1. Introduction
The loss and degradation of coastal wetland vegetation due to anthropogenic activities and climatic changes motivate researchers to seek sustainable and efficient management strategies. The ability to understand the dynamics of wetland vegetation is hindered by access limitations due to the risk of damaging habitats and species, fine scale variations of vegetation and hydrology [
1]. Mapping, identification, and classification of plant types and species are vital in planning, restoring, and managing coastal wetlands. Capturing the distribution of alien plant species and particularly controlling the invasive ones is a significant challenge that wetland managers and policy makers face [
2]. Early identification and accurate information about the distribution of invasive species are necessary to anticipate, assess, control, and mitigate their negative impacts on the existing ecosystem health [
3,
4]. These alien invasive plants make an impact on the composition and function of both natural and managed ecosystems with substantial economic cost in response to losing or degrading land use and eradication efforts [
5,
6]. The success of an invasion of alien species depends on the plant’s ability to invade a new region and the susceptibility of the ecological system which is invaded [
7]. Blackburn et al. [
8] dissect the plant invasion process into several stages, namely, transport, introduction, establishment, and spread. During this invasion process, an alien species should pass sequential barriers (e.g., geographical, survival, reproductive, and dispersion) to enter, survive, and spread in a new territory. The invasive species which successfully pass the barriers, compete for space and nutrients in the ecosystem and alter the soil structure and nutrient cycles [
9,
10].
Phragmites australis (later in the text as
Phragmites) is one of the most widespread plants globally and it is seen as a threat to wetlands worldwide [
11].
Phragmites is a tall erect perennial grass that aggressively dispersed over eastern North America during the last two decades [
10,
11,
12,
13]. The
Phragmites haplotype M, which was introduced from Eurasia, has been rapidly replacing its native types and other local plants in most North American wetlands [
14]. They disperse to new areas predominantly by seed germination and spread asexually by stolons or rhizomes around the existing patches [
15,
16]. Dense
Phragmites patches reduce the quality of habitats for fish and bird species, especially due to drying out the littoral zones and affecting sedimentation [
17,
18].
Remote sensing is a widely used technology that is capable of providing spatial and temporal information about invasive species in wetlands [
14]. As remote sensing data analyses become more advanced, data integration methods such as multi-sensor and temporal data fusion become prevalent in enhancing the extraction of information. The tradeoffs among spatial extents, and spatial and spectral resolutions of imagery, affect the quality of information. Several studies fostered hyperspectral data with their continuous spectral band configuration, which provide more details on the spectral characteristics of plants than multispectral imagery [
19]. For example, the Compact Airborne Spectrographic Imager (CASI)-1500 and the Airborne Hyperspectral Scanner (AHS) sensors, used to identify the invasive plant
Spartina densiflora in a wetland, showed promising results using four spectral target detection algorithms [
20]. The hyperspectral imagery of the Airborne Visible/Infrared Imaging Spectrometer (AVIRIS) was found to be capable of mapping invasive plants distributed over large areas with high overall accuracy [
21], although another study suggested that AVIRIS data were not appropriate to map small and highly heterogeneous areas comprised of invasive plants due to the inadequate spatial resolution [
22].
In recent years, many studies have stated the necessity of high spatial resolution imagery to map wetlands to compensate for the spectral similarity among plant types [
22,
23,
24,
25]. The commercially available high-resolution (sub-meter spatial resolution, e.g., WorldView and QuickBird) satellite sensors provide more spatially detailed images with a small geometric distortion [
26]. A classification study conducted to distinguish emergent invasive plants in a diked wetland in the western basin of Lake Erie using QuickBird (2.4 m spatial resolution) images demonstrated the ability of this sensor to distinguish long and narrow patches of invasive plants (
Phragmites australis and
Typha) [
27], while the hyperspectral Hyperion imagery (30 m spectral resolution) was not successful in identifying the small and linear arrangements of
Phragmites australis in the west coast of the Green Bay shoreline [
28]. Distribution maps of the invasive plant
Hakea sericea monitored with WorldView 2 images showed a high overall accuracy [
29] although the maps were not suitable to detect
Hakea sericea at early stages of invasion due to the insufficient spatial resolution of the images. A major drawback of using a commercially available high-resolution satellite data is the high cost of the images and pre-ordering process related to data acquisition. In recent years, the use of an unmanned aerial vehicle (UAV) for detection of invasive species is seen as an economical way of obtaining remote sensing images at any desired time.
UAVs can acquire very high spatial resolution data (~10 cm) with a user defined flight plan and flexible revisit time [
30]. In addition, UAVs allow flying at different heights which can be utilized to adjust the spatial resolution of the images [
31]. Consequently, very high spatial resolution imagery captured by UAVs became practical in natural resource management to monitor invasive plant species in several different ecosystems [
31,
32,
33]. Several recent studies [
22,
31,
32,
33,
34,
35,
36] have proved that the use of UAV-borne remote sensing is an effective method to classify vegetation. Pande-Chhetri [
37] used UAV data to classify wetland vegetation with pixel-based and hierarchical object-based classification approaches. The object-based classification with a support vector machine (SVM) classifier resulted in the highest overall accuracy in the study. While searching for the optimum method to discriminate invasive
Lantana camara from the forested landscape, Nipadhkar et al. [
32] found that the object-based classification provided the better visual organization of plant classification and performed satisfactorily. Müllerová et al. [
22] emphasized the importance of the temporal flexibility in data collection with UAVs over Pleiades satellite images in monitoring invasive plants. The best classification accuracy for invasive
Heracleum mantegazzianum (giant hogweed) was reached during the flowering time using the object-based classification approach. This study highlighted the importance of collecting data at the correct time of the growing season. Samiappan et al. [
33] used five band UAV images to map invasive
Phragmites australis in a tidal marsh to identify the impact of features such as normalized difference vegetation index (NDVI), soil adjusted vegetation index (SAVI) and morphological attribute profiles (MAPs). Further, a canopy height model (CHM) generated using the digital terrain model (DTM) and the digital surface model (DSM) derived from UAV data becomes useful among remote sensing researchers [
38,
39,
40]. The use of UAV and light detection and ranging (LiDAR) derived CHM was identified as an important feature to improve the accuracy in vegetation classification [
41].
