Non-Destructive Early Detection and Quantitative Severity Stage Classification of Tomato Chlorosis Virus (ToCV) Infection in Young Tomato Plants Using Vis–NIR Spectroscopy



 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. ToCV Infection and Quantitative Analysis
2.3. Optical Measurements
2.4. Spectral Data Pre-Processing and Feature Selection
2.5. Feature Selection
2.6. Class Division
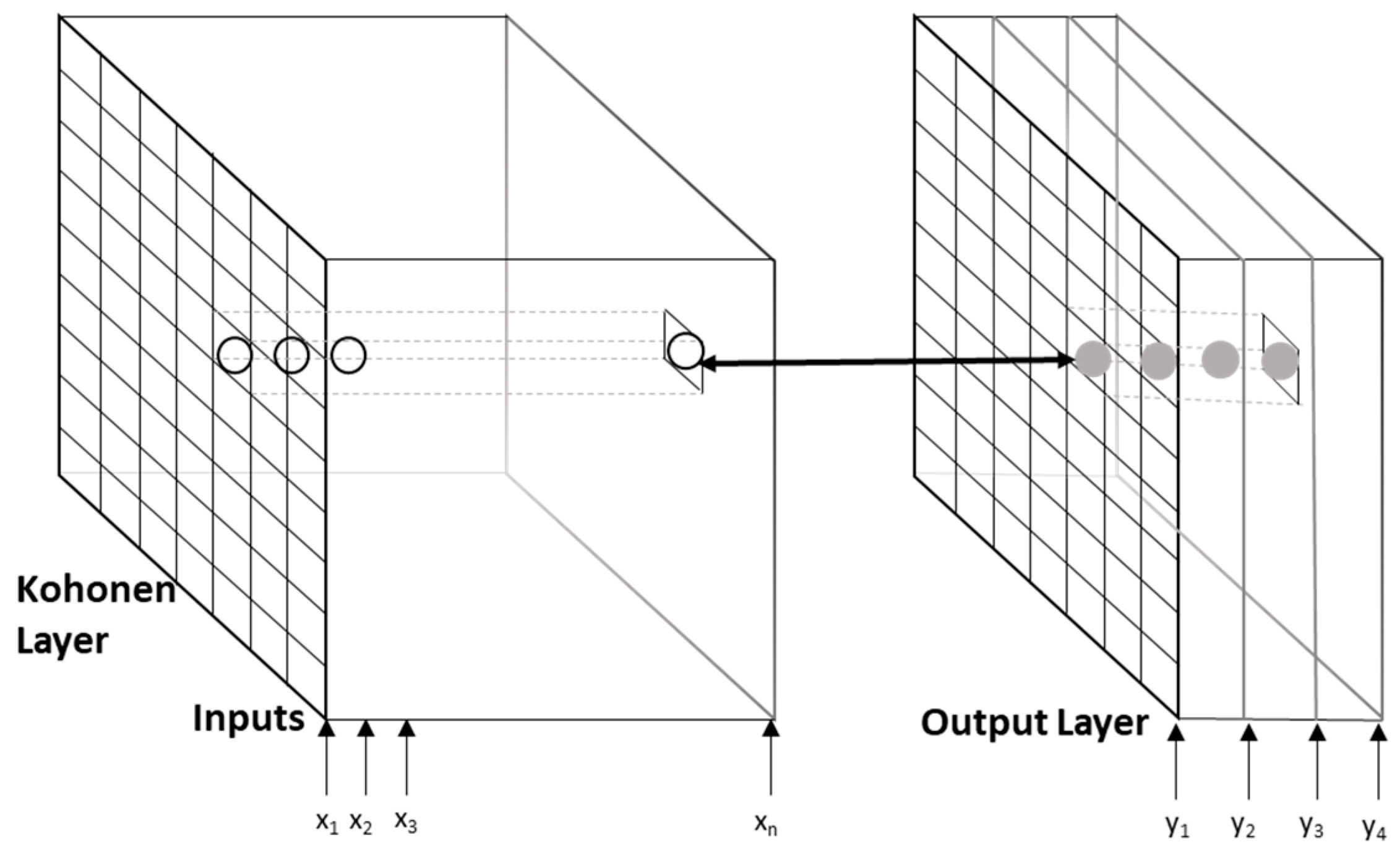
2.7. Machine Learning Techniques
2.8. Model Evaluation Metrics
3. Results
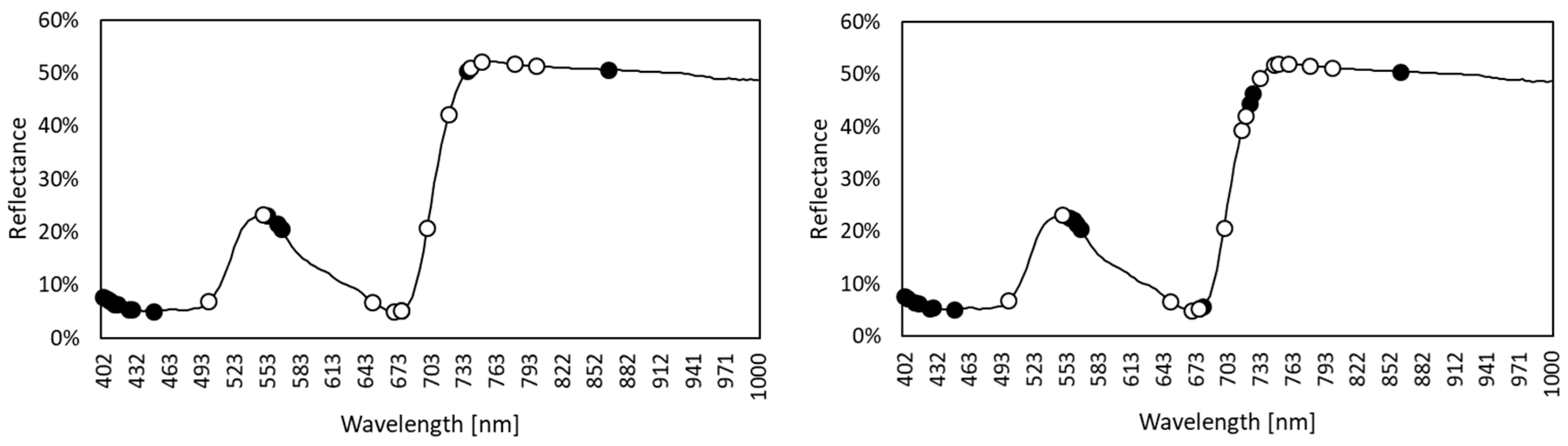
3.1. Spectral Data Overview
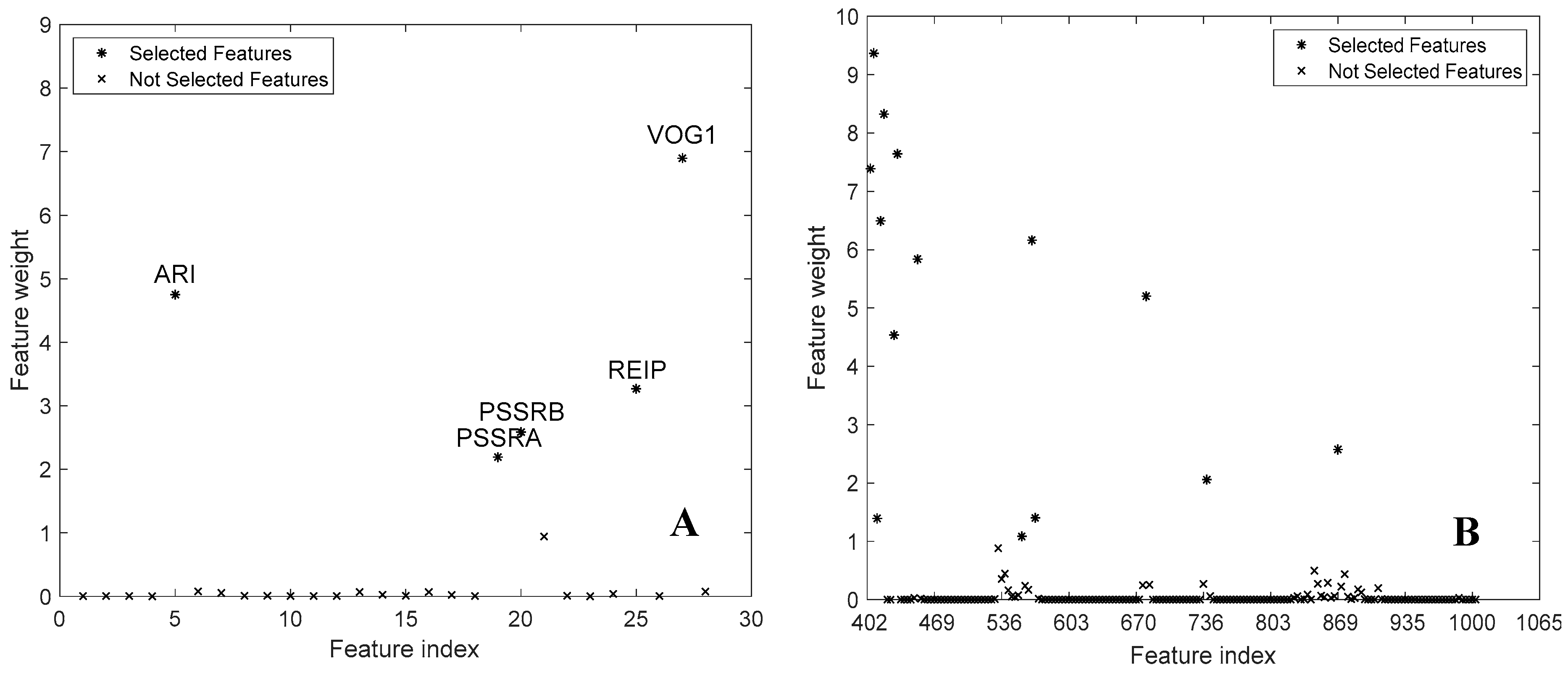
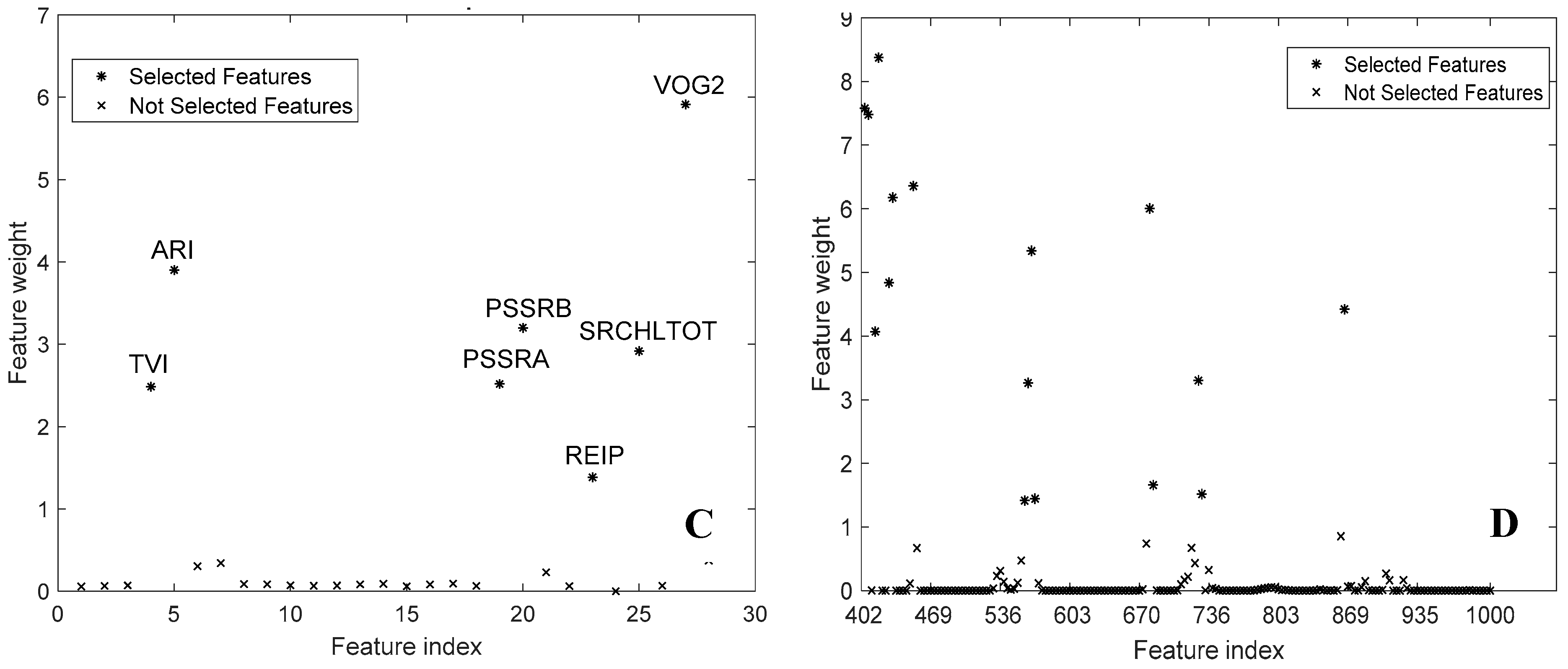
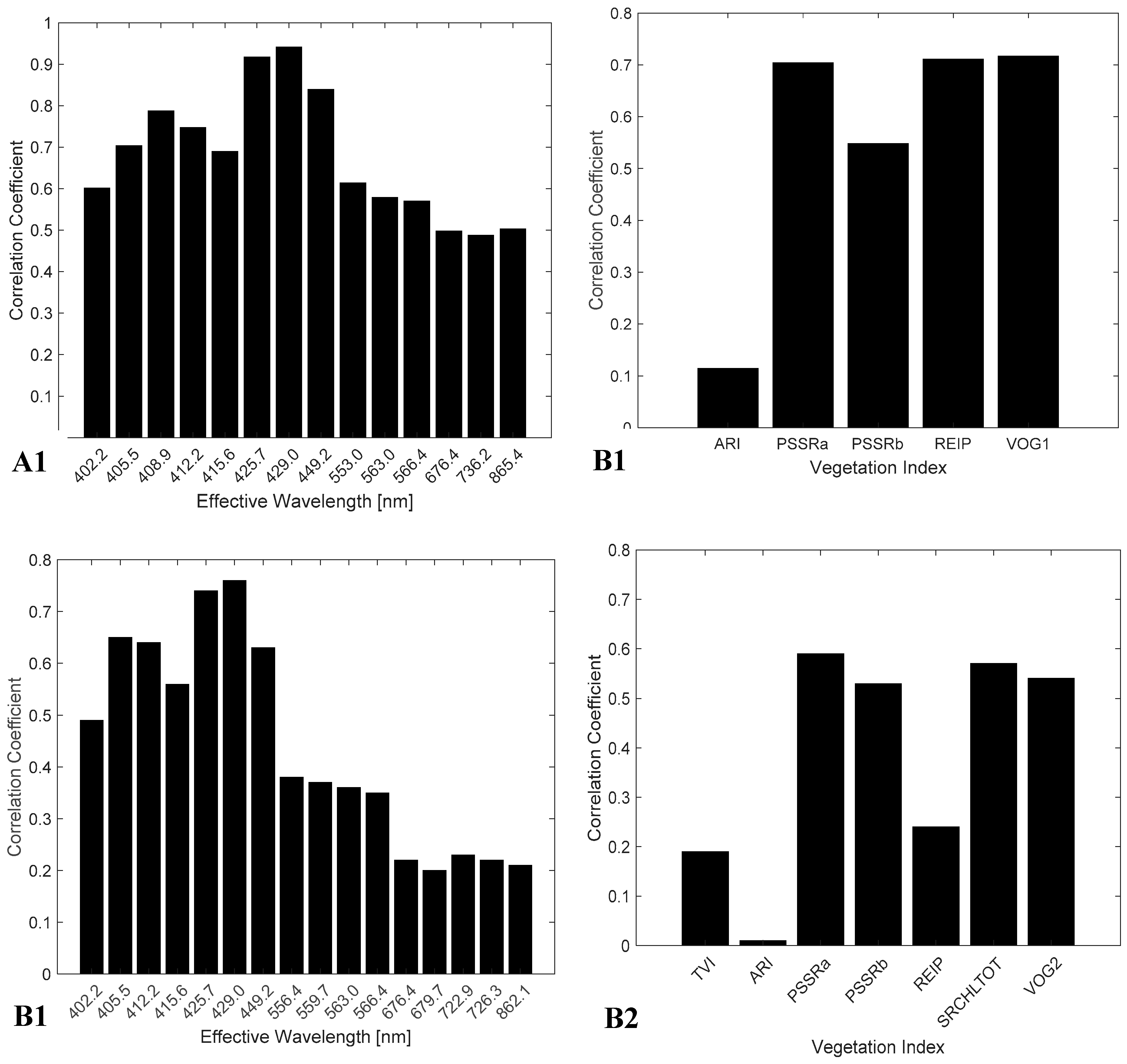
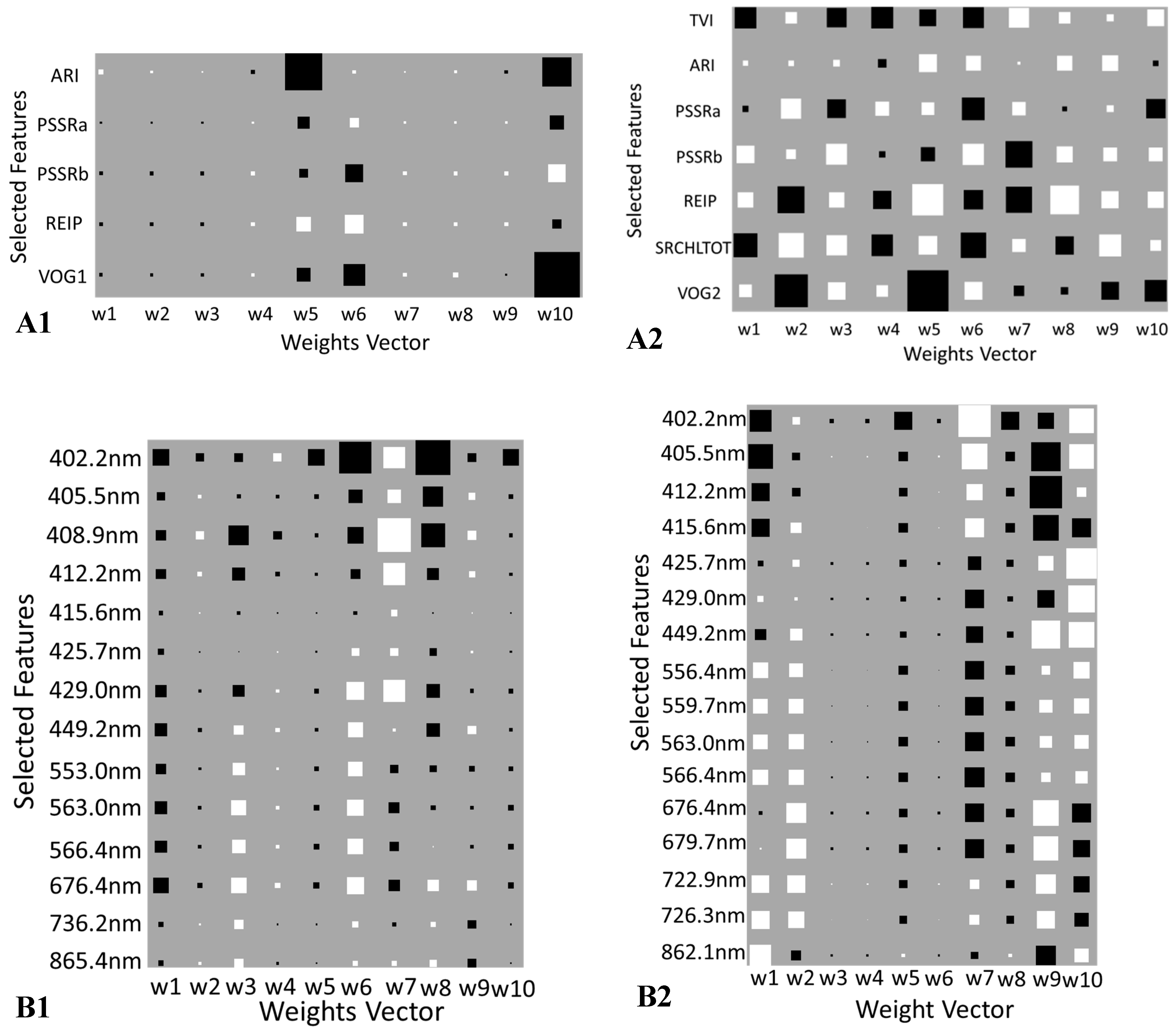
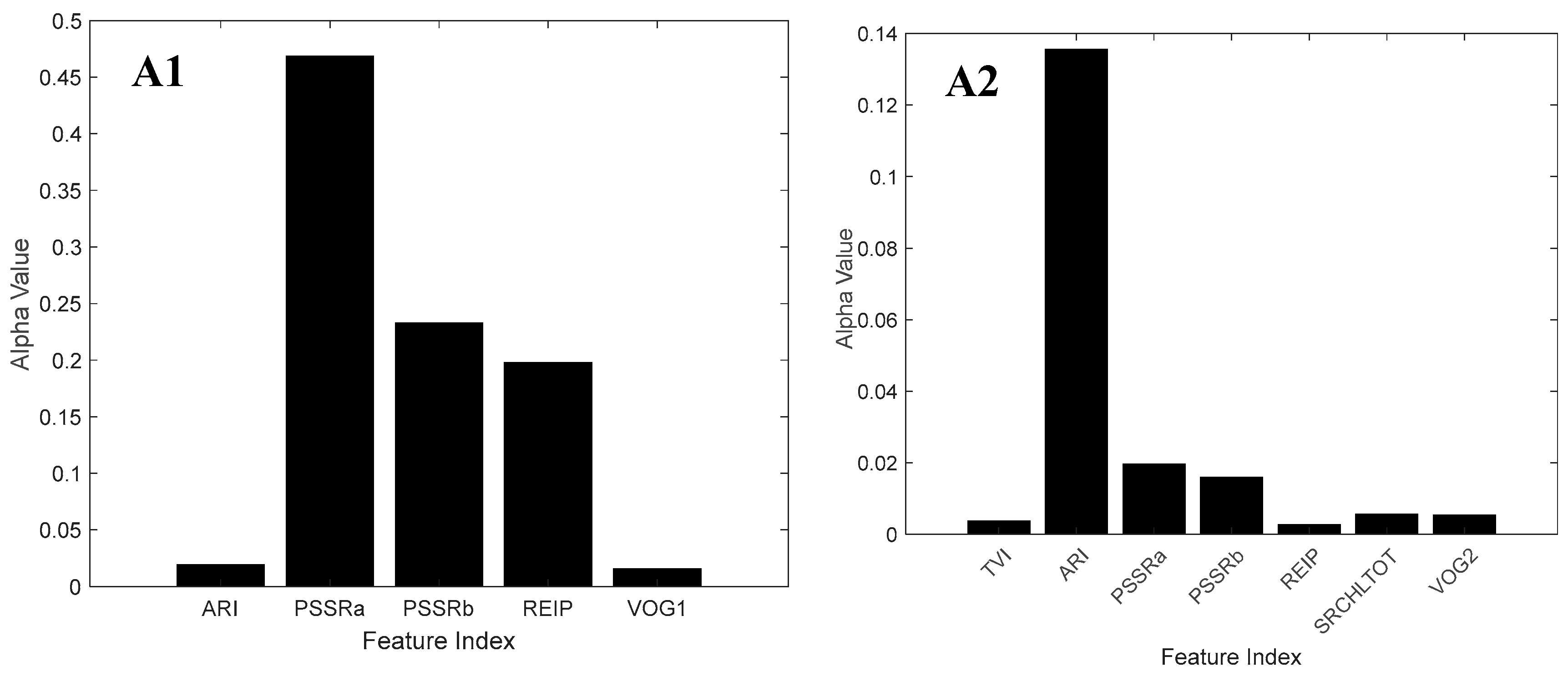
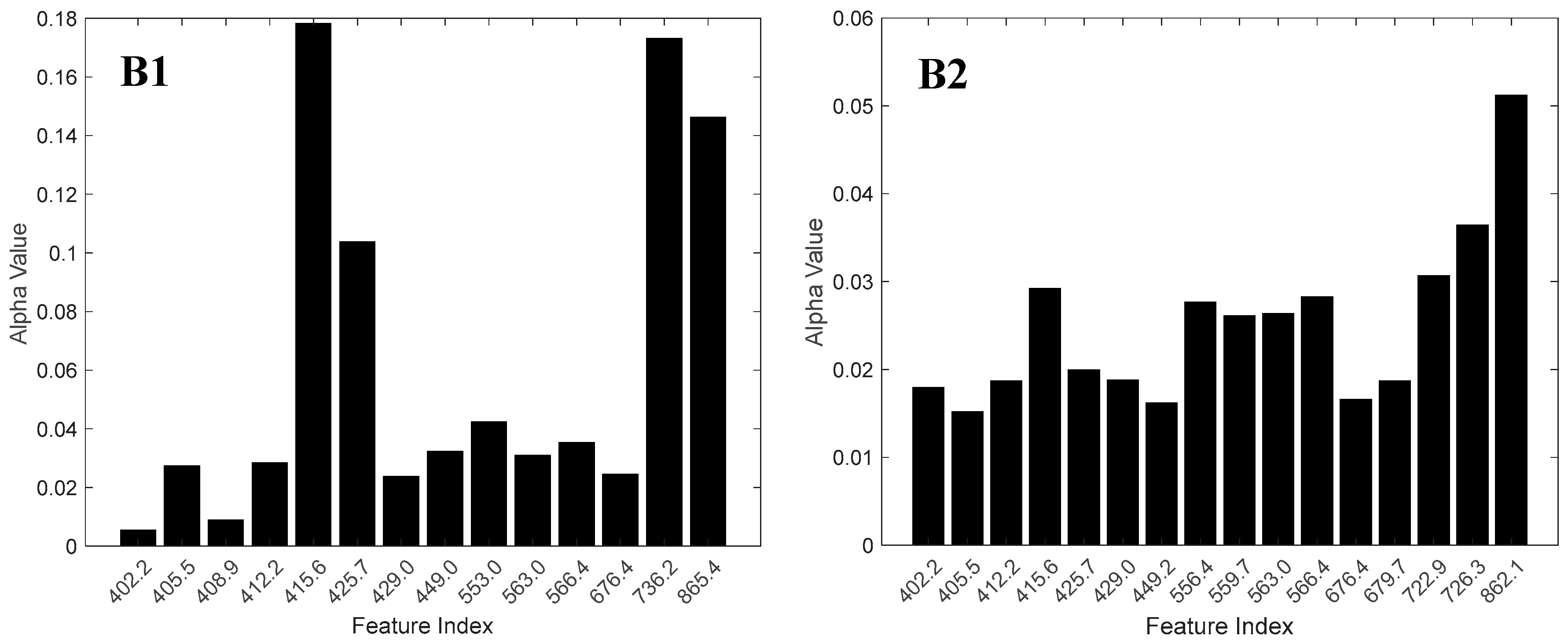
3.2. Feature Selection
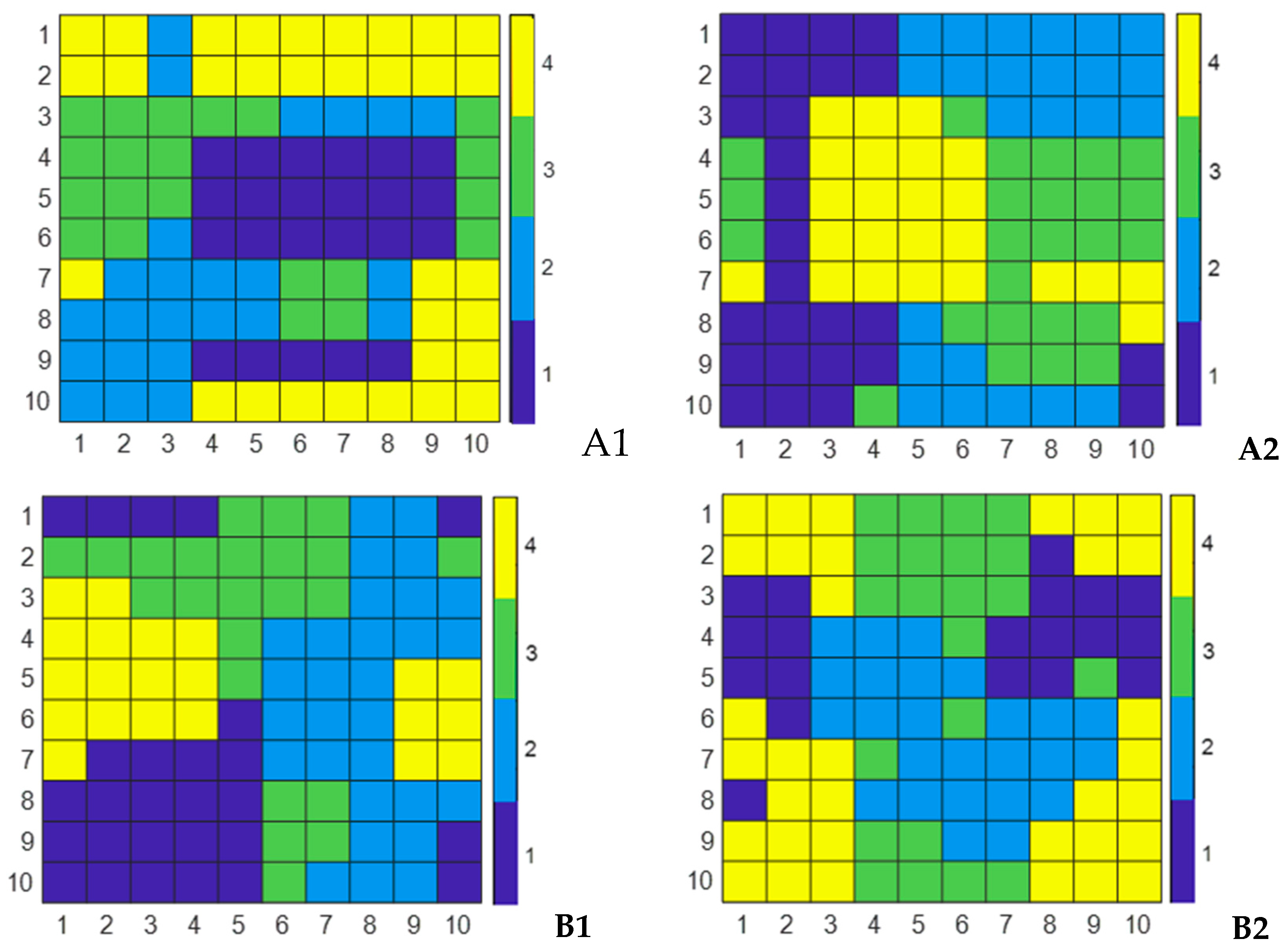
3.3. XY-F classifier
3.4. MLP–ARD Classifier
4. Discussion
4.1. General and Spectral Data Overview
4.2. The Effect of Outliers in the Models
4.3. Spectral Bands and Vegetation Indices Selected by NCA
4.4. Classifier Results
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liakos, K.G.; Busato, P.; Moshou, D.; Pearson, S.; Bochtis, D. Machine learning in agriculture: A review. Sensors 2018, 18, 2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulla, D.J. Twenty five years of remote sensing in precision agriculture: Key advances and remaining knowledge gaps. Biosyst. Eng. 2013, 114, 358–371. [Google Scholar] [CrossRef]
- Golhani, K.; Balasundram, S.K.; Vadamalai, G.; Pradhan, B. A review of neural networks in plant disease detection using hyperspectral data. Inf. Process. Agric. 2018, 5, 354–371. [Google Scholar] [CrossRef]
- Berdugo, C.A.; Mahlein, A.K.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Sensors and imaging techniques for the assessment of the delay of wheat senescence induced by fungicides. Funct. Plant Biol. 2013, 40, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef]
- Mahlein, A.K.; Oerke, E.C.; Steiner, U.; Dehne, H.W. Recent advances in sensing plant diseases for precision crop protection. Eur. J. Plant Pathol. 2012, 133, 197–209. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Wisler, G.C. Vector specificity, host range, and genetic diversity of Tomato chlorosis virus. Plant Dis. 2006, 90, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Orfanidou, C.G.; Dimitriou, C.; Papayiannis, L.C.; Maliogka, V.I.; Katis, N.I. Epidemiology and genetic diversity of criniviruses associated with tomato yellows disease in Greece. Virus Res. 2014, 186, 120–129. [Google Scholar] [CrossRef]
