1. Introduction
Recent research efforts in marine and climate disciplines highlight the effects that increasing atmospheric CO
2 concentrations are having on the Earth’s climate and in its control over the oceanic carbonate system and pH [
1]. As one of the greenhouse gases, its air–sea flux is a critical part of the climate system.
Atmospheric carbon dioxide (CO
2) has been increasing since the industrial revolution due to the burning of fossil fuels, cement production and land use change. While it is generally accepted that the ocean system annually absorbs up to 30% of the CO
2 released by burning fossil fuels [
2], it is not clear whether this ocean carbon ‘sink’ is increasing or decreasing, and considerable temporal and spatial inter-annual variation appears to occur [
3]. The flux of CO
2 between the atmosphere and the ocean (air–sea) is primarily controlled by conditions of the physical environment like wind speed, sea state and sea-surface temperature but also by biological processes taking place in the euphotic zone. Adding CO
2 to seawater disrupts the carbonate system, leading to an increase in free CO
2/carbonic acid and the bicarbonate ion concentration while decreasing the carbonate ion, pH and carbonate saturation states [
4]. All of these have implications for a range of biological and chemical processes like calcification, primary production and reproduction [
5,
6].
In shelf seas, CO
2 dynamics are highly variable in space and time [
7,
8] partly driven by the high primary productivity, leading to a large draw-down of dissolved inorganic carbon (DIC, the sum of CO
2, bicarbonate and carbonate ions) and consequent rise in pH [
9]. This is further complicated by the close link between benthic processes and the pelagic carbon cycle and alkalinity [
10] and riverine inputs of DIC and total alkalinity (TA) into coastal systems [
11].
In addition to these processes, diapycnal mixing across the thermocline drives a continuous change in surface CO
2 as a result of vertical fluxes of DIC and nutrients that stimulate primary production [
12]. The existence of near-surface temperature variations was identified by Ward et al. [
13] and their significance for air–sea CO
2 fluxes has been recently highlighted by Woolf et al. [
14].
Clearly, further understanding the CO2 pathways, sources, sinks, their respective budgets and their impacts on Earth’s climate system is essential for monitoring and projecting future climate scenarios.
In this work we evaluate the sensitivity of model derived CO
2 fluxes to physical and biological drivers in a shallow shelf sea location (L4) using the GOTM–ERSEM model [
15]. The use of a 1D model here is appropriate because in shallow shelf seas, the vertical mixing is mainly driven by the balance between tidal mixing and atmospheric fluxes, especially solar heating.
L4 (4 nm south of Plymouth, 50°15.00′N, 4°13.02′W, depth of 51 m) is a coastal station in the western English Channel where sustained observations have been carried out since the late 1980s and where variability in planktonic communities has been described extensively [
16]. In recent years, physical observations have intensified in the framework of the Western Channel Observatory [