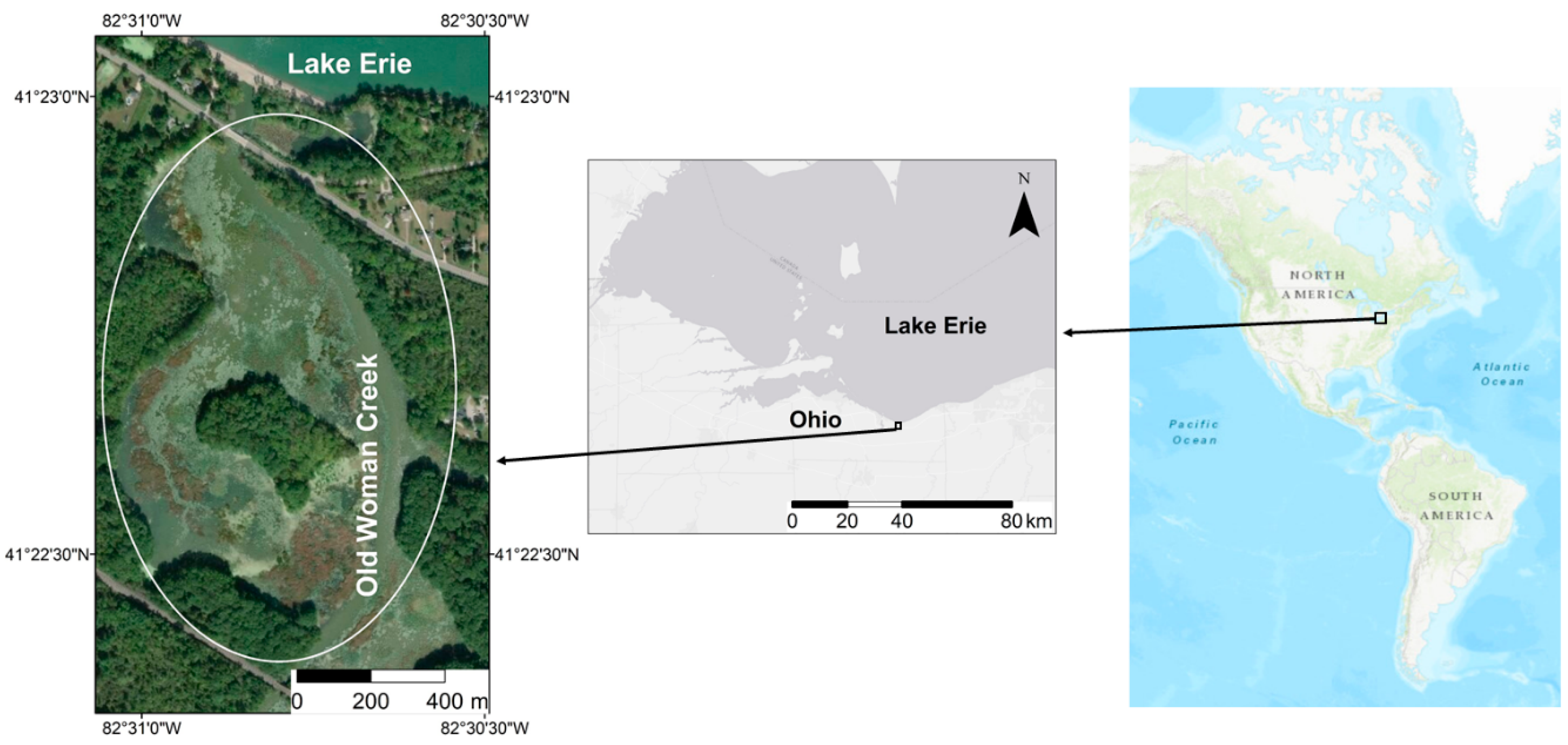
The goal of this study is to explore the effectiveness of UAVs in mapping Phragmites, in the Old Woman Creek (OWC) estuary, located in the Lake Erie region in Ohio, using machine learning (ML) classifiers: k-nearest neighbors (kNN), support vector machine (SVM), and neural network (NN), and their possible advantages over the more traditional approach, maximum likelihood classifier (MLC) using pixel- and object-based classification methods. The objectives of the study are: (i) identify the best machine learning classification algorithm to detect Phragmites as well as to compare it with the parametric MLC; (ii) explore the impact of different feature layers derived from the UAV data on the performance of the classifiers, including various vegetation indices from mid- and late-growing season, image texture, principal components (PC) and the canopy height model (CHM); (iii) assess the optimum use of sample design and cross-validation sampling techniques. The results of this study will be helpful to understand the dispersion of Phragmites in the OWC estuary and to plan the eradication strategies efficiently.
3. Results
The image mosaics created for the study area for August (called 4
sq hereafter) and October using Sequoia camera (
Figure 3a,b) show senescence of some wetland plants in October. This temporal variation is critical to derive NDVI from the October image (NDVI
Oct) and to improve the classification results, as shown below. The RGB image (
Figure 3c) shows more detail and was useful as supplementary data in the ROI collection and validation processes. Most of the shaded areas, especially in the Sequoia images, are removed in the process of masking, as most of the shadows were over water, roads or within trees.