- Fiallo-Olive, E.; Navas-Castillo, J. Tomato chlorosis virus, an emergent plant virus still expanding its geographical and host ranges. Mol. Plant Pathol. 2019, 20, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Wisler, G.C.; Li, R.-H.; Liu, H.Y.; Lowry, D.S.; Duffus, J.E. Tomato Chlorosis Virus: A New Whitefly-Transmitted, Phloem-Limited, Bipartite Closterovirus of Tomato. Phytopathology 1998, 88, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Gold, K.M.; Townsend, P.A.; Chlus, A.; Herrmann, I.; Couture, J.J.; Larson, E.R.; Gevens, A.J. Hyperspectral Measurements Enable Pre-Symptomatic Detection and Differentiation of Contrasting Physiological Effects of Late Blight and Early Blight in Potato. Remote Sens. 2020, 12, 286. [Google Scholar] [CrossRef] [Green Version]
- Fernández, C.I.; Leblon, B.; Haddadi, A.; Wang, K.; Wang, J. Potato late blight detection at the leaf and canopy levels based in the red and red-edge spectral regions. Remote Sens. 2020, 12, 1292. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, I.; Berenstein, M.; Paz-Kagan, T.; Sade, A.; Karnieli, A. Spectral assessment of two-spotted spider mite damage levels in the leaves of greenhouse-grown pepper and bean. Biosyst. Eng. 2017, 157, 72–85. [Google Scholar] [CrossRef]
- Papayiannis, L.C.; Harkou, I.S.; Markou, Y.M.; Demetriou, C.N.; Katis, N.I. Rapid discrimination of Tomato chlorosis virus, Tomato infectious chlorosis virus and co-amplification of plant internal control using real-time RT-PCR. J. Virol. Methods 2011, 176, 53–59. [Google Scholar] [CrossRef]
- Duffus, J.E.; Liu, H.Y.; Wisler, G.C. Tomato infectious chlorosis virus—A new clostero-like virus transmitted by Trialeurodes vaporariorum. Eur. J. Plant. Pathol. 1996, 102, 219–226. [Google Scholar] [CrossRef]
- Jacquemond, M.; Verdin, E.; Dalmon, A.; Guilbaud, L.; Gognalons, P. Serological and molecular detection of Tomato chlorosis virus and Tomato infectious chlorosis virus in tomato. Plant. Pathol. 2009, 58, 210–220. [Google Scholar] [CrossRef]
- Morris, J.; Steel, E.; Smith, P.; Boonham, N.; Spence, N.; Barker, I. Host range studies for Tomato chlorosis virus and Cucumber vein yellowing virus transmitted by Bemisia tabaci (Gennadius). Eur. J. Plant. Pathol. 2005, 114, 265–273. [Google Scholar] [CrossRef]
- Thomas, S.; Kuska, M.T.; Bohnenkamp, D.; Brugger, A.; Alisaac, E.; Wahabzada, M.; Behman, J.; Mahlein, A.K. Benefits of hyperspectral imaging for plant disease detection and plant protection: A technical perspective. J. Plant Dis. Prot. 2018, 125, 5–20. [Google Scholar] [CrossRef]
- Guan, J.; Nutter, F.W., Jr. Relationships between defoliation, leaf area index, canopy reflectance, and forage yield in the alfalfa-leaf spot pathosystem. Comput. Electr. Agric. 2002, 37, 97–112. [Google Scholar] [CrossRef]
