Understanding Tree-to-Tree Variations in Stone Pine (Pinus pinea L.) Cone Production Using Terrestrial Laser Scanner

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Region
2.2. Permanent Plots for Monitoring Cone Production
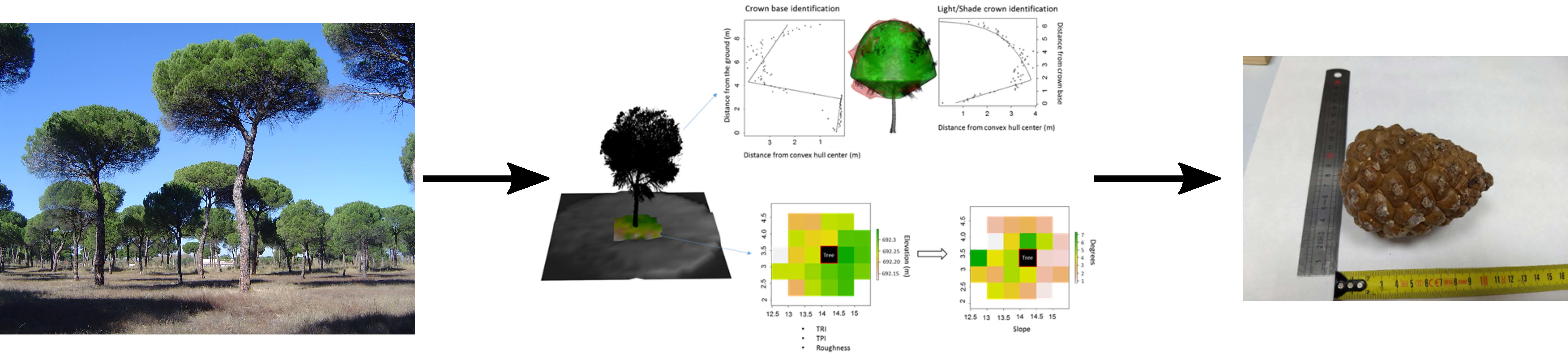
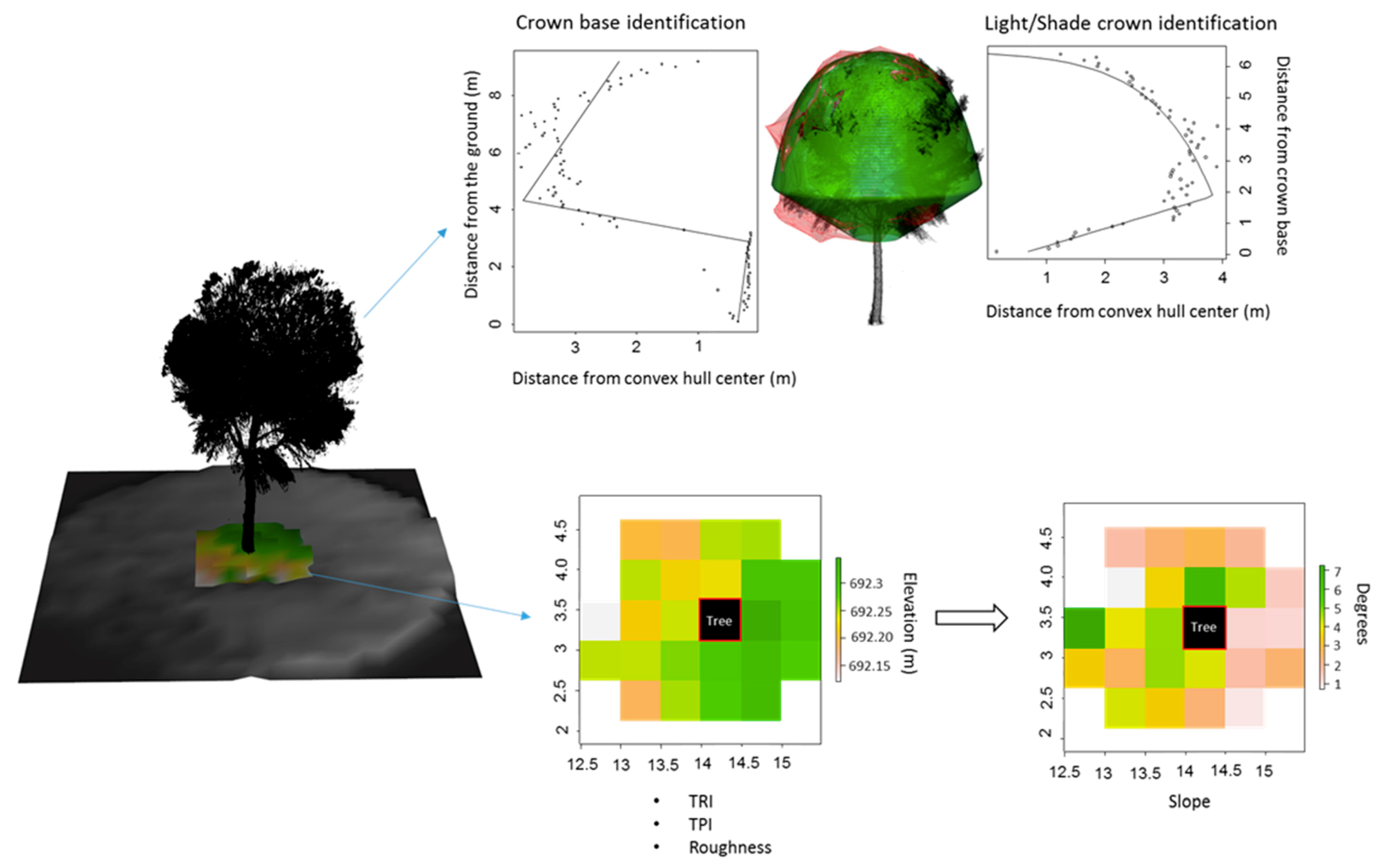
2.3. Terrestrial Laser Scanner Data Acquisition and Metric Extraction
2.3.1. Crown Metrics
- -
- Crown volume (CV) and surface area (SA): volume and area of a 3D alpha-shape fitted to the points belonging to the crown cloud points;
- -
- Light crown volume (LCV) and surface area (LCSA): the farthest distance from the crown center of each 10-cm slice of the crown was used to establish the maximum limits of the crown. A linear–log segmented regression was then used to find the transition between the shade and sun-exposed parts [32]. The volume and surface area of the crown above the transition zone were then calculated by fitting a 3D alpha-shape to the sun-exposed crown;
- -
- Height of the base of the sun-exposed crown (LCB): breakpoint of the linear–log segmented regression;
- -
- Crown length (CL): difference between total tree height and height to crown base;
- -
- Projected crown area (PCA): area of the projected crown on the XY-plane;
- -
- Maximum crown radius (maxCr): maximum of all the radii measured in each 10-cm slice of the crown;
- -
- Crown asymmetry (ASYM): ratio of the smallest and largest crown radius of the projected surface area;
- -
- Crown density: ratio of volume of non-empty 10 cm edged crown voxels to total crown volume, calculated for the complete crown (CDen) and the light crown only (LCDen).
2.3.2. Micro-Topography
- -
- Relative topographic position index (TPI): each surrounding cell of the tree is compared to the cell that contains the tree position by subtracting the height of the cell with the sample tree by mean height of the surrounding cells [33].
- -
- Relative ruggedness index (TRI): TRI is obtained by summing the difference in absolute values of the height of each neighboring cell and the height of the cell with the sample tree [34].
- -
- Roughness: difference between the maximum and minimum height of the cells surrounding the sample tree [35].