17] run by the Plymouth Marine Laboratory (PML) in cooperation with the Marine Biological Association (MBA). The environmental and water conditions at L4 have been sampled weekly and physico–chemical and biological observations are freely available. Recently, a mid-ocean buoy has been installed at L4 [
18]. As in other shelf seas, physics at L4 are mainly the result of a balance between tidal mixing and atmospheric forcing and a seasonal temperature stratification appears when summer heating overcomes the tidal induced mixing. Nutrient dynamics and ecosystem dynamics are strongly dependent on this variability [
17]. There are numerous reports of strong intra-seasonal and inter-annual variability in the plankton production and community composition in the western English Channel and in other shelf seas e.g., [
16,
19,
20], all of which help to modulate and alter the air–sea flux of CO
2. Collectively, these data provide a substantial dataset to parameterize and evaluate the performance of coupled hydrodynamic–ecosystem models.
3. Results
The in situ observations used in this work have been extensively described in numerous papers (e.g., [
16,
17,
37,
45]) but we provide a brief overview here. The seasonality of nutrients show typical maximum values in January (8 μM m
−3 for Nitrate, 0.5 μM m
−3 Phosphate, 5 μM m
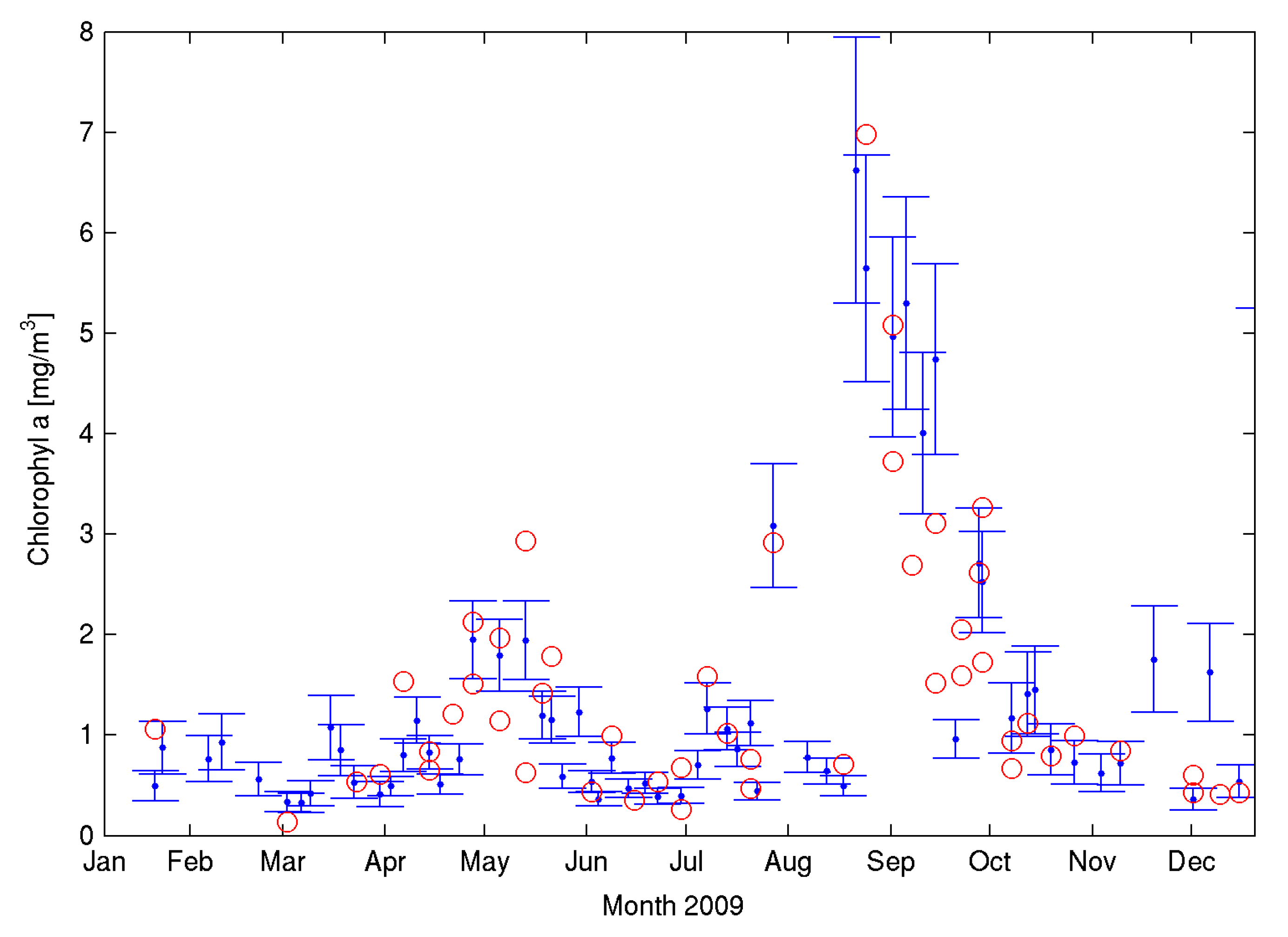
−3 Silicate) and minima after the spring bloom (from May) until it starts increasing again in September following the weakening of the thermal stratification. Chl-a also shows a marked seasonality at L4 (e.g.,
Figure 1). Two peaks are identified year on year in the data, one coincidental with the spring bloom (April–May) and one in September–October. The spring bloom is generally dominated by diatoms but with significant contributions from coccolitophores and phaeocystis. The analysis of 15 years of phytoplankton abundance at L4 [
16] has shown significant seasonal and inter-annual changes in abundance and composition although seasonality is the largest determinant of these changes. The authors found that the winter community seems relatively stable over the years. The spring period represents the largest change event in both composition and abundance while the summer period is the most susceptible to inter-annual changes in composition [
16]. Overall, flagellates are the dominant group representing 87% of the total phytoplankton cell abundance on average with the smaller cells (2–4 μm) accounting for 63% of the total abundance. In terms of biomass, they represent a small but significant proportion of the total biomass.
3.1. Reference Simulation; Experiment A
3.1.1. Physics
We have evaluated both the model capacity to reproduce the seasonal changes in the physical environment as well as sub-daily changes associated with the semi-diurnal tides. Initial calibration of the model and simulations to evaluate the sensitivity of the model to variations in forcing have been performed for the years 2009–2010, when data from the oceanographical buoy are available and can help to reduce uncertainty in the meteorological forcing. In August 2010, we deployed a bottom mounted Teledyne RD Instruments 600 kHz Broadband Acoustic Doppler Current Profiler (ADCP) at L4 for 20 days [
61].