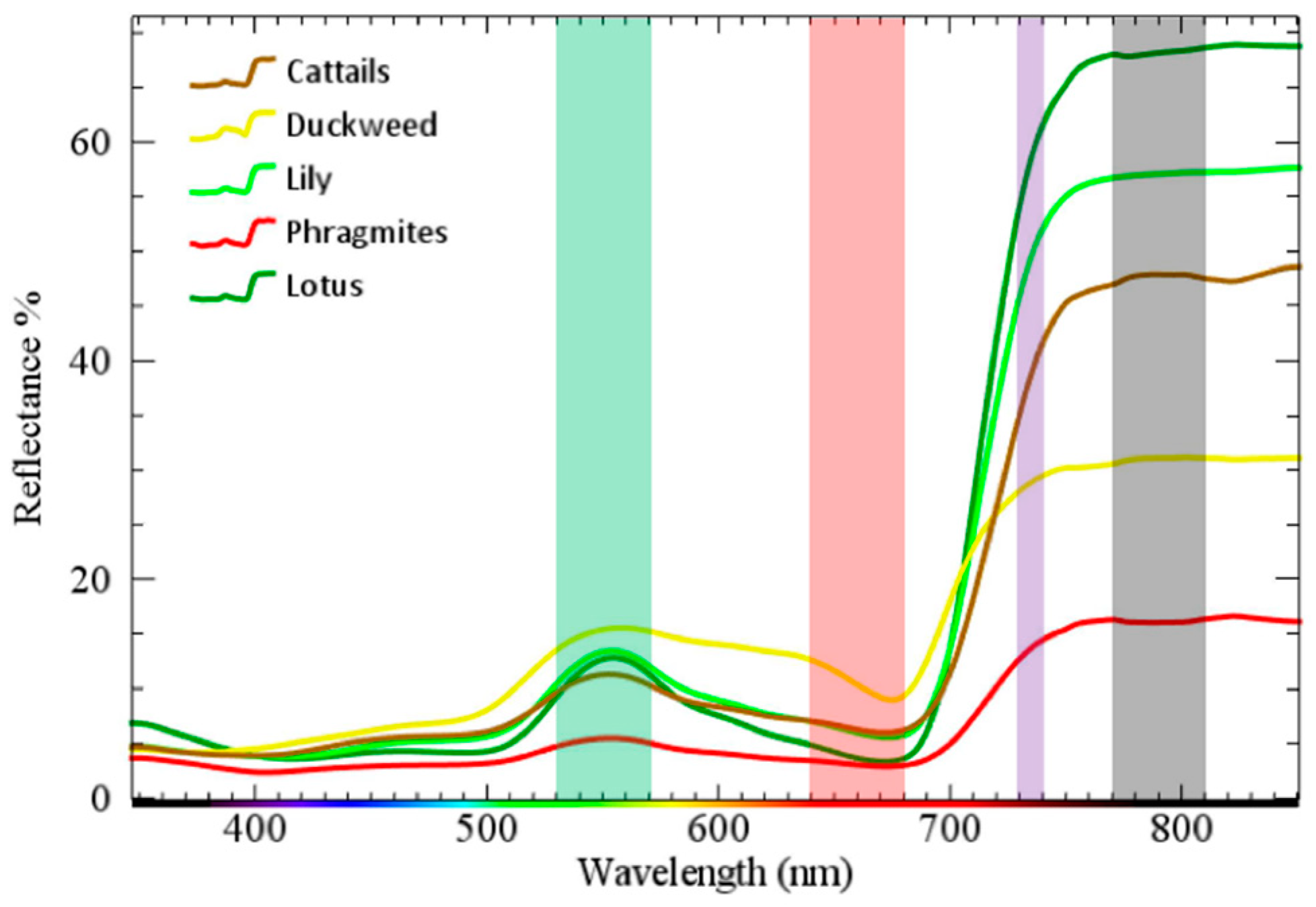
Distinctive differences between the plants’ spectral signatures, collected by the hand-held spectroradiometer, are observed in NIR and RE spectral ranges where lotus exhibits the highest and
Phragmites the lowest values (
Figure 4). In the green spectral region, the spectral signal is similar between cattails, lotus, and lilies, somewhat higher for duckweeds and relatively low for
Phragmites. A similar trend is observed in the red spectral range except for lotus whose reflectance decreases and becomes almost identical to the reflectance of
Phragmites. The
Phragmites signature is relatively distinctive from other plants as it keeps similar values in red and green and the lowest values in NIR/RE regions, which is expected given the earthy color of the plant. In particular, the similarity of the reflectance in the red and the dissimilarity in the NIR bands between
Phragmites and lotus is in accordance with our findings shown below that NDVI index perform best to differentiate
Phragmites and lotus.