- -
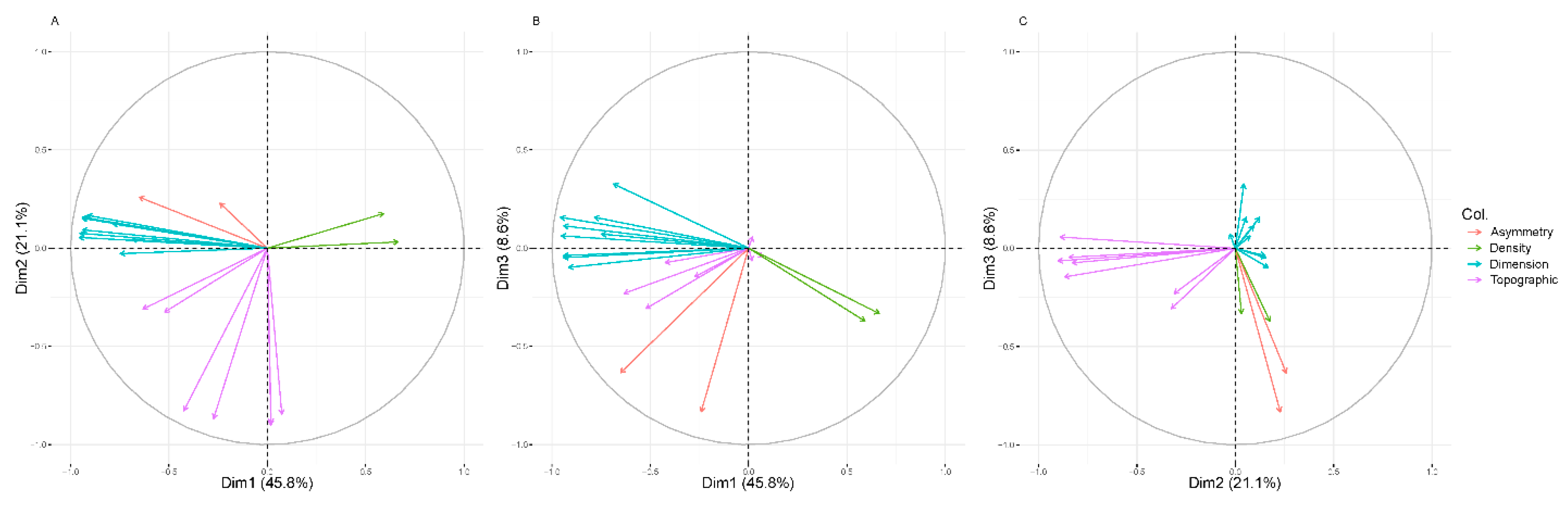
2.4. Data Analysis
2.4.1. Number of Cones
2.4.2. Average Cone Weight
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dassot, M.; Colin, A.; Santenoise, P.; Fournier, M.; Constant, T. Terrestrial laser scanning for measuring the solid wood volume, including branches, of adult standing trees in the forest environment. Comput. Electron. Agric. 2012, 89, 86–93. [Google Scholar] [CrossRef]
- Liang, X.; Kankare, V.; Hyyppä, J.; Wang, Y.; Kukko, A.; Haggrén, H.; Yu, X.; Kaartinen, H.; Jaakkola, A.; Guan, F. Terrestrial laser scanning in forest inventories. ISPRS J. Photogramm. Remote Sens. 2016, 115, 63–77. [Google Scholar] [CrossRef]
- Maas, H.-G.; Bienert, A.; Scheller, S.; Keane, E. Automatic forest inventory parameter determination from terrestrial laser scanner data. Int. J. Remote Sens. 2008, 29, 1579–1593. [Google Scholar] [CrossRef]
- Barbeito, I.; Dassot, M.; Bayer, D.; Collet, C.; Drössler, L.; Löf, M.; Del Rio, M.; Ruiz-Peinado, R.; Forrester, D.I.; Bravo-Oviedo, A. Terrestrial laser scanning reveals differences in crown structure of Fagus sylvatica in mixed vs. pure European forests. For. Ecol. Manag. 2017, 405, 381–390. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Müller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Martin-Ducup, O.; Schneider, R.; Fournier, R.A. Response of sugar maple (Acer saccharum, Marsh.) tree crown structure to competition in pure versus mixed stands. For. Ecol. Manag. 2016, 374, 20–32. [Google Scholar]
- Metz, J.; Seidel, D.; Schall, P.; Scheffer, D.; Schulze, E.-D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra- and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Pimont, F.; Allard, D.; Soma, M.; Dupuy, J.-L. Estimators and confidence intervals for plant area density at voxel scale with T-LiDAR. Remote Sens. Environ. 2018, 215, 343–370. [Google Scholar] [CrossRef]
- Béland, M.; Widlowski, J.L.; Fournier, R.A.; Côté, J.F.; Verstraete, M.M. Estimating leaf area distribution in savanna trees from terrestrial LiDAR measurements. Agric. For. Meteorol. 2011, 151, 1252–1266. [Google Scholar] [CrossRef]
- Martin-Ducup, O.; Schneider, R.; Fournier, R. Analyzing the Vertical Distribution of Crown Material in Mixed Stand Composed of Two Temperate Tree Species. Forests 2018, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Côté, J.-F.; Fournier, R.A.; Frazer, G.W.; Niemann, K.O. A fine-scale architectural model of trees to enhance LiDAR-derived measurements of forest canopy structure. Agric. For. Meteorol. 2012, 166, 72–85. [Google Scholar] [CrossRef]
- Hackenberg, J.; Spiecker, H.; Calders, K.; Disney, M.; Raumonen, P. SimpleTree—an efficient open source tool to build tree models from TLS clouds. Forests 2015, 6, 4245–4294. [Google Scholar] [CrossRef]