- West, J.S.; Bravo, C.; Oberti, R.; Lemaire, D.; Moshou, D.; McCartney, H.A. The potential of optical canopy measurements for targeted control of field crop diseases. Ann. Rev. Phytopathol. 2003, 41, 593–614. [Google Scholar] [CrossRef] [Green Version]
- Bravo, C.; Moshou, D.; West, J.; McCartney, A.; Ramon, H. Early disease detection in wheat fields using spectral reflectance. Biosyst. Eng. 2003, 84, 137–145. [Google Scholar] [CrossRef]
- Moshou, D.; Bravo, C.; Oberti, R.; West, J.; Bodria, L.; McCartney, A.; Ramon, H. Plant disease detection based on data fusion of hyper-spectral and multi-spectral fluorescence imaging using Kohonen maps. Real Time Imaging 2005, 11, 75–83. [Google Scholar] [CrossRef]
- Grisham, M.P.; Johnson, R.M.; Zimba, P.V. Detecting Sugarcane yellow leaf virus infection in asymptomatic leaves with hyperspectral remote sensing and associated leaf pigment changes. J. Virol. Methods 2010, 167, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Bürling, K.; Hunsche, M.; Noga, G. Presymptomatic Detection of Powdery Mildew Infection in Winter Wheat Cultivars by Laser-Induced Fluorescence. Appl. Spectrosc. 2012, 66, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Arens, N.; Backhaus, A.; Döll, S.; Fischer, S.; Seiffert, U.; Mock, H.-P. Non-invasive Presymptomatic Detection of Cercospora beticola Infection and Identification of Early Metabolic Responses in Sugar Beet. Front. Plant Sci. 2016, 7, 1377. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Chu, B.; Zhang, C.; Liu, F.; Jiang, L.; He, Y. Hyperspectral imaging for presymptomatic detection of tobacco disease with successive projections algorithm and machine-learning classifiers. Sci. Rep. 2017, 7, 4125. [Google Scholar] [CrossRef] [Green Version]
- Moshou, D.; Bravo, C.; West, J.; Wahlen, S.; McCartney, A.; Ramon, H. Automatic detection of ‘yellow rust’ in wheat using reflectance measurements and neural networks. Comput. Electron. Agric. 2004, 44, 173–188. [Google Scholar] [CrossRef]
- Moshou, D.; Bravo, C.; Oberti, R.; West, J.S.; Ramon, H.; Vougioukas, S.; Bochtis, D. Intelligent multi-sensor system for the detection and treatment of fungal diseases in arable crops. Biosyst. Eng. 2011, 108, 311–321. [Google Scholar] [CrossRef]
- Pantazi, X.E.; Tamouridou, A.A.; Alexandridis, T.K.; Lagopodi, A.L.; Kashefi, J.; Moshou, D. Evaluation of hierarchical self-organising maps for weed mapping using UAS multispectral imagery. Comput. Electron. Agric. 2017, 139, 224–230. [Google Scholar] [CrossRef]
