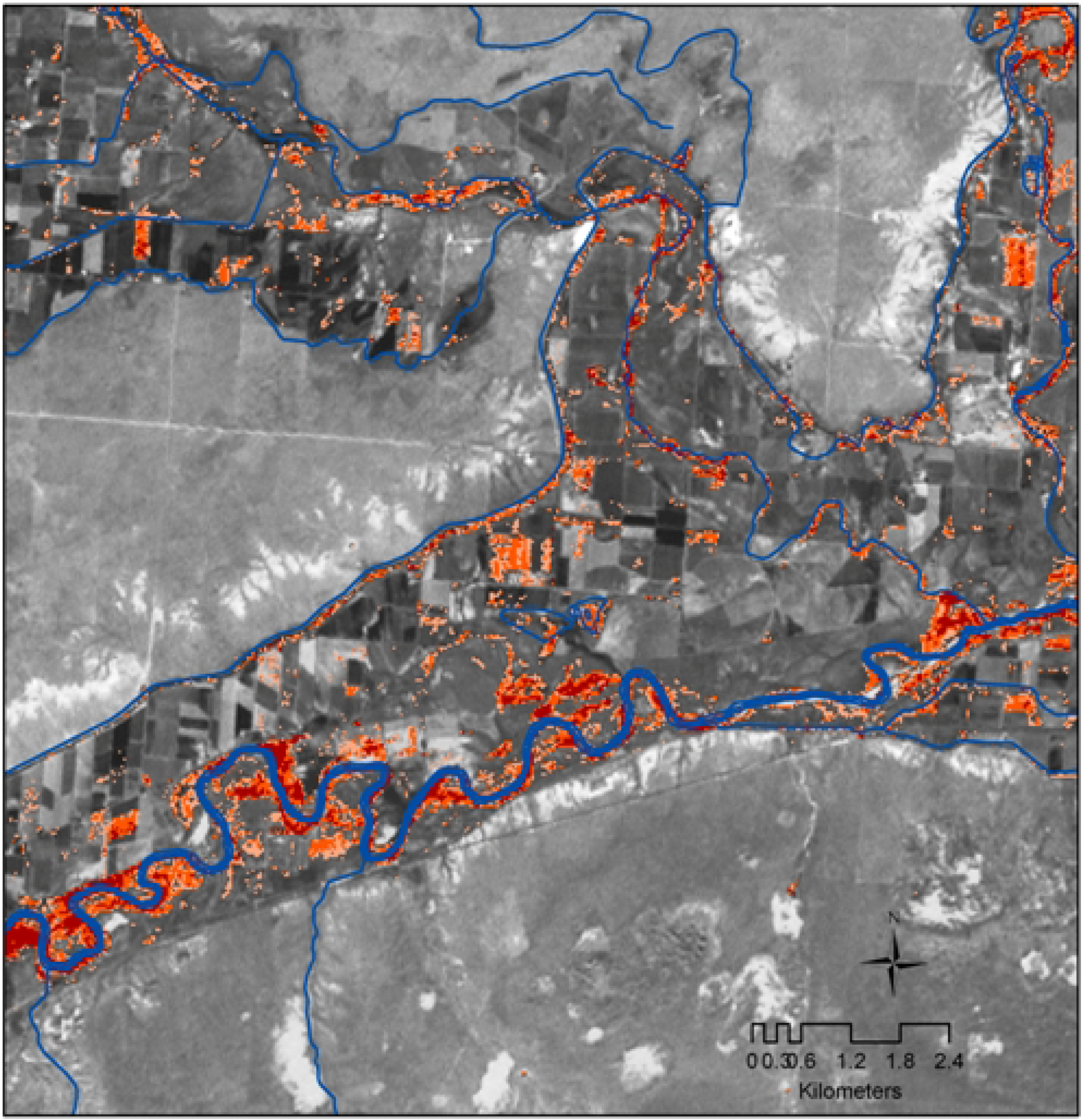
3. Results and Discussion

{kind=link}
{kind=link}
| Scene analysis | Num. variables | AUC | Sensitivity | Specificity | % Correct | Kappa |
|---|---|---|---|---|---|---|
| April | 12 | 0.89 | 0.75 | 0.89 | 0.82 | 0.64 |
| May | 12 | 0.88 | 0.83 | 0.84 | 0.83 | 0.67 |
| June | 12 | 0.92 | 0.93 | 0.76 | 0.84 | 0.69 |
| August | 12 | 0.91 | 0.91 | 0.79 | 0.85 | 0.70 |
| September | 12 | 0.91 | 0.83 | 0.89 | 0.86 | 0.71 |
| October | 12 | 0.89 | 0.77 | 0.94 | 0.85 | 0.71 |
| Time-series1 | 72 | 0.96 | 0.93 | 0.86 | 0.90 | 0.79 |
| Time-series2 | 7 | 0.93 | 0.85 | 0.84 | 0.84 | 0.69 |

| Scene Analysis | Variable | Contribution (%) |
|---|---|---|
| April | band 7 | 25.5 |
| band 4 | 20.6 | |
| NDVI | 17.8 | |
| May | band 7 | 37.7 |
| tasselled cap wetness | 31.9 | |
| band 1 | 9.9 | |
| June | tasselled cap wetness | 78.5 |
| band 1 | 8.6 | |
| band 4 | 5.5 | |
| August | tasselled cap wetness | 59 |
| band 4 | 13.6 | |
| band 1 | 9.6 | |
| September | tasselled cap wetness | 42.2 |
| band 5 | 16.1 | |
| band 7 | 14.3 | |
| October | band 3 | 30.2 |
| tasselled cap wetness | 21.4 | |
| band 7 | 17.9 | |
| Time-series1 | (June) tasselled cap wetness | 25.8 |
| (Sept) tasselled cap wetness | 16.4 | |
| (Oct) band 3 | 11.6 | |
| Time-series2 | (June) tasselled cap wetness | 63.1 |
| (April) NDVI | 9.7 | |
| (Oct) band 3 | 7.8 |
4. Conclusions
Acknowledgements
References and Notes
- Crosier, C.S.; Stohlgren, T.J. Improving biodiversity knowledge with data set synergy: a case study of nonnative plants in Colorado. Weed Technol. 2004, 18, 1441–1444. [Google Scholar] [CrossRef]
- Dewey, S.A.; Anderson, K.A. Distinct roles of surveys, inventories, and monitoring in adaptive weed management. Weed Technol. 2004, 18, 1449–1452. [Google Scholar] [CrossRef]
- Anderson, G.L.; Everitt, J.H.; Richardon, A.J.; Escobar, D.E. Using satellite data to map false broomweed (Ericameria austrotexana) infestations on south Texas rangelands. Weed Technol. 1993, 7, 865–871. [Google Scholar]