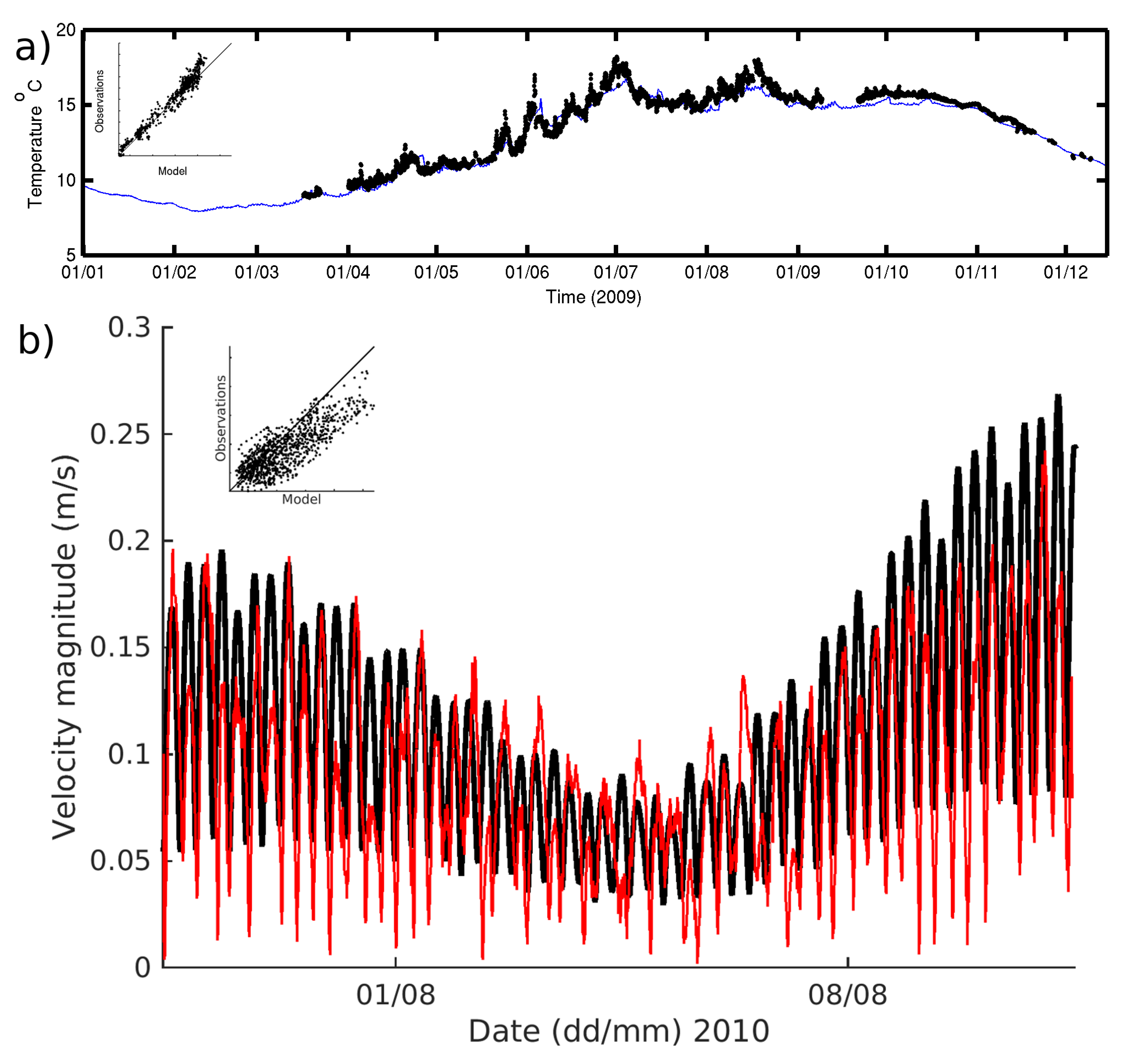
The seasonality in hydrography is well reproduced by the model (
Figure 2a). This arises partly from the fact that we are using a basic nudging technique to relax the model temperature (T) and salinity (S) to the observed CTD profiles with a relaxation time of two weeks. The correlation for near-surface (1.5 m) temperature between the independent observations from the L4 monitoring buoy and the model was larger than 0.9 and significant at the 99.9% confidence level (full statistics can be found in
Table A4).
The ability of the model to reproduce the velocity structure was evaluated during a simulation of the August ADCP deployment. The results indicate that the model setup reproduces the barotropic component of the tidal velocity well considering the 1D approach. The correlation between the depth mean averaged velocity between 2 and 30 m was 0.7 at the 99.9% confidence level. The dominant E–W component is better resolved albeit with a small overestimation of the flood peak velocity (correlation of 0.9 at 99.9% confidence level). The N–S component shows a larger amplitude in magnitude than the observations which resulted in lower correlations of 0.7. The velocities closest to the bottom (averaged between 1 and 2 m above the seabed, (
Figure 2b)) have a better agreement between model and observations with an overall correlation of 0.8 for the velocity magnitude. As it is a tidally dominated station, this is key to ensuring a realistic level of mixing is imposed to the biological model.