- Tamouridou, A.; Pantazi, X.; Alexandridis, T.; Lagopodi, A.; Kontouris, G.; Moshou, D. Spectral identification of disease in weeds using multilayer perceptron with automatic relevance determination. Sensors 2018, 18, 2770. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Asaari, M.S.M.; Herrero-Langreo, A.; Lohumi, S.; Diezma, B.; Scheunders, P. Close range hyperspectral imaging of plants: A review. Biosyst. Eng. 2017, 164, 49–67. [Google Scholar] [CrossRef]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Haas, J.; Lozano, E.R.; Poppy, G.M. A simple, light clip-cage for experiments with aphids. Agric. For. Entomol. 2018, 20, 589–592. [Google Scholar] [CrossRef]
- Orfanidou, C.G.; Pappi, P.G.; Efthimiou, K.E.; Katis, N.I.; Maliogka, V.I. Transmission of Tomato chlorosis virus (ToCV) by Bemisiatabaci biotype Q and evaluation of four weed species as viral sourced. Plant Dis. 2016, 100, 2043–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morellos, A.; Pantazi, X.E.; Moshou, D.; Alexandridis, T.; Whetton, R.; Tziotzios, G.; Wiebensohn, J.; Bill, R.; Mouazen, A.M. Machine learning based prediction of soil total nitrogen, organic carbon and moisture content by using VIS-NIR spectroscopy. Biosyst. Eng. 2016, 152, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Kochubey, S.M.; Kazantsev, T.A. Derivative vegetation indices as a new approach in remote sensing of vegetation. Front. Earth Sci. 2012, 6, 188–195. [Google Scholar] [CrossRef]
- Yao, X.; Ren, H.; Cao, Z.; Tian, Y.; Cao, W.; Zhu, Y.; Cheng, T. Detecting leaf nitrogen content in wheat with canopy hyperspectrum under different soil backgrounds. Int. J. Appl. Earth Obs. Geoinf. 2014, 32, 114–124. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Xu, H.; Caramanis, C.; Mannor, S. Outlier-robust PCA: The high-dimensional case. IEEE Trans. Inf. Theory 2013, 59, 546–572. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Rumpf, T.; Welke, P.; Dehne, H.W.; Plümer, L.; Steiner, U.; Oerke, E.C. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Rouse, J.W.; Hass, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. In Proceedings of the Third Earth Resources Technology Satellite-1 Symposium, Washington, DC, USA, 10–14 December 1972. [Google Scholar]
- Liu, L.; Huang, W.; Pu, R.; Wang, J. Detection of Internal Leaf Structure Deterioration Using a New Spectral Ratio Index in the Near-Infrared Shoulder Region. J. Integr. Agric. 2014, 13, 760–769. [Google Scholar] [CrossRef]
- Barnes, E.M.; Clarke, T.R.; Richards, S.E.; Colaizzi, P.D.; Haberland, J.; Kostrzewski, M.; Lascano, R.J. Coincident detection of crop water stress, nitrogen status and canopy density using ground based multispectral data. In Proceedings of the Fifth International Conference on Precision Agriculture, Bloomington, MN, USA, 16–19 July 2000; Volume 1619. [Google Scholar]