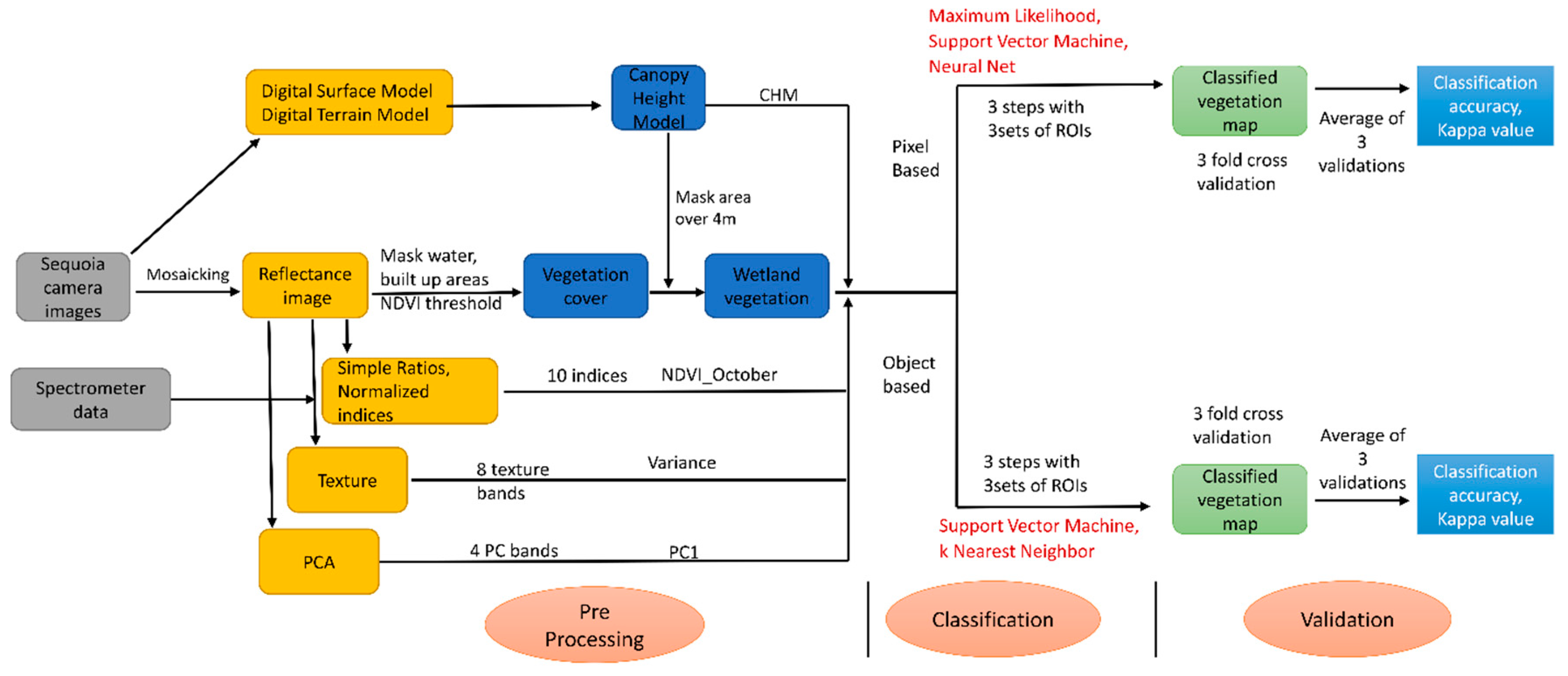
3.1. Pixel-Based Classification
The best results for 4
sq are attained with the SVM classifier (OA = 90.47%) and then with MLC and NN are 88.23% and 84.58%, respectively (
Table 3). While OA reaches relatively high values for all classifiers, the class classification accuracy (i.e., correctly classified percentage per class) is considerably lower for
Phragmites and lotus than other plants, 82.42% and 82.35%, respectively (
Table 3). It is observed from the results that
Phragmites are commonly misclassified as lotus and to a lesser extent as cattails, 15.99% and 1.59%, respectively, and that 1.15% of cattails and 8.89% lotus are classified as
Phragmites. The errors of commission (CE) and omission (OE) are relatively high for
Phragmites (CE = 15.68% and OE = 17.58%, respectively).
Although differences between spectral signatures are observed in
Figure 4, the UAV-derived NDVI values between
Phragmites, cattails, and lotus do not differ significantly from each other in August (
Table 4). Thus, as expected, the NDVI feature layer from August was found not to be beneficial to the classifications and it was excluded from further analysis. The reduction in NDVI from August to October is greater for cattails and lotus compared to
Phragmites as suggested by the Tukey–Kramer test (
Table 4). Nevertheless, NDVI
Oct is statistically different among the plants and it improves the classifications (
Table 5). This seasonal trend was observed for all vegetation indices as shown in
Table 5.
The SVM accuracy parameters for the sensitivity analysis of classification, where the feature layers (band indices, texture, CHM, and PC bands) are stacked to the 4
sq image one by one, are shown in
Table 5. Out of all indices used in the study, NDVI
Oct performs best with the highest overall accuracy of 93.43%, the lowest error of omission of 5.56%, and second lowest error of commission of 4.86%.
Just a slight improvement of the OA values is observed when texture information is added to 4
sq. While the errors of commission are considerably decreased down to 2.08% for several texture layers, the errors of omission are increased for most of them. Variance is the only layer that results in a somewhat, but insignificantly, lower error of omission (OE = 17.24%) when compared to the 4
sq case (OE = 17.58%, see
Table 3), and thus, it was selected as the texture layer with the best contribution to the classification. CHM performs well, resulting in high OA (OA = 93.59%) and relatively low errors of commission and omission (1.75% and 9.53%, respectively). PCs do not contribute considerably to the classification of
Phragmites. PC1 results in somewhat, but not considerably, lower error of omission (OE = 15.52%) than observed in the 4
sq case (OE = 17.58%, see
Table 3).
In summary, the sensitivity analysis suggests that each additional layer to 4sq increases OA (except PC3 and PC4), although in many cases the increase is negligible. The lowest error of omission is reached with NDVIOct; the second lowest error of omission for the Phragmites class is achieved by using the CHM layer; and then negligibly lower by using PC1 and Variance (Var) layers, respectively.
The resulting classification accuracies based on the combination of the selected feature layers for the pixel-based classifiers are shown in
Table 6. Among the three pixel-based classifiers, when both NDVI
Oct and CHM layers are added to 4
sq, the highest overall classification accuracy with the value 94.80% and the highest kappa value of 0.93 are achieved with the NN classifier. The overall accuracy and kappa values decrease when adding more layers (PC1 and Var) to the NN and SVM classifiers. The maximum overall accuracy for MLC is reached for the 4
sq+NDVI
Oct combination (93.46%), and it decreases to 92.92% when CHM is added. The kappa values follow the trend. Any additional layer either decreases the accuracy or produces non-realistic values for MLC (
Table 6). The ML methods do not yield considerably better results than MLC.
Similarly, the class classification accuracies for
Phragmites increase for all classifiers when NDVI
Oct is added and further increase when CHM is included. Maximum values are reached for the 4
sq+NDVI
Oct+CHM combination for all three classifiers. The errors of omission are significantly lowered with NDVI
Oct and further reduced by adding CHM for all classifiers. The lowest error of omission is achieved with the 4
sq+NDVI
Oct+CHM combination using NN (OE = 1.59%). MLC performs well with the lowest error of commission among all classifiers and somewhat higher error of omission than NN (CE = 2.22%; OE = 3.17) for the same (4
sq+NDVI
Oct+CHM) combination (
Table 6;
Figure 5). The additional PC1 and Var layers do not improve the performance of any of the classifiers.