3.1.2. Biogeochemistry
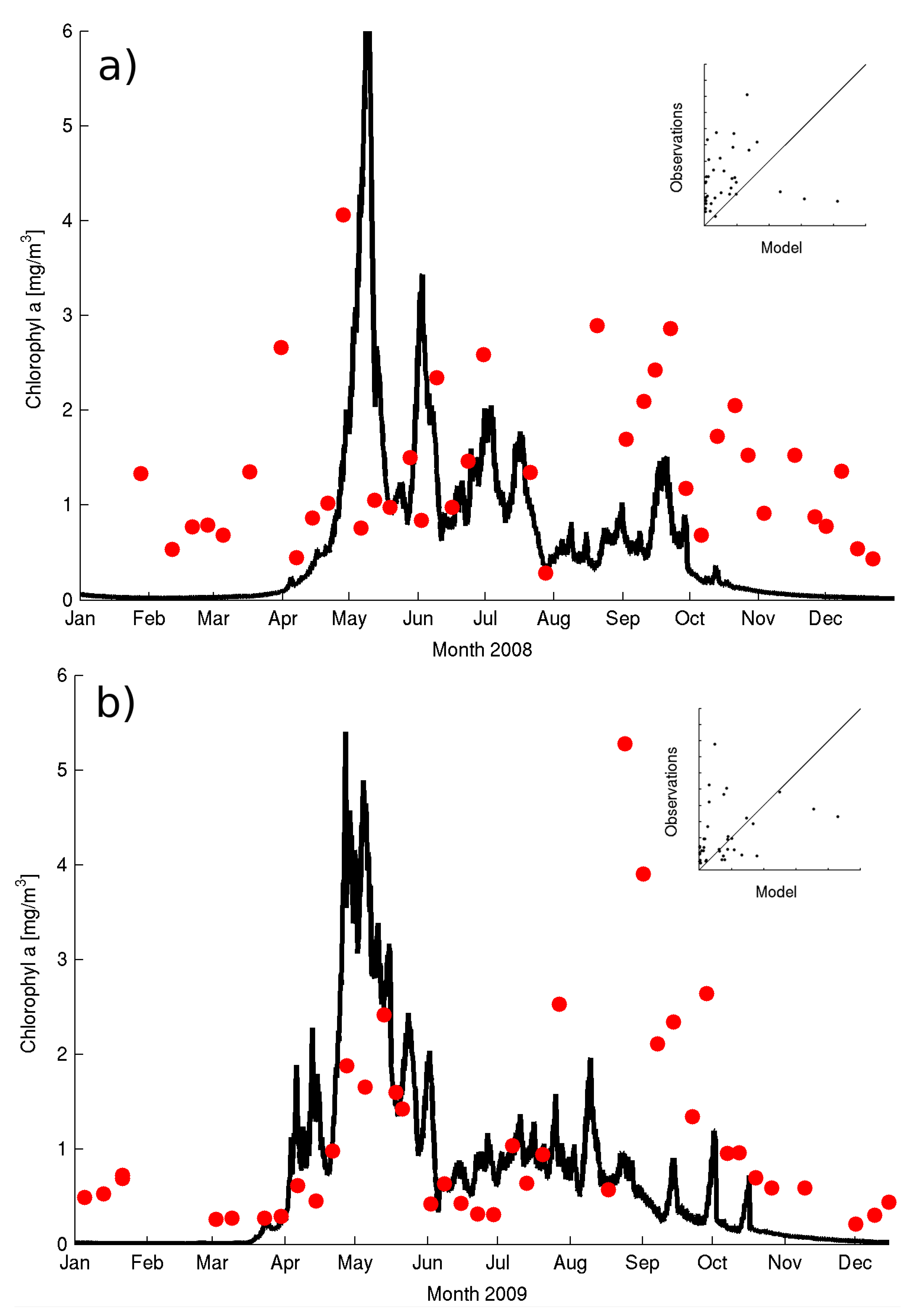
The model output at six hourly intervals in 2008–2009 at the sampling depth of 10 m (
Figure 3a,b) shows the model correctly reproduces the approximate timing and duration of the spring bloom. The model indicates a short sharp spring bloom in 2008 (
Figure 3a) with subsequent short-lived blooms during June–July in agreement with the available observations. The spring bloom in 2009 is by contrast longer (
Figure 3b) although the model overestimates the chl-a concentrations. Nonetheless, the observations suggest a substantial overestimation of diatom biomass in 2008 (
Figure 4b) but the correct order of magnitude in May 2009 (
Figure 4b). The model underestimates total chl-a from January to mid-April and from September to December in 2008 and 2009. The main discrepancy with the observations is the late dinoflagellate bloom (not shown) which the model underestimate in both years although the miss-match is most evident in 2009 (
Figure 3b).
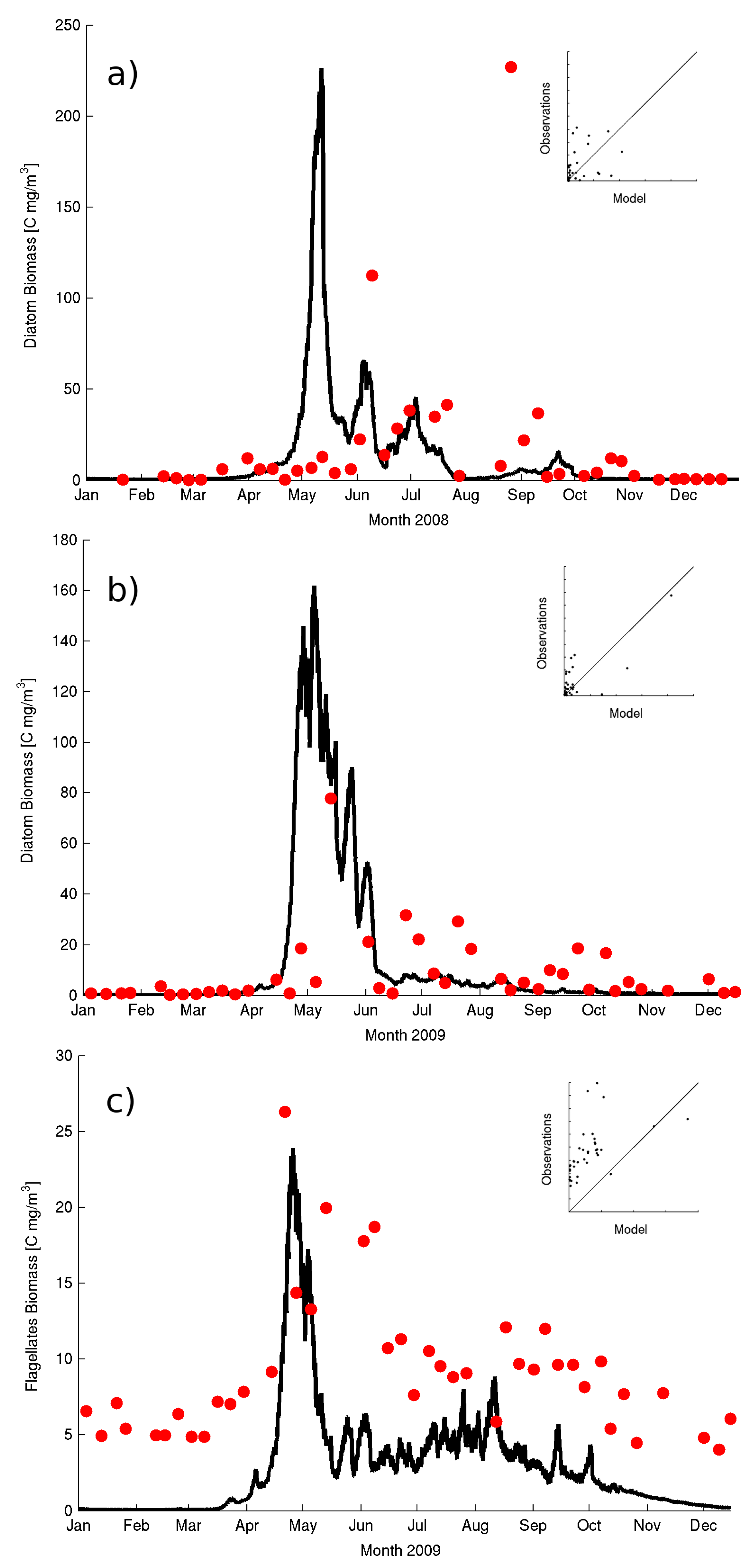
The main functional group responsible for the winter surface chl-a and spring bloom is diatoms (