- Broge, N.H.; Leblanc, E. Comparing prediction power and stability of broadband and hyperspectral vegetation indices for estimation of green leaf area index and canopy chlorophyll density. Remote Sens. Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.A. Ratios of leaf reflectances in narrow wavebands as indicators of plant stress. Remote Sens. 1994, 15, 697–703. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of chlorophyll a, chlorophyll b, chlorophyll a + b, and total carotenoid content in eucalyptus leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]
- Datt, B. A new reflectance index for remote sensing of chlorophyll content in higher plants: Tests using Eucalyptus leaves. J. Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Gitelson, A.; Lang, M. Non-Destructive Determination of Chlorophyll Content of Leaves of a Green and an Aurea Mutant of Tobacco by Reflectance Measurements. J. Plant Physiol. 1996, 148, 483–493. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Blackburn, G.A. Quantifying chlorophylls and carotenoids at leaf and canopy scales: An evaluation of some hyperspectral approaches. Remote Sens. Environ. 1998, 66, 273–285. [Google Scholar] [CrossRef]
- Roujean, J.-L.; Breon, F.-M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Dawson, T.P.; Curran, P.J. Technical note A new technique for interpolating the reflectance red edge position. Int. J. Remote Sens. 1998, 19, 2133–2139. [Google Scholar] [CrossRef]
- Guyot, G.; Baret, F.; Major, D.J. High spectral resolution: Determination of spectral shifts between the red and the near infrared. Int. Arch. Photogramm. Remote Sens. 1988, 11, 750–760. [Google Scholar]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Yang, W.; Wang, K.; Zuo, W. Neighborhood Component Feature Selection for High-Dimensional Data. J. Comput. 2012, 7, 161–168. [Google Scholar] [CrossRef]
- Goldberger, J.; Hinton, G.E.; Roweis, S.T.; Salakhutdinov, R.R. Neighbourhood components analysis. In Advances in Neural Information Processing Systems; MIT Press: Cambridge, MA, USA, 2005; pp. 513–520. [Google Scholar]
- Torresani, L.; Lee, K.C. Large margin component analysis. In Advances in Neural Information Processing Systems; MIT Press: Cambridge, MA, USA, 2007; pp. 1385–1392. [Google Scholar]
- Raghu, S.; Sriraam, N. Classification of focal and non-focal EEG signals using neighborhood component analysis and machine learning algorithms. Expert Syst. Appl. 2018, 113, 18–32. [Google Scholar] [CrossRef]
- Melssen, W.; Wehrens, R.; Buydens, L. Supervised Kohonen networks for classification problems. Chemom. Intell. Lab. Syst. 2006, 83, 99–113. [Google Scholar] [CrossRef]