- Raumonen, P.; Kaasalainen, M.; Åkerblom, M.; Kaasalainen, S.; Kaartinen, H.; Vastaranta, M.; Holopainen, M.; Disney, M.; Lewis, P. Fast Automatic Precision Tree Models from Terrestrial Laser Scanner Data. Remote Sens. 2013, 5, 491–520. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez de Tanago, J.; Lau, A.; Bartholomeus, H.; Herold, M.; Avitabile, V.; Raumonen, P.; Martius, C.; Goodman, R.C.; Disney, M.; Manuri, S. Estimation of above-ground biomass of large tropical trees with terrestrial LiDAR. Methods Ecol. Evol. 2018, 9, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Momo Takoudjou, S.; Ploton, P.; Sonké, B.; Hackenberg, J.; Griffon, S.; De Coligny, F.; Kamdem, N.G.; Libalah, M.; Mofack, G.I.; Le Moguédec, G. Using terrestrial laser scanning data to estimate large tropical trees biomass and calibrate allometric models: A comparison with traditional destructive approach. Methods Ecol. Evol. 2018, 9, 905–916. [Google Scholar] [CrossRef]
- Guo, Q.; Wu, F.; Pang, S.; Zhao, X.; Chen, L.; Liu, J.; Xue, B.; Xu, G.; Li, L.; Jing, H. Crop 3D—a LiDAR based platform for 3D high-throughput crop phenotyping. Sci. China Life Sci. 2018, 61, 328–339. [Google Scholar] [CrossRef]
- Ovando, P.; Campos, P.; Calama, R.; Montero, G. Landowner net benefit from stone pine (Pinus pinea L.) afforestation of dry-land cereal fields in Valladolid, Spain. J. For. Econ. 2010, 16, 83–100. [Google Scholar] [CrossRef]
- Mutke, S.; Sievänen, R.; Nikinmaa, E.; Perttunen, J.; Gil, L. Crown architecture of grafted Stone pine (Pinus pinea L.): shoot growth and bud differentiation. Trees 2005, 19, 15–25. [Google Scholar] [CrossRef]
- Calama, R.; Alonso, F.J.G.; Casanueva, G.M.; Regneri, S.M.; Conde, M.; González, G.M.; Minguez, M.P. Enhanced tools for predicting annual stone pine (Pinus pinea L.) cone production at tree and forest scale in inner Spain. For. Syst. 2016, 25, 14. [Google Scholar] [CrossRef] [Green Version]
- Mutke, S.; Calama, R.; Nasrallah Neaymeh, E.; Roques, A. Impact of the Dry Cone Syndrome on commercial kernel yield of stone pine cones. In Options Méditerranéennes Sér. Sémin. Méditerranéens; CIHEAM: Zaragoza, Spain, 2017; pp. 79–84. [Google Scholar]
- Calama, R.; Gordo, F.J.; Mutke, S.; Montero, G. An empirical ecological-type model for predicting stone pine (Pinus pinea L.) cone production in the Northern Plateau (Spain). For. Ecol. Manag. 2008, 255, 660–673. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Cañellas, I.; Calama, R.; Gordo, J.; Sánchez-González, M. Thinning increases cone production of stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ann. For. Sci. 2013, 70, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Loewe, V.; Venegas, A.; Delard, C.; González, M. Thinning effect in two young stone pine plantations (Pinus pinea L.) in central southern Chile. Opt. Méditerranéennes 2013, 105, 44–55. [Google Scholar]
- Boutheina, A.; El-Aouni, M.H.; Balandier, P. Influence of stand and tree attributes and silviculture on cone and seed productions in forests of Pinus pinea L. in northern Tunisia. Opt. Méditerranéennes Sér. Sémin. Méditerranéens 2013, 105, 9–14. [Google Scholar]