Figure 4a,b) with a significant contribution from flagellates (
Figure 4c) which ranges from maximum of 40% during the spring bloom to 10–20% during the summer months.
The modeled spring bloom has knock-on effects in the simulated plankton succession. The collapse of the spring bloom generates substantial concentrations of ammonium which in turn triggers the onset of a succession of summer picoplankton blooms. The small pico-phytoplankton functional group dominates both chl-a and net production. Their higher affinity for ammonium and faster growth enables them to out compete both diatoms and flagellates and they are responsible for the disappearance of surface nutrients. The short-lived blooms in
Figure 3 are both a consequence of the variability in the observed irradiance and the model dynamics of nutrient–prey–predator cycles. The largest chl-a values are however not at the surface but in the well-developed subsurface chl-a maximum [
62].
Total biomass, here considered to be the sum of diatoms, flagellates and dinoflagellates, agrees well with the observations (
Table 1) from April to August with the largest discrepancy observed at the time of a mono-specific dinoflagellate bloom in early September 2009. These mono-specific blooms are one of the best-known weaknesses in biogeochemical models that embrace a functional group approach [
63]. Despite the model shortcomings, we still resolve the observed total biomass seasonality with correlations larger than 0.7 in both years (see
Table 1). The daily and depth integrated net primary production (
Figure 5) has a lower correlation of 0.6 (see
Table A1).
Nonetheless, smaller blooms of diatoms are still reproduced in the model simulations with a good agreement with the observations (
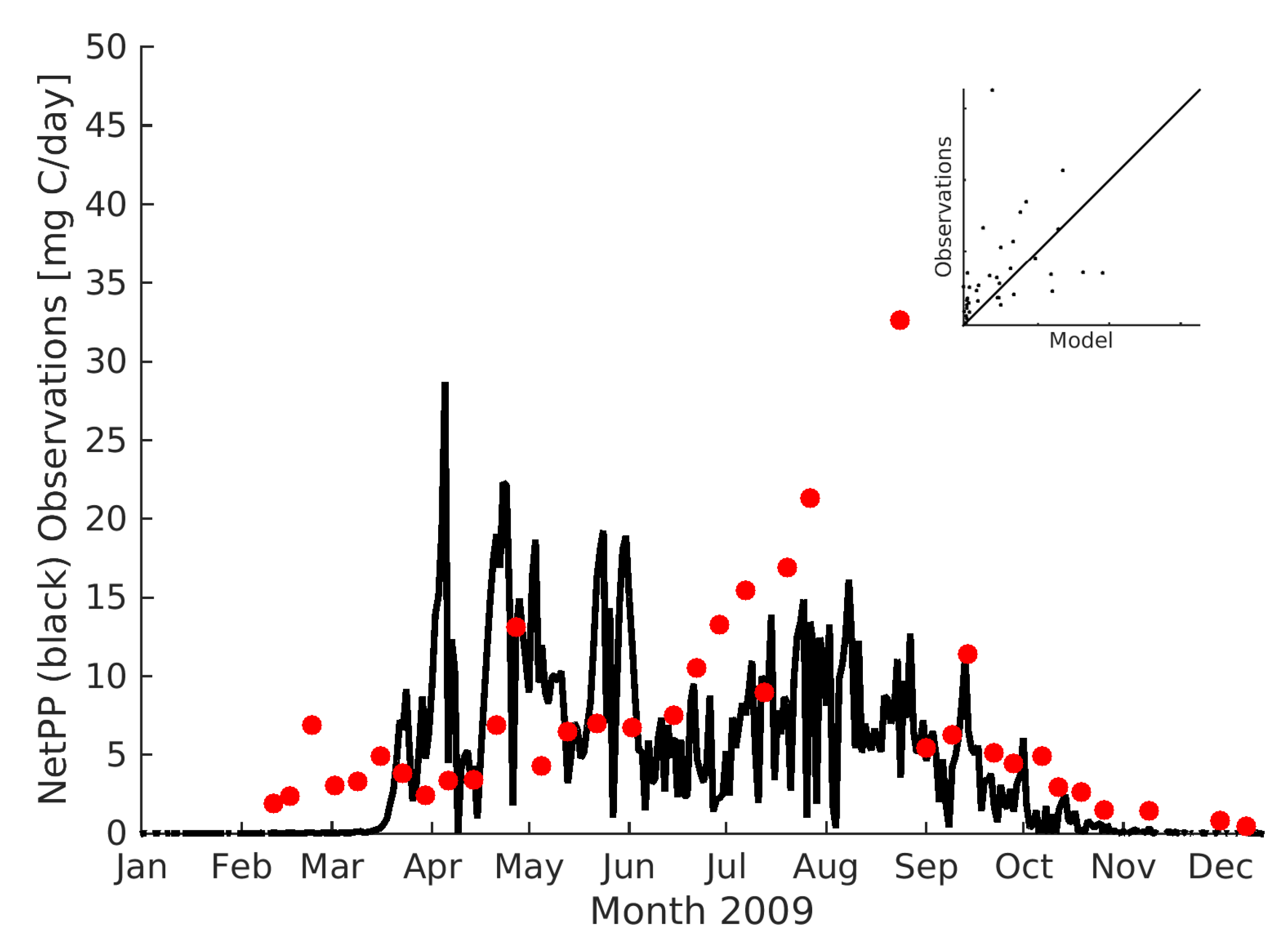
Figure 4a). The observed phytoplankton production (PP) at 10m indicates a strong seasonality with maximum values during July August (e.g.,
Figure 5). In contrast, the model suggests a high PP during both the spring bloom and the summer months of July and August.
All the measured surface nutrients display a strong seasonal signal which is captured by the model (e.g.,
Figure 6a,b) with the exception of ammonium (