3.2. Object-Based Classification
The segmented images, generated prior to the object-based classification, were classified following the same band combinations. The results of the classifications are shown in
Table 7, for the overall and class accuracy, respectively.
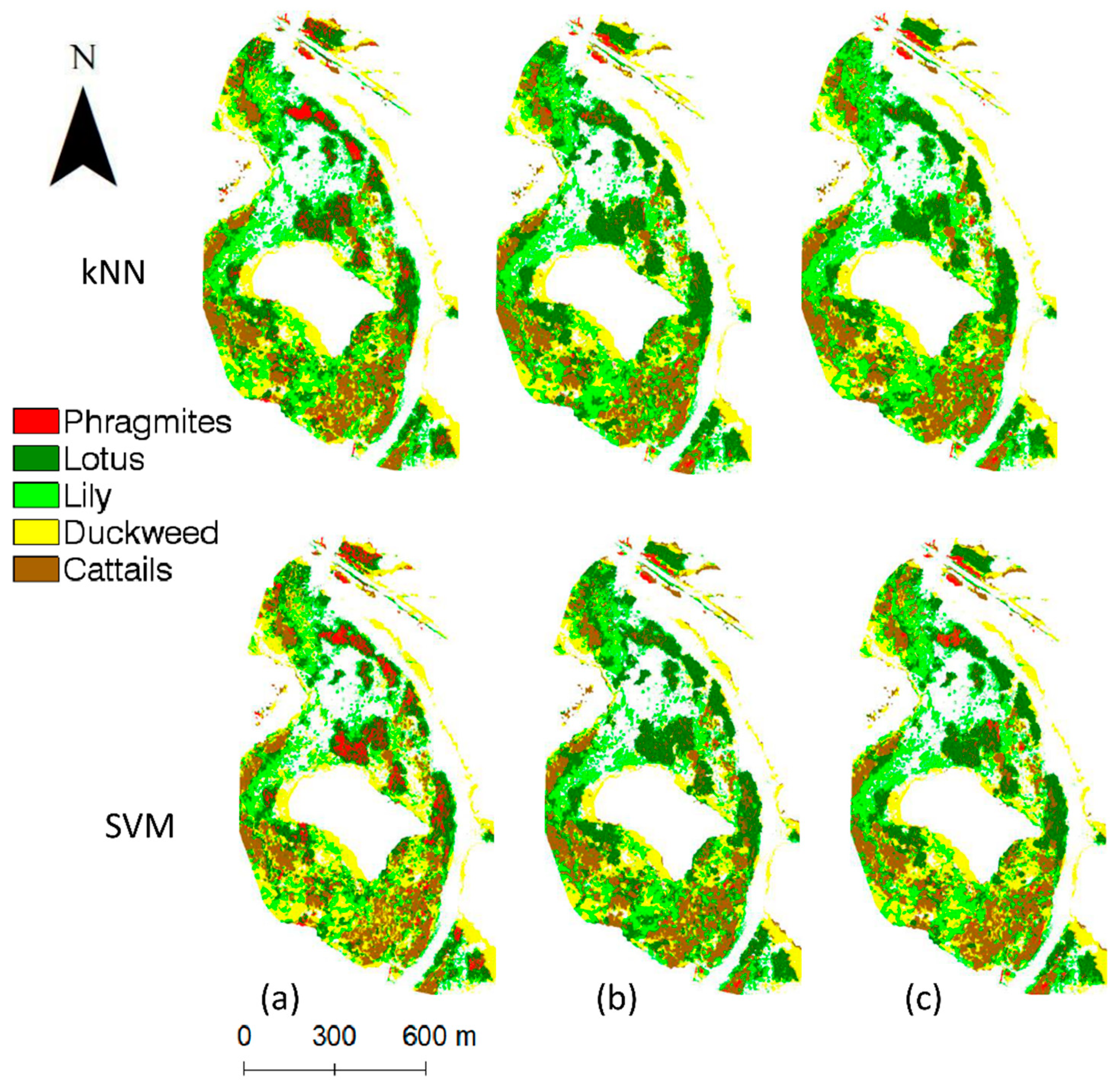
SVM performs slightly better than the kNN for all layer combinations but overall, both classifiers showed lower OA than any of the pixel-based classifiers. The addition of the NDVIOct to 4sq improves the OA values for SVM by approximately 2% (from 87.69% to 89.23%), while the overall accuracy for kNN is reduced by 2% (from 86.92% to 84.62%). Similar to the pixel-based classifications, the highest overall classification accuracy was achieved with SVM for the band combination of 4Sq+NDVIOct+CHM. Although the differences are not significant, the combination of more bands does not improve the overall accuracy and kappa values, with an exception when PC1 is added to kNN.
The combination of NDVI
Oct+CHM layers reduces the error of commission for the
Phragmites class from 28.57% to 3.57% with SVM and from 2.13% to 1.78% with kNN. Similar to the pixel-based classification approach, the minimum errors of omission are observed for the 4
sq+NDVI
Oct+CHM combination for SVM (from 25.00% to 5.32%), and no further reduction is observed by adding the layers (
Table 7). The 4
sq+NDVI
Oct+CHM is the combination that represents the cutoff point where the errors of omission and commission do not improve after adding additional layers. The positive attribute of kNN is that this classifier exhibits lower errors of commission while SVM is more consistent and predictable in reducing the errors of omission (
Table 7). Both classifiers exhibit lower class accuracies and significantly higher errors of omission than the pixel-based approaches.