- Pantazi, X.E.; Moshou, D.; Oberti, R.; West, J.; Mouazen, A.M.; Bochtis, D. Detection of biotic and abiotic stresses in crops by using hierarchical self organizing classifiers. Precis. Agric. 2017, 18, 383–393. [Google Scholar] [CrossRef]
- Bishop, C.M. Pattern Recognition and Machine Learning; Springer: Berlin, Germany, 1995. [Google Scholar]
- Ballabio, D.; Vasighi, M.; Consonni, V.; Kompany-Zareh, M. Genetic Algorithms for architecture optimisation of Counter-Propagation Artificial Neural Networks. Chemom. Intell. Lab. Syst. 2010, 105, 56. [Google Scholar] [CrossRef]
- Fabelo, H.; Ortega, S.; Ravi, D.; Kiran, B.R.; Sosa, C.; Bulters, D.; Callico, G.; Bulstrode, H.; Szolna, A.; Pineiro, J.; et al. Spatio-spectral classification of hyperspectral images for brain cancer detection during surgical operations. PLoS ONE 2018, 13, e0193721. [Google Scholar] [CrossRef] [Green Version]
- Dovas, C.I.; Katis, N.I.; Avgelis, A.D. Multiplex detection of criniviruses associated with epidemics of a yellowing disease of tomato in Greece. Plant Dis. 2002, 86, 1345–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Gazala, I.S.; Sahoo, R.N.; Pandey, R.; Mandal, B.; Gupta, V.K.; Singh, R.; Sinha, P. Spectral reflectance pattern in soybean for assessing yellow mosaic disease. Indian J. Virol. 2013, 24, 242–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.L.; Philpot, W.D.; Norvell, W.A. Yellowness index: An application of spectral second derivatives to estimate chlorosis of leaves in stressed vegetation. Int. J. Remote Sens. 1999, 20, 3663–3675. [Google Scholar] [CrossRef]
- Kim, K.S.; Flores, E.M. Nuclear changes associated with Euphorbia mosaic virus transmitted by the whitefly. Phytopathology 1979, 69, 984. [Google Scholar] [CrossRef]
- Kalacska, M.; Sanchez-Azofeifa, G.A.; Rivard, B.; Calvo-Alvarado, J.C.; Quesada, M. Baseline assessment for environmental services payments from satellite imagery: A case study from Costa Rica and Mexico. J. Environ. Manag. 2008, 88, 348–359. [Google Scholar] [CrossRef]
- Hoque, E.; Hutzler, P.J.S. Spectral blue-shift of red edge monitors damage class of beech trees. Remote Sens. Environ. 1992, 39, 81. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Tang, Q.; Huang, W. Estimating chlorophyll content from hyperspectral vegetation indices: Modeling and validation. Agric. For. Meteorol. 2008, 148, 1230–1241. [Google Scholar] [CrossRef]
- Lu, J.; Ehsani, R.; Shi, Y.; Castro, A.I.; Wang, S. Detection of multi-tomato leaf diseases (late blight, target and bacterial spots) in different stages by using a spectral-based sensor. Sci. Rep. 2018, 8, 2793. [Google Scholar] [CrossRef] [Green Version]