Table A3). The main discrepancies relate to the rapid consumption of nutrients (silicate, nitrate and phosphate) during the spring bloom and the low nutrient concentrations during the summer period (mid-May to early September). The observations suggest that diatoms start drawing dissolved silicate from February (
Figure 6a) decreasing steadily until mid-April followed by a sharp drop to limiting levels by mid-late June. The early slow growth is likely controlled by light limitation and possibly by weak predation pressures. The sharp end of the bloom in May is potentially the result of predation from mesozooplankton. Light is a key driver of the pelagic ecosystem which during the winter period is dominated by the natural low levels at this northern latitude and the local re-suspension of benthic sediments, a process not resolved in the present GOTM–ERSEM implementation. Our hypothesis is that light limitation is responsible for the overestimation in the modeled biomass during the spring bloom with respect to the observations.
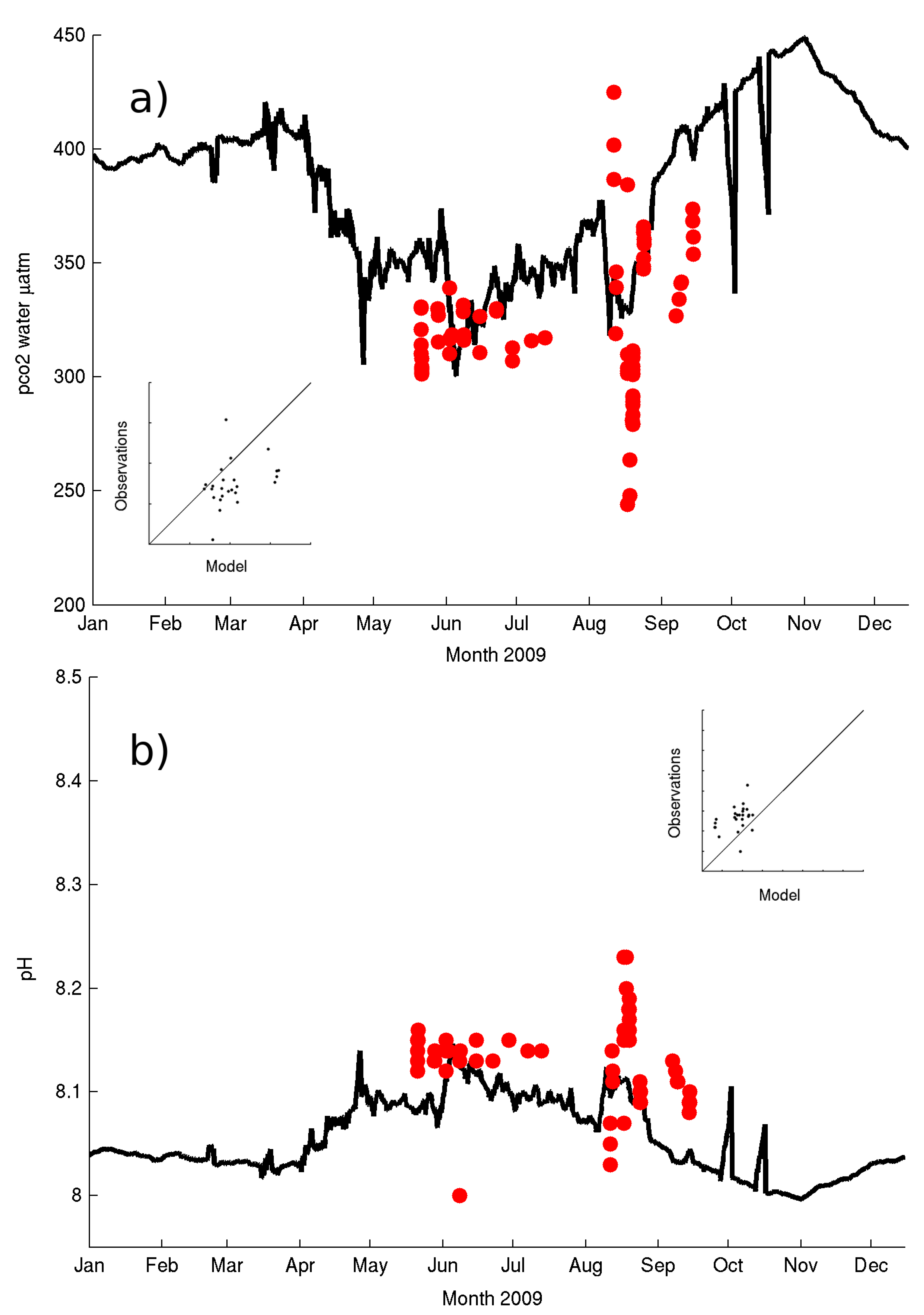
The evolution of the observed variables related to the carbonate system at the surface (namely pH and CO