- Everitt, J.H.; Anderson, G.L.; Escobar, D.E.; Davis, M.R.; Spencer, N.R.; Andrascik, R.J. Use of remote sensing for detecting and mapping leafy spurge (Euphorbia esula). Weed Technol. 1995, 9, 599–609. [Google Scholar]
- Rowlinson, L.C.; Summerton, M.; Ahmed, F. Comparison of remote sensing data sources and techniques for identifying and classifying alien invasive vegetation in riparian zones. Water SA 1999, 25, 497–500. [Google Scholar]
- Medlin, C.R.; Shaw, D.R.; Gerard, P.D.; LaMastus, F.E. Using remote sensing to detect weed infestations in Glycine max. Weed Sci. 2000, 48, 393–398. [Google Scholar] [CrossRef]
- Lopez-Granados, F.; Garcia-Torres, L. Using remote sensing for identification of late-season grass weed patches in wheat. Weed Sci. 2006, 54, 346–353. [Google Scholar]
- Lass, W.L.; Prather, T.S.; Glenn, N.F.; Weber, K.T.; Mundt, J.T.; Pettingill, J. A review of remote sensing of invasive weeds and example of the early detection of spotted knapweed (Centaurea maculosa) and babysbreath (Gypsophila paniculata) with a hyperspectral sensor. Weed Sci. 2005, 53, 242–251. [Google Scholar] [CrossRef]
- Laba, M.; Tsai, F.; Ogurcak, D.; Smith, S.; Richmond, M.E. Field determination of optimal dates for the discrimination of invasive wetland plant species using derivative spectral analysis. Photogramm. Eng. Remote Sensing 2005, 71, 603–611. [Google Scholar] [CrossRef]
- Carleton, M.A. Adaptation of the tamarisk for dry lands. Science 1914, 39, 692–694. [Google Scholar] [CrossRef] [PubMed]
- DiTomaso, J.M. Impact, biology, and ecology of saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol. 1998, 12, 326–336. [Google Scholar]
- U.S. Department of Agriculture (USDA) Plants Database; Natural Resources Conservation Service. 2008. Available online: http://plants.usda.gov/ (accessed on August 28, 2009).