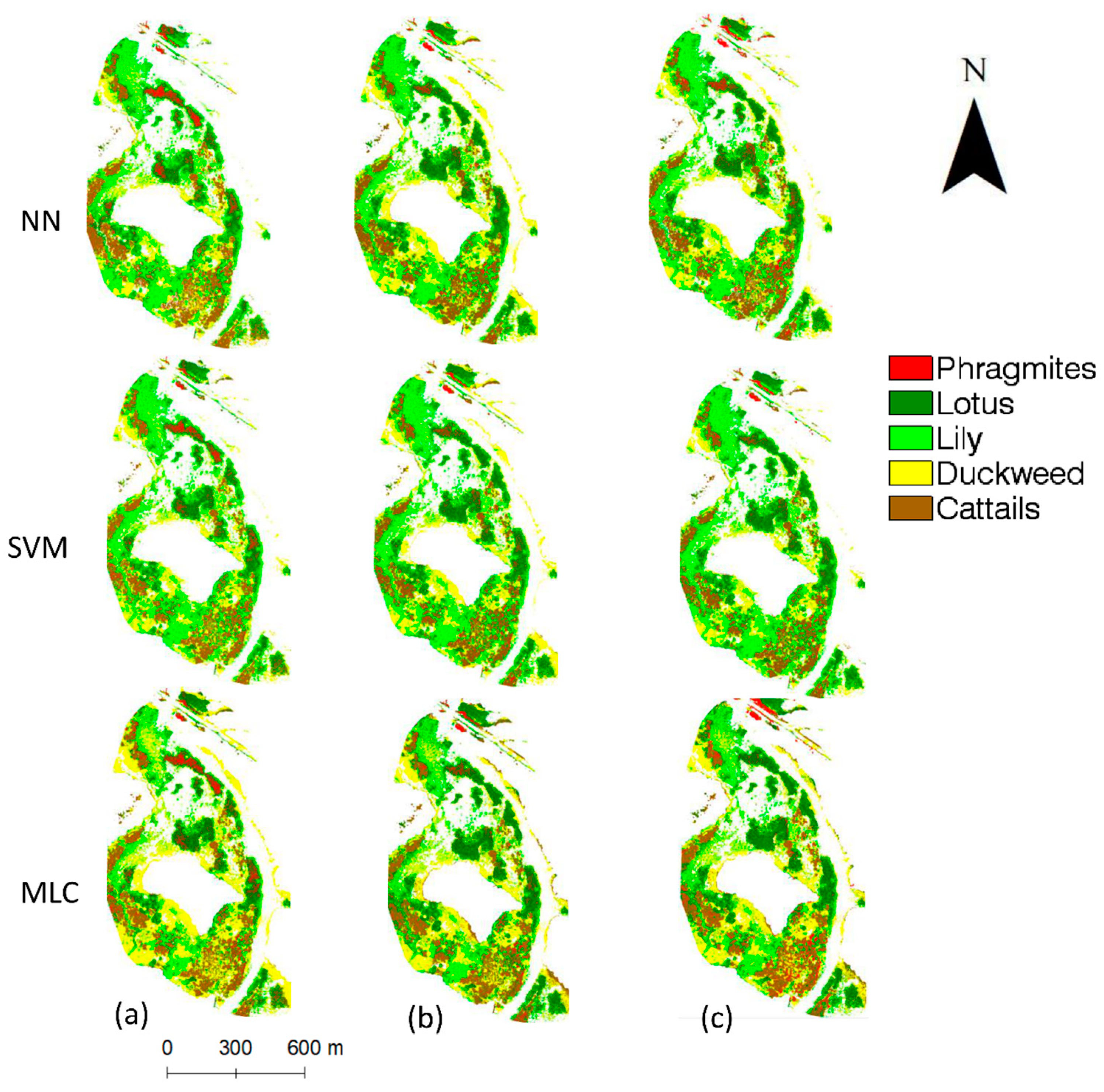
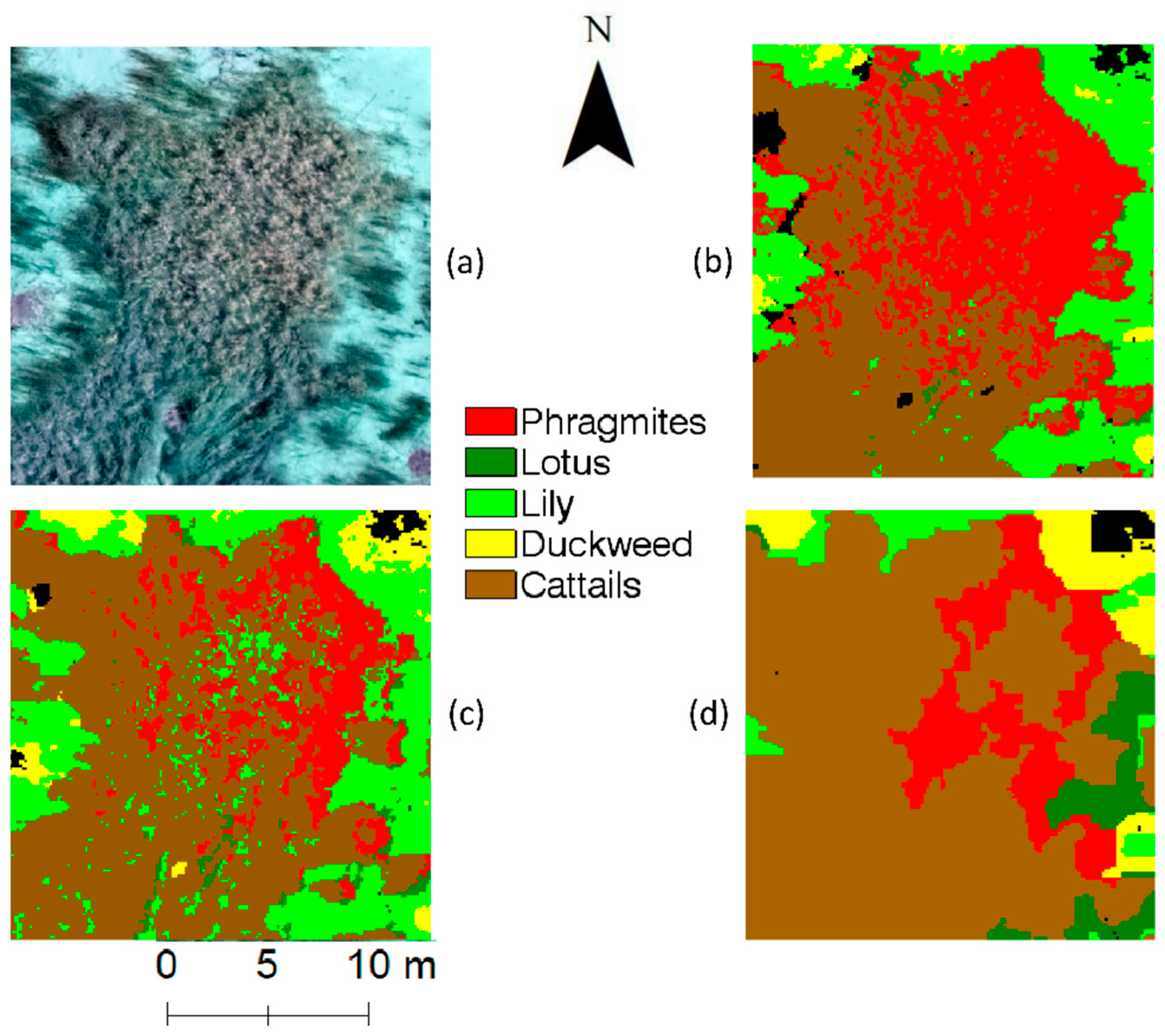
The overall findings in this study can also be supported by visualizing the classified UAV images (
Figure 6). The NN classifier corresponds to the highest classification accuracy of
Phragmites and lowest misclassification of
Phragmites into cattails. Among the selected classifiers in
Figure 6, the object-based SVM shows the lowest
Phragmites classification accuracy due to the misclassification of
Phragmites into the cattails class. Similar visual details can be observed between Phragmites and lotus across the study site.
In comparison, between the pixel-based and object-based classifiers used in this study, there is a noticeable trend that the pixel-based NN classifier demonstrated the best results to detect
Phragmites and the lowest error of omission (
Figure 7). Furthermore, the findings suggest that there are no substantial differences between the ML classifiers and MLC in this study.
4. Discussion
This study demonstrates the advantage of using UAVs to map wetland species, especially to map small patches of invasive Phragmites in the OWC estuary, which would not be possible with coarse airborne and satellite images. This is well demonstrated by high OA for all sophisticated classifiers used in the study, and by relatively low errors of commission and omission for the given combination of feature layers.
Based on careful selection of ROIs, robust sampling design using cross-validation, and careful parameterization of the classifiers, the findings clearly demonstrate that the selection of feature layers is critical. It was clearly demonstrated that the combination of the raw images (4
sq) with feature layers NDVI
Oct, and CHM, derived from UAV, produces the highest overall accuracy (OA = 94.80%) and the lowest error of omission (OE = 1.59%) as well as relatively low error of commission (CE = 4.51%) for
Phragmites when NN is used. It was observed, during the field work, that
Phragmites spread faster in the estuary than other plant types and to improve the eradication of
Phragmites, the lowest omission error is a critical requirement.
Phragmites can be effectively removed by mowing, burning, and applying herbicides during the summer or fall [
77]. The low error of omission for
Phragmites resulted in the study would provide an advantage for estuary management to reach and apply eradicating measures to almost all the
Phragmites patches in the OWC estuary.
4.1. Feature Selection
The study demonstrates that too many features could decrease the classification accuracy, which agrees with the study of Price et al. [
78]. The feature layers should be selected in such a way that they are most optimal in differentiating the classes [
79], and this study has proposed to use NDVI
Oct and CHM to advance the classification of the UAV data. Although the main idea of the study was to concentrate on the August image, the integration of data acquired at different times, in late summer and in mid-fall, shows a clear advantage in this study [