- Ruiz de la Torre, J. Árboles y arbustos de la España peninsular. ETS Ing. Montes Madr. Spain 1979. [Google Scholar]
- Mutke, S.; Calama, R.; González-Martínez, S.C.; Montero, G.; Javier Gordo, F.; Bono, D.; Gil, L. Mediterranean Stone Pine: Botany and Horticulture. Hortic. Rev. 2012, 39, 153–201. [Google Scholar]
- Rodrigues, A.; Silva, G.L.; Casquilho, M.; Freire, J.; Carrasquinho, I.; Tomé, M. Linear mixed modelling of cone production for Stone Pine in Portugal. Silva Lusit. 2014, 22, 1–27. [Google Scholar]
- United Nations Environment Programme (UNEP) World atlas of desertification. In World Atlas of Desertification; UNEP: Nairobi, Kenya, 1992.
- Muggeo, V.M.R. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Muggeo, V.M. Segmented: an R package to fit regression models with broken-line relationships. R News 2008, 8, 20–25. [Google Scholar]
- Barber, C.B.; Habel, K.; Grasman, R.; Stahel, A.; Stahel, A.; Sterratt, D.C. Geometry: Mesh Generation and Surface Tesselation. 2014. Available online: https://davidcsterratt.github.io/geometry/ (accessed on 2 January 2020).
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer: Berlin, Germany, 2009. [Google Scholar]
- Riley, S.J. Index that quantifies topographic heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Wilson, M.F.; O’Connell, B.; Brown, C.; Guinan, J.C.; Grehan, A.J. Multiscale terrain analysis of multibeam bathymetry data for habitat mapping on the continental slope. Mar. Geod. 2007, 30, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Dartnell, P. Applying Remote Sensing Techniques to Map Seafloor Geology/Habitat Relationships. Ph.D. Thesis, San Francisco State University, San Francisco, CA, USA, 2000. [Google Scholar]
- Burrough, P.A.; McDonnell, R.; McDonnell, R.A.; Lloyd, C.D. Principles of Geographical Information Systems; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- VanDerWal, J.; Falconi, L.; Januchowski, S.; Shoo, L.; Storlie, C. SDMTools: Species Distribution Modelling Tools: Tools for processing data associated with species distribution modelling exercises. R Package Version 2014, 1, 1–221. [Google Scholar]
- King, J.R.; Jackson, D.A. Variable selection in large environmental data sets using principal components analysis. Environmetrics 1999, 10, 67–77. [Google Scholar] [CrossRef]
- Quinn, G.G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Hall, D.B. Zero-Inflated Poisson and Binomial Regression with Random Effects: A Case Study. Biometrics 2000, 56, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Rizopoulos, D. GLMMadaptive: Generalized Linear Mixed Models Using Adaptive Gaussian Quadrature. R Package Version 0.4-0. 2018. Available online: https://rdrr.io/cran/GLMMadaptive/man/GLMMadaptive.html (accessed on 27 December 2019).