- Christiansen, E.M. The rate of naturalization of Tamarix in Utah. Am. Midl. Nat. 1962, 68, 51–57. [Google Scholar] [CrossRef]
- Robinson, T.W. Introduction, spread, and aerial extent of saldcedar (Tamarix) in the western states. In Professional Paper 491-A; U.S. Department of the Interior, Geological Survey: Washington, DC, USA, 1965; p. 11. [Google Scholar]
- Harris, D.R. Recent plant invasions in the arid and semi-arid Southwest of the United States. Ann. Assn. Amer. Geogr. 1966, 56, 408–422. [Google Scholar] [CrossRef]
- Lass, L.W.; Thill, D.C.; Shafii, B.; Prather, T.S. Detecting spotted knapweed (Centaurea maculosa) with hyperspectral remote sensing technology. Weed Technol. 2002, 16, 535–545. [Google Scholar] [CrossRef]
- Hirano, A.; Madden, M.; Welch, R. Hypersptectral image data for mapping wetland vegetation. Wetlands 2003, 23, 436–448. [Google Scholar] [CrossRef]
- Hamada, Y.; Stow, D.A.; Coulter, L.L.; Jafolla, J.C.; Hendricks, L.W. Detecting Tamarisk species (Tamarix spp.) in riparian habitats of Southern California using high spatial resolution hyperspectral imagery. Remote Sens. Environ. 2007, 109, 237–248. [Google Scholar] [CrossRef]
- Asner, G.P.; Jones, M.O.; Martin, R.E.; Knapp, D.E.; Hughes, R.F. Remote sensing of native and invasive species in Hawaiian forests. Remote Sens. Environ. 2008, 112, 1912–1926. [Google Scholar] [CrossRef]
- Williams, A.P.; Hunt, E.R., Jr. Estimation of leafy spurge cover from hyperspectral imagery using mixture tuned matched filtering. Remote Sens. Environ. 2002, 82, 446–456. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Everitt, J.H.; Ritchie, J.C.; Moran, M.S.; Booth, D.T.; Anderson, G.L.; Clard, P.E.; Seyfried, M.S. Applications and research using remote sensing for rangeland management. Photogramm. Eng. Remote Sensing 2003, 69, 675–693. [Google Scholar] [CrossRef]
- Everitt, J.H.; Deloach, C.J. Remote sensing of Chinese tamarisk (Tamarix chinensis) and associated vegetation. Weed Sci. 1990, 38, 273–278. [Google Scholar]
- Lass, L.W.; Carson, H.W.; Callihan, R.W. Detection of yellow starthistle (Centaurea solstitialis) and common St. Johnswort (Hypericum perforatum) with multispectral digital imagery. Weed Technol. 1996, 10, 466–474. [Google Scholar]
- Lass, L.W.; Callihan, R.H. 1997. The effect of phenological stage on detectability of yellow hawkweed (Hieracium pretense) and oxeye daisy (Chrysanthemum leucanthemum) with remote multispectral digital imagery. Weed Technol. 1997, 11, 248–256. [Google Scholar]
- Peters, A.J.; Reed, B.C.; Eve, M.D.; McDaniel, K.C. Remote sensing of broom snakeweed (Gutierrezia sarothrae) with NOAA-10 spectral image processing. Weed Technol. 1992, 6, 1015–1020. [Google Scholar]
- Bradley, B.A.; Mustard, J.F. Identifying land cover variability distinct from land cover change: cheatgrass in the Great Basin. Remote Sens. Environ. 2005, 94, 204–213. [Google Scholar] [CrossRef]
- Pavri, F.; Aber, J.S. Characterizing wetland landscapes: a spatiotemporal analysis of remotely sensed data at Cheyenne Bottoms, Kansas. Phys. Geogr. 2004, 25, 86–104. [Google Scholar] [CrossRef]
- Robinson, T.P.; van Klinken, R.D.; Metternicht, G. Spatial and temporal rates and patterns of mesquite (Prosopis species) invasion in Western Australia. J. Arid Environ. 2008, 72, 175–188. [Google Scholar] [CrossRef]