23]. A similar observation was reported by Lantz and Wang [
80] and by several other studies where images at the end of the summer were successfully used to detect invasive
Phragmites [
33,
81,
82]. UAV data acquired in late spring (instead of late-growing season) might be equally useful. This would allow early detection and eradication of
Phragmites and would help to overcome limitations observed in this study where the identification of hidden thin patches of
Phragmites located under dense tree canopies was a challenge. Our efforts to collect UAV data over the study site in late spring were not successful due to frequent disturbances of the UAV flights by eagles nesting in the vicinity.
The importance of using CHM to identify
Phragmites, as it reduces both errors of omission and commission, is also clearly demonstrated, especially when CHM is combined with NDVI
Oct. Several other studies showed the advantages of CHM [
83,
84,
85]. The reduction of errors of omission and commission for
Phragmites was observed in the study of Samiappan et al. [
33], where the author used the SVM classifier. On the other hand, the use of textural and PC measurements does not show a considerable reduction of errors of omission and commission for
Phragmites in this study, which was not the case in several studies [
14,
81,
86] but was the case in the study of Bradly [
22]. A similar trend was observed in both pixel- and object-based classification methods.
4.2. Sampling Design
A good sampling design is vital to achieve high classification accuracy [
81]. A previous study [
78] also highlighted that the complexity of the study site, characteristics of the remote sensing data, image pre-processing methods, and classification approach dictated the sampling design. As described by Stehman [
87], an ideal sampling design should be cost-effective, providing meaningful results to achieve the classification objectives and accommodate any sampling data errors. However, it is not often practical to create a perfect sample design due to the limits of resources and field accessibility [
87]. Foody et al. [
75] studied possible methods to reduce the required training sample size without losing classification accuracy, suggesting that it was possible to use a small dataset when the mechanism of the classifier was known, and when the objective of the study was to map a specific class. However, selecting a sufficient number of training samples becomes a challenge in classification studies if the landscape consists of a smaller number of patches of a particular plant type or if the landscape is complex [
78]. Based on all these principles, after lengthy exploration of various sampling approaches, the results in this study suggest that the stratified random sampling design is necessary for a wetland setup such as OWC. The reduction of the number of training and validation samples for
Phragmites and duckweed classes is not expected to affect classification accuracies as the stratified random sampling design decreases any possible negative effect in this case.