- Bates, D.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c); Universidade de Lisboa: Lisbon, Portugal, 2017. [Google Scholar]
- Wagenmakers, E.-J.; Farrell, S. AIC model selection using Akaike weights. Psychon. Bull. Rev. 2004, 11, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Cañadas, M.N. Pinus pinea L. en el Sistema Central (Valles del Tiétar y del Alberche): desarrollo de un modelo de crecimiento y producción de piña. Ph.D. Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 2000. [Google Scholar]
- Freire, J.P.A. Modelação do crescimento e da produção de pinha no pinheiro manso. Ph.D. Thesis, ISA-UTL Lisbon Port, Lisboa, Portugal, 2009. [Google Scholar]
- Piqué, M.; Vericat, P.; Beltran, M.; Calama, R.; Cervera, T. Models de Gestió per a les Pinedes de pi Pinyer (Pinus pinea L.): Producció de Fusta i Pinya i Prevenció de Incendis Forestales; Centre de la Propietat Forestal. Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural; Generalitat de Catalunya: Barcelona, Spain, 2016; ISBN B17190-2015. [Google Scholar]
- Castellani, C. La produzione legnosa e del fruto e la durata economico delle pinete coetanee di pino domestico (Pinus pinea L.) in un complesso assestato a prevalente funzione produttiva in Italia. Ann. ISAFA 1989, 12, 161–221. [Google Scholar]
- Calama, R.; Mutke, S.; Tomé, J.; Gordo, J.; Montero, G.; Tomé, M. Modelling spatial and temporal variability in a zero-inflated variable: The case of stone pine (Pinus pinea L.) cone production. Ecol. Model. 2011, 222, 606–618. [Google Scholar] [CrossRef]
- Montero, G.; Calama, R.; Ruiz Peinado, R. Selvicultura de Pinus pinea L. In Compendio de Selvicultura de Especies. INIA: Fundación Conde del Valle de Salazar, Madrid; INIA Andes: Montevideo, Uruguay, 2008; pp. 431–470. [Google Scholar]
- Sirois, L. Spatiotemporal variation in black spruce cone and seed crops along a boreal forest-tree line transect. Can. J. For. Res. 2000, 30, 900–909. [Google Scholar] [CrossRef]
- Verkaik, I.; Espelta, J.M. Post-fire regeneration thinning, cone production, serotiny and regeneration age in Pinus halepensis. For. Ecol. Manag. 2006, 231, 155–163. [Google Scholar] [CrossRef]
- Bravo Oviedo, F.; Maguire, D.A.; González Martínez, S.C. Factors affecting cone production in Pinus pinaster Ait.: lack of growth-reproduction trade-offs but significant effects of climate and tree and stand characteristics. For. Syst. 2017, 26, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lanner, R.M. An observation on apical dominance and the umbrella-crown of Italian stone pine (Pinus pinea, Pinaceae). Econ. Bot. 1989, 43, 128–130. [Google Scholar]
- Wang, W.-J.; Watanabe, Y.; Endo, I.; Kitaoka, S.; Koike, T. Seasonal changes in the photosynthetic capacity of cones on a larch (Larix kaempferi) canopy. Photosynthetica 2006, 44, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Mutke, S.; Calama, R.; Guadano, C.; Leon, D.; Gordo, J.; Montero, G. Efecto de la poda sobre la producción de piña en pino piñonero injertado. In Proceedings of the Poster. 7o Congreso Forestal, Plasencia, Spain, 26–30 July 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot-Level Variables | |
|---|---|
| Stand density (SD, st∙ha−1) | 167 (101) |
| Stand age (A, years) | 81.4 (27.3) |
| Basal area (BA, m2∙ha−1) | 20.44 (10.02) |
| Mean quadratic diameter (MQD, cm) | 40.3 (9.12) |