- Anderson, G.L.; Carruthers, R.I.; Ge, S.K.; Gong, P. Monitoring of invasive Tamarix distribution and effects of biological control with airborne hyperspectral remote sensing. Int. J. Remote Sens. 2005, 26, 2487–2489. [Google Scholar] [CrossRef]
- Everett, J.H.; Yang, C.; Fletcher, R.S.; Deloach, C.J.; Davis, M.R. Using remote sensing to assess biological control of saltcedar. Southwest. Entomologist 2007, 32, 93–103. [Google Scholar] [CrossRef]
- Peterson, E.B. Estimating cover of an invasive grass (Bromus tectorum) using tobit regression and phenology derived from two dates of Landsat ETM+ data. Int. J. Remote Sens. 2005, 26, 2491–2507. [Google Scholar] [CrossRef]
- Ge, S.; Carruthers, R.; Gong, P.; Herrera, A. Texture analysis for mapping Tamarix parviflora using aerial photographs along Cache Creek, California. Environ. Monit. Assess. 2006, 114, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Akasheh, O.Z.; Neale, C.M.U.; Jayanthi, H. Detailed mapping of riparian vegetation in the middle Rio Grande River using high resolution multi-spectral airborne remote sensing. J. Arid Environ. 2008, 72, 1734–1744. [Google Scholar] [CrossRef]
- Morissette, J.T.; Jarnevich, C.S.; Ullah, A.; Cai, W.; Pedelty, J.A.; Gentle, J.E.; Stohlgren, T.J.; Schnase, J.L. A tamarisk habitat suitability map for the continental United States. Front. Ecol. Environ. 2006, 4, 11–17. [Google Scholar] [CrossRef]
- Evangelista, P.H.; Kumar, S.; Stohlgren, T.J.; Jarnevich, C.S.; Crall, A.W.; Norman, B.; Barnett, D.T. Modelling invasion for a habitat generalist and a specialist plant species. Divers. Distrib. 2008, 14, 808–817. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the 21st International Conference on Machine Learning; ACM Press: New York, NY, USA, 2004; pp. 655–662. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of problematic alien invasive species-the American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Kumar, S.; Spaulding, S.A.; Stohlgren, T.J.; Hermann, K.A.; Schmidt, T.S.; Bahls, L.L. Potential habitat distribution for the freshwater diatom Didymosphenia geminata in the continental US. Front. Ecol. Environ. 2009, 7. [Google Scholar] [CrossRef]
- Kammerer, J.C. Largest Rivers in the United States. 1990. Available online: http://pubs.usgs.gov/of/1987/ofr87-242/ (accessed on August 28, 2009). [Google Scholar]
- Lindauer, I.E. A comparison of the plant communities of the South Platte and Arkansas River drainages in eastern Colorado. Southwest. Naturalist 1983, 28, 249–259. [Google Scholar] [CrossRef]
- Tamarisk Coalition. Riparian Restoration: Assessment of alternative technologies for tamarisk control, biomass reduction and revegetation. 2008. Available online: http://www.tamariskcoalition.org (accessed on August 28, 2009).
- Leica. ERDAS Imagine 9.1; Leica Geosystems Geospatial Imaging, LLC: Atlanta, GA, USA, 1991–2005. Available online: http://www.erdas.com/Leica.
- ESRI. ArcGIS 9.1; ESRI: Redlands, CA, USA, 2004. Available online: http://www.esri.com/index.html (accessed on August 28, 2009).
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of the Third Earth Resources Technology Satellite-1 Symp., Greenbelt, MD, USA, 1974.