Furthermore, Lucas et al. [
88] claimed that a single pixel is the most suitable sample unit in a raster image for a pixel-based classification. However, if there are limitations such as poor accessibility, which was the case in this study, a sampling unit can consist of multiple pixels with an applied spatial smoothing technique as shown in this study [
88]. A sample should include a sufficient number of basic classification sample units to represent all spectral properties of each class [
69,
77]. The grown ROIs include four or five pixels on average for each sampling point in this study covering higher spectral variability within each class. This approach increases the probability of classifying more pixels into the
Phragmites class. In other words, the method can potentially reduce the errors of omission. The inclusion of more heterogeneous pixels in training and validation data is found to be promising to improve the classification accuracy [
89]. The cross-validation method was applied to compensate for the relatively low, but still sufficient, number of samples per class in this study.
4.3. Object- vs. Pixel-Based Classification and Classification Algorithms
Several studies concluded that object-based classification performs better than pixel-based classification because it creates uniform objects by merging similar pixels into one object [
37,
72,
90]. Specifically, Lantz and Wang [
79] emphasized that the object-based classification method resulted in higher accuracy over pixel-based classification to identify invasive
Phragmites, while Bradley [
23] and Pande-Chhetri et al. [
37] reported the opposite results when identifying some other invasive plants. Interestingly, the current study results in a higher error of omission for the object-based methods, similar to the study of Pande-Chhetri et al. [
37]. Although the best scale parameter for segmentation was carefully selected in this study (as explained above), the segmentation could be slightly erroneous to the extent where
Phragmites pixels are confused with the pixels of cattails due to similar pixel values. This confusion could lead
Phragmites pixels to be aggregated in the segments that include cattails. This situation was observed during the process of segmentation in this study, especially at the boundaries between cattails and
Phragmites. Therefore, the process of segmentation was an initial step where some uncertainties could be generated suggesting that this step is critical as small variability could lead to unexpected results. Edge enhancement techniques could be used to overcome such errors at the boundaries of segments and improve classification accuracy [
91], which was not considered in this study.
Overall, no considerable advantages of ML classifiers over MLC are observed in this study. The similar overall accuracy, lower errors of commission, and the slightly higher errors of omission for MLC praise this parametric method as highly advanced. The findings show that the ML methods are not considerably better for the most optimal layer combination (4
sq + NDVI
Oct + CHM), and that the tradeoffs between the ML classifiers and MLC should be considered given that the parameter optimization of SVM and NN classifiers need more time and effort compared to MLC. The disadvantage encountered with MLC in this study suggests that this classifier does not provide meaningful results when the number of bands increases to more than six. This was also demonstrated in the study of Cheeseman et al. [
92]. In contrast, non-parametric classifiers (NN and SVM) are not considerably affected when the number of feature layers increases. High classification accuracies with the same classifiers were also achieved in the previous studies of Foody and Mathur [
69], Ndehedehe et al. [
65] and Qian et al. [
68]. While some studies suggested that SVM outperformed NN in overall classification accuracy [
66,
89,
93], the finding results in this study suggest that NN is exceptionally efficient when the minimum error of omission is required. Both high classification accuracy and the lowest error of omission for
Phragmites are reached with a NN classifier restricted to one hidden layer [
67]. Higher flexibility of the NN classifier due to the availability of a large number of different synaptic weights between each pair of nodes [
62] has classified more pixels situated at the boundaries between
Phragmites and cattail patches into the
Phragmites, resulting in a lower error of omission.
5. Conclusions
High-resolution images acquired by UAVs are useful in mapping and evaluating wetland invasive plants because of image spatial resolution, ease of handling, and time and cost flexibility. This study used three pixel-based (NN, SVM, and traditional MLC) and two object-based (SVM and kNN) classifiers to detect Phragmites in the Old Woman Creek (OWC) Estuary in Ohio, USA. The UAV image acquired on 8 August 2017 was classified at three different levels of complexity using the proposed classifiers based on: (i) the four original bands (G, R, RE, NIR) (4sq); (ii) the sensitivity analysis where one feature layer at a time was added to 4sq, including information from the October image; and (iii) the combination of representative feature layers with the best performance added to 4sq.
It was clearly demonstrated that the combination of the raw images (4sq) with feature layers NDVIOct and CHM, derived from UAV, produced the highest overall accuracy (OA = 94.80%) and the lowest error of omission (OE = 1.59%) as well as a relatively low error of commission (CE = 4.51%) for Phragmites when NN is used. NN is recognized as the most effective approach in minimizing the error of omission. The findings suggest that the pixel-based classification was advantageous over the object-based approach to identify small patches of Phragmites as found in the OWC estuary.
The study also included a detailed analysis of the sampling design and the number and distribution of ROIs, suggesting that stratified random sampling design with multi-pixels as a sampling unit was the most appropriate method to map small Phragmites patches. It was suggested that the sampling method combined with the cross-validation statistical approach is critical for any of the classifiers used in the study. Temporal variability of NDVI in combination with CHM are the two most important feature layers to reach the best performance of any of the classifiers. Any additional information, such as image texture and PC, was found not to be useful in this study, having a negative or neutral impact when combined with other layers in the process of classification. The findings show that the ML methods are not considerably better than MLC for the most optimal layer combination (4sq + NDVIOct + CHM), and that the tradeoffs between the ML classifiers and MLC should be considered in future studies. The study provides a method to detect invasive Phragmites with high accuracy in a small area using a limited number of samples.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}