- López-López, M.; Calderón, R.; González-Dugo, V.; Zarco-Tejada, P.; Fereres, E. Early detection and quantification of almond red leaf blotch using high-resolution hyperspectral and thermal imagery. Remote Sens. 2016, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.K.; Kim, M.K.; Kwak, H.R.; Choi, H.S.; Nam, M.; Choe, J.; Koi, B.; Han, S.J.; Kang, J.H.; Jung, C. Molecular dissection of distinct symptoms induced by tomato chlorosis virus and tomato yellow leaf curl virus based on comparative transcriptome analysis. Virology 2018, 516, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Devadas, R.; Lamb, D.W.; Simpfendorfer, S.; Backhouse, D. Evaluating ten spectral vegetation indices for identifying rust infection in individual wheat leaves. Precis. Agric. 2009, 10, 459–470. [Google Scholar] [CrossRef]
- Schor, N.; Bechar, A.; Ignat, T.; Dombrovsky, A.; Elad, Y.; Berman, S. Robotic Disease Detection in Greenhouses: Combined Detection of Powdery Mildew and Tomato Spotted Wilt Virus. IEEE Robot. Autom. Lett. 2016, 1, 354–360. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index Abbreviation | Index Name | Index Formula | Reference |
|---|---|---|---|
| NDVI | Normalized Difference Vegetation Index | [41] | |
| NSRI | NIR Shoulder Ratio Index | [42] | |
| NDRE | Normalized Difference Red Edge | [43] | |
| TVI | Triangular Vegetation Index | [44] | |
| ARI | Anthocyanin Reflectance Index | [45] | |
| CSI1 | Carter Stress Index | [46] | |
| GM1 | Gitelson and Merzylak Index 1 | [47] | |
| gNDVI | green Normalized Difference Vegetation Index | [48] | |
| LCI | Leaf Chlorophyll Index | [49] | |
| LIC1 | Lichtenhaler Index | [50] | |
| MCARI1 | Modified Chlorophyll Absorption Ratio Index 1 | [51] | |
| MCARI2 | Modified Chlorophyll Absorption Ratio Index 2 | [51] | |
| MTVI1 | Modified Triangular Vegetation Index 1 | [51] | |
| MTVI2 | Modified Triangular Vegetation Index 2 | [51] | |
| PSSRa | Pigment Specific Simple Ratio (Chl a) | [52] | |
| PSSRb | Pigment Specific Simple Ratio (Chl b) | [52] | |
| PSSRc | Pigment Specific Simple Ratio (Carotenoids) | [52] | |
| RDVI | Renormalized Difference Vegetation Index | [53] | |
| REIP | Red Edge Inflection Point | [54] | |
| REIP1 | Modified Red Edge Inflection Point | [55] | |
| SRCHLTOT | Simple Ratio of Total Chlorophyll Content | [48] | |
| VOG1 | Vogelmann Index 1 | [56] | |
| VOG2 | Vogelmann Index 2 | [56] | |
| VOG3 | Vogelmann Index 3 | [56] |
| Ct Value | Class Characteristics | Class Label |
|---|---|---|
| >43.5 | Control—Not infected | 1 |
| 43.5–30 | Very low infection—No symptoms and hardly detectable viral concentration | 2 |
| 30–25.5 | Low to mid infection—No symptoms and detectable viral concentration | 3 |
| 25.5–20 | Mid to high infection—First symptoms | 4 |
| Network Hyperparameters | Values |
|---|---|
| Training Epoch Number | 50, 100, 200, 300, 500, 600, 1000 |