- Kriegler, F.J.; Malila, W.A.; Nalepka, W.A.; Richardson, W. Preprocessing transformations and their effects on multispectral recognition. In Proceedings of the Sixth International Symposium on Remote Sensing of Environment; University of Michigan: Ann Arbor, MI, USA, 1969; pp. 97–131. [Google Scholar]
- Myneni, R.B.; Ramakrishna, R.; Nemani, R.; Running, S.W. Estimation of global leaf area index and absorbed par using radiative transfer models. IEEE Geosci. Remote Sens. 1997, 35, 1380–1393. [Google Scholar] [CrossRef]
- Todd, S.W.; Hoffer, R.M.; Milchunas, D.G. Biomass estimation on grazed and ungrazed rangelands using spectral indices. Int. J. Remote Sens. 1998, 19, 427–438. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Liu, W.G.; Gao, W.; Gao, Z.Q.; Wang, X.L.; Slusser, J. An analysis between the biomass of ecosystem of oasis and the vegetation index. In Remote Sensing and Modeling of Ecosystems for Sustainability III; SPIE: Bellingham, WA, USA, 2006; pp. 62982M-1–62982M-7. [Google Scholar]
- Huang, C.; Wylie, B.; Yang, L.; Homer, C.; Zylstra, G. Derivation of a tasselled cap transformation based on Landsat 7 at-satellite reflectance. Int. J. Remote Sens. 2002, 23, 1741–1748. [Google Scholar] [CrossRef]
- Kauth, R.J.; Thomas, G.S. The tasselled cap-a graphic description of the spectral-temporal development of agricultural crops as seen in Landsat. In Proceedings of the Symposium on Machine Processing of Remotely Sensed Data; LARS, Purdue University: West Lafayette, IN, USA, 1976; pp. 41–51. [Google Scholar]
- Cohen, W.B.; Spies, T.A.; Fiorella, M. Estimating the age and structure of forests in a multi-ownership landscape of western Oregon, USA. Int. J. Remote Sens. 1995, 16, 721–746. [Google Scholar] [CrossRef]
- Jin, S.; Sader, S. Comparison of time series tasselled cap wetness and the normalized difference moisture index in detecting forest disturbances. Remote Sens. Environ. 2005, 94, 364–372. [Google Scholar] [CrossRef]
- Huete, A.R. A Soil-Adjusted Vegetation Index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.J.; Anderson, R.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.; Lehmann, A.; Li, J.; Lohmann, L.; Loiselle, B.; Manion, G.; Moritz, C.; Nakamura, M.; Nakazawa, Y.; Overton, J.; Peterson, A.T.; Phillips, S.; Richardson, K.; Scachetti-Pereia, R.; Schapire, R.; Soberon, J.; Williams, S.; Wisz, M.; Zimmermann, N. Novel methods improve prediction of species distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement of nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: prevalence, Kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.C. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Franklin, S.; Jagielko, C.; Lavigne, M. Sensitivity of the Landsat enhanced wetness difference index (EWDI) to temporal resolution. Can. J. Remote Sens. 2005, 31, 149–152. [Google Scholar] [CrossRef]
- Healey, S.; Yang, Z.; Cohen, W.; Pierce, D.J. Application of two regression-based methods to estimate the effects of partial harvest on forest structure using Landsat data. Remote Sens. Environ. 2006, 101, 115–126. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Evangelista, P.H.; Stohlgren, T.J.; Morisette, J.T.; Kumar, S. Mapping Invasive Tamarisk (Tamarix): A Comparison of Single-Scene and Time-Series Analyses of Remotely Sensed Data. Remote Sens. 2009, 1, 519-533. https://0-doi-org.brum.beds.ac.uk/10.3390/rs1030519
Evangelista PH, Stohlgren TJ, Morisette JT, Kumar S. Mapping Invasive Tamarisk (Tamarix): A Comparison of Single-Scene and Time-Series Analyses of Remotely Sensed Data. Remote Sensing. 2009; 1(3):519-533. https://0-doi-org.brum.beds.ac.uk/10.3390/rs1030519
Chicago/Turabian StyleEvangelista, Paul H., Thomas J. Stohlgren, Jeffrey T. Morisette, and Sunil Kumar. 2009. "Mapping Invasive Tamarisk (Tamarix): A Comparison of Single-Scene and Time-Series Analyses of Remotely Sensed Data" Remote Sensing 1, no. 3: 519-533. https://0-doi-org.brum.beds.ac.uk/10.3390/rs1030519