| Dominant stand height (domH, m) | 12.56 (3.15) |
| Site index (SI, m) | 15.33 (3.06) |
| Tree-level variables | |
| Number of cones | 14.1 (20.4) |
| Average cone weight (wt, g) | 196.9 (74.2) |
| Diameter at breast height (dbh, cm) | 40.9 (9.0) |
| Crown length (CL, m) | 6.85 (1.44) |
| Crown diameter (CD, m) | 7.30 (1.71) |
| Total tree height (H, m) | 12.42 (3.16) |
| LiDAR-derived variables | |
| Topographic position index (TPI) | 0.0641 (0.0426) |
| Topographic roughness index (TRI) | 0.0766 (0.0302) |
| Roughness (R) | 0.1974 (0.0705) |
| Slope (S) | 0.0778 (0.0451) |
| Crown length (CLL, m) | 7.43 (1.91) |
| Projected crown area (PCA, m2) | 54.65 (22.57) |
| Projected crown radius (CR, m) | 4.09 (0.84) |
| Maximum crown radius (maxCr, m) | 3.85 (0.82) |
| Crown surface area (SA, m2) | 129.33 (64.36) |
| Crown volume (CV, m3) | 189.27 (98.99) |
| Crown density (CDen) | 0.4010 (0.1139) |
| Height to light crown base (LCB, m) | 8.74 (3.47) |
| Light crown volume (LCV, m3) | 104.66 (79.36) |
| Light crown surface area (LCSA, m2) | 34.41 (33.68) |
| Light crown density (LCDen) | 0.3656 (0.1107) |
| Crown asymmetry (ASYM) | 0.5290 (0.1882) |
| Minimum projected crown radius (minCR, m) | 2.89 (1.09) |
| Topography | Crown Dimensions | Crown Density | Crown Asymmetry | |
|---|---|---|---|---|
| Topography | 0.57 | |||
| Crown dimensions | 1.04 | 0.22 | ||
| Crown density | 1.24 | 1.57 | 0.17 | |
| Crown asymmetry | 1.22 | 0.92 | 1.17 | 0.45 |
| Model | Number of Variables in Logistic Component | Number of Variables in Negative Binomial Component | Log (Dispersion) | AIC | AIC Weight | ||
|---|---|---|---|---|---|---|---|
| 0. Null model | 0 | 0 | 1.0280 | 2.9193 | 0.2140 | 1400.19 | >0.0001 |
| 1. Plot | 1 | 3 | 0.4256 | 4.6833 | 0.2266 | 1383.76 | >0.0001 |
| 2. Tree | 1 | 2 | 0.5222 | 3.3970 | 0.4205 | 1349.8 | 0.0001 |
| 3. LST | 2 | 2 | 0.4850 | 5.5951 | 0.4788 | 1337.01 | 0.0816 |
| 4. Plot and tree | 1 | 5 | 0.3926 | 3.2902 | 0.4324 | 1346.53 | 0.0007 |
| 5. Plot and LST | 2 | 4 | 0.4252 | 4.6492 | 0.5127 | 1332.17 | 0.9176 |
| Model | Number of Variables | AICc | AIC Weight | |||
|---|---|---|---|---|---|---|
| 0. Null | 0 | 1788 | 8190 | 2132 | 1814.91 | 0.0002 |
| 1. Plot | 1 | 1418 | 8195 | 2131 | 1807.89 | 0.0072 |
| 2. Tree | 1 | 1754 | 8377 | 2092 | 1809.12 | 0.0039 |
| 3. LiDAR | 2 | 1418 | 8295 | 2105 | 1803.78 | 0.0562 |
| 4. Plot and tree | 1 | 1754 | 8377 | 2092 | 1809.12 | 0.0039 |
| 5. Plot and liDAR | 4 | 1257 | 8306 | 2123 | 1798.17 | 0.9286 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, R.; Calama, R.; Martin-Ducup, O. Understanding Tree-to-Tree Variations in Stone Pine (Pinus pinea L.) Cone Production Using Terrestrial Laser Scanner. Remote Sens. 2020, 12, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12010173
Schneider R, Calama R, Martin-Ducup O. Understanding Tree-to-Tree Variations in Stone Pine (Pinus pinea L.) Cone Production Using Terrestrial Laser Scanner. Remote Sensing. 2020; 12(1):173. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12010173
Chicago/Turabian StyleSchneider, Robert, Rafael Calama, and Olivier Martin-Ducup. 2020. "Understanding Tree-to-Tree Variations in Stone Pine (Pinus pinea L.) Cone Production Using Terrestrial Laser Scanner" Remote Sensing 12, no. 1: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12010173