| Self Organizing Map (SOM)/Hidden Layer Size | 4, 8, 10, 15, 20, 25, 30 |
| Learning Rate | 0.001, 0.005, 0.01, 0.02, 0.05 |
| Before Outlier Elimination | After Outlier Elimination | ||
|---|---|---|---|
| Vegetation Indices | Effective Wavelenghts (nm) | Vegetation Indices | Effective Wavelenghts (nm) |
| ARI PSSRa PSSRb REIP VOG1 | 402.2 405.5 408.9 412.2 415.6 425.7 429.0 449.2 553.0 563.0 566.4 676.4 736.2 865.4 | TVI ARI PSSRa PSSRb REIP SRCHLTOT VOG2 | 402.2 405.5 412.2 415.6 425.7 429.0 449.2 556.4 559.7 563.0 566.4 676.4 679.7 722.9 726.3 862.1 |
| Outliers | Features | Class Prediction Accuracy (%) | Overall | F1 Score | |||
|---|---|---|---|---|---|---|---|
| Healthy | ToCV1 | ToCV2 | ToCV3 | ||||
| Before Elimination | Effective Wavelengths (EW) | 87.6 | 82.6 | 86.2 | 88.0 | 86.1 | 0.861 |
| Vegetation Indices (VI) | 92.9 | 91.7 | 89.0 | 88.2 | 90.4 | 0.904 | |
| After Elimination | Effective Wavelengths (EW) | 99.8 | 99.8 | 100 | 100 | 99.8 | 0.998 |
| Vegetation Indices (VI) | 99.9 | 99.9 | 100 | 100 | 99.9 | 0.999 | |
| Outliers | Features | Class Prediction Accuracy (%) | Overall | F1 Score | |||
|---|---|---|---|---|---|---|---|
| Healthy | ToCV1 | ToCV2 | ToCV3 | ||||
| Before Elimination | EW | 91.4 | 92.8 | 91.2 | 90.7 | 91.6 | 0.915 |
| VI | 94.7 | 92.9 | 92.4 | 90.7 | 92.7 | 0.927 | |
| After Elimination | EW | 100 | 100 | 100 | 100 | 100 | 1 |
| VI | 100 | 100 | 100 | 100 | 100 | 1 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morellos, A.; Tziotzios, G.; Orfanidou, C.; Pantazi, X.E.; Sarantaris, C.; Maliogka, V.; Alexandridis, T.K.; Moshou, D. Non-Destructive Early Detection and Quantitative Severity Stage Classification of Tomato Chlorosis Virus (ToCV) Infection in Young Tomato Plants Using Vis–NIR Spectroscopy. Remote Sens. 2020, 12, 1920. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12121920
Morellos A, Tziotzios G, Orfanidou C, Pantazi XE, Sarantaris C, Maliogka V, Alexandridis TK, Moshou D. Non-Destructive Early Detection and Quantitative Severity Stage Classification of Tomato Chlorosis Virus (ToCV) Infection in Young Tomato Plants Using Vis–NIR Spectroscopy. Remote Sensing. 2020; 12(12):1920. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12121920
Chicago/Turabian StyleMorellos, Antonios, Georgios Tziotzios, Chrysoula Orfanidou, Xanthoula Eirini Pantazi, Christos Sarantaris, Varvara Maliogka, Thomas K. Alexandridis, and Dimitrios Moshou. 2020. "Non-Destructive Early Detection and Quantitative Severity Stage Classification of Tomato Chlorosis Virus (ToCV) Infection in Young Tomato Plants Using Vis–NIR Spectroscopy" Remote Sensing 12, no. 12: 1920. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12121920