2 partial pressure [pCO
2] ) follow the available data for 2008 and 2009 although the model underestimates the large daily fluctuations that can be seen in the data (
Figure 7a,b). The seasonal signal described in Kitidis et al. [
45] of increased pCO
2 and low pH in winter and autumn (October to March) and low pCO
2 and high pH during summer and spring (April to September) is well reproduced by the model in all years. The range in the observed values is also well captured by the model (~200 μatm for pCO
2 and ~0.15 pH units). Both model and observations show increased variability during the summer/stratified periods strongly coupled to phytoplankton blooms with timescales of 1–3 weeks.
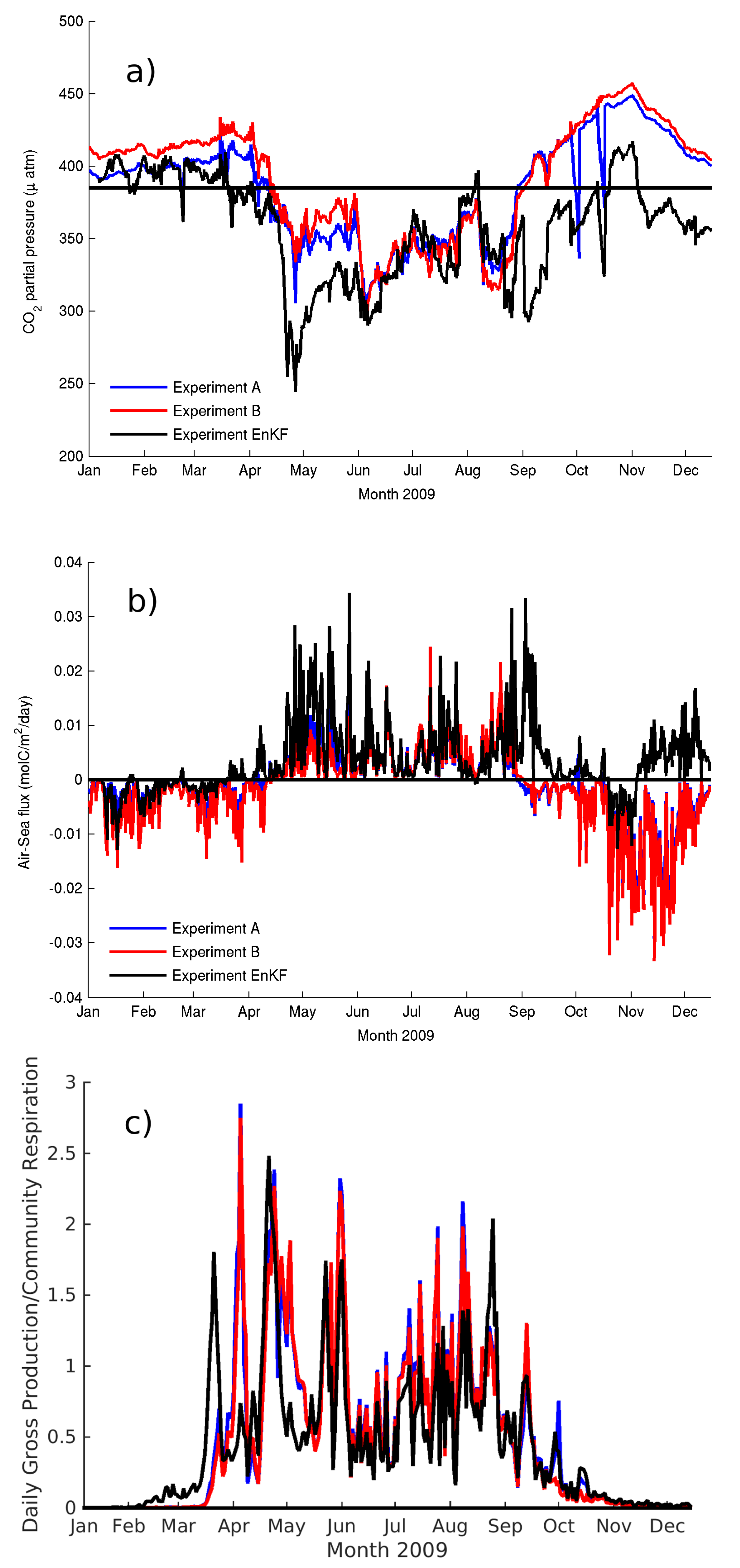
3.2. Enhance Surface Resolution; Experiment B
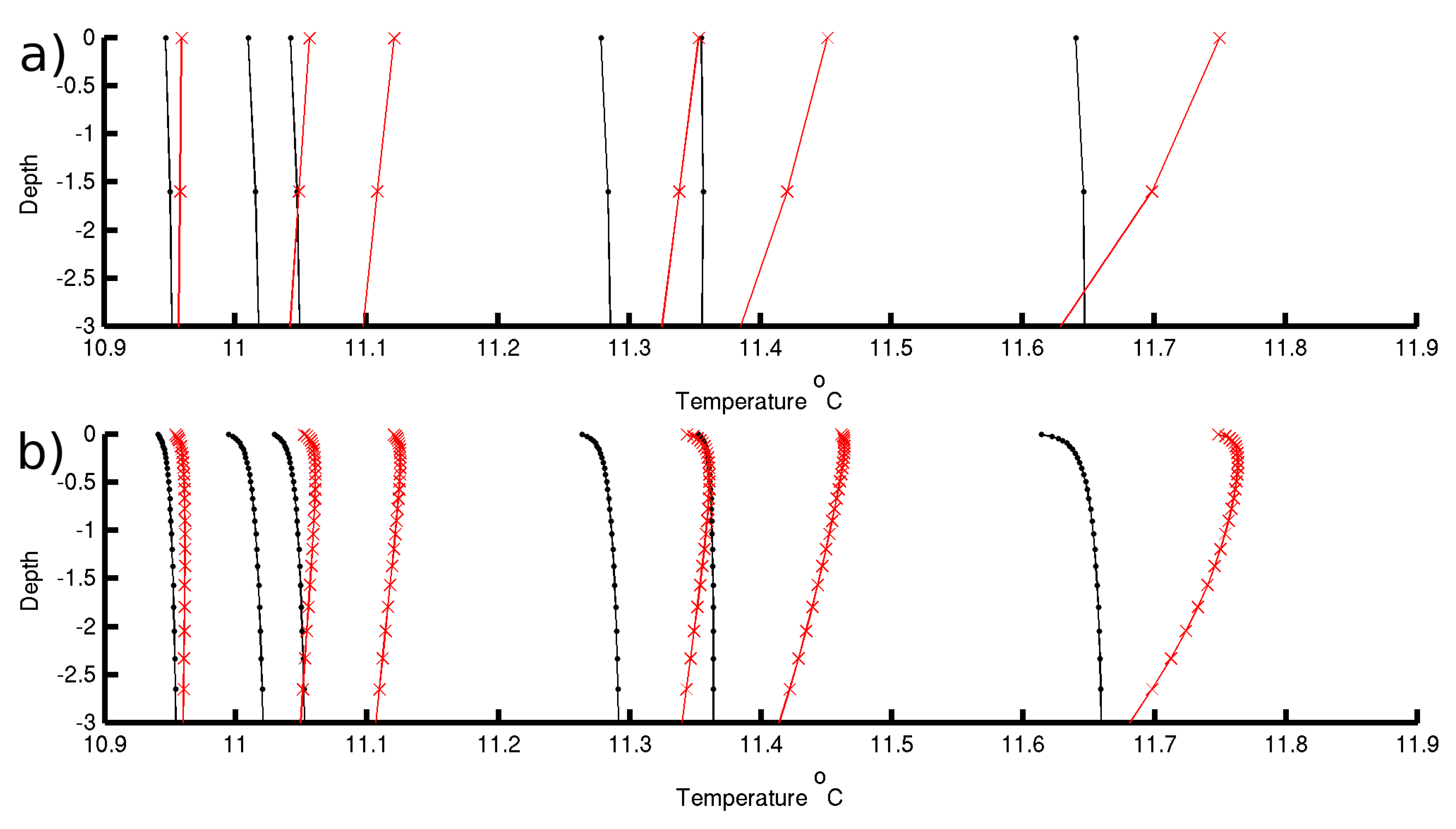
To evaluate the effect of resolving the diurnal cycle of the near-surface temperature on the air–sea fluxes we modified the baseline experiment by changing the vertical setup. The vertical grid is enhanced at the surface, and the resolution increased logarithmically from 2 cm at the surface to 1 m at 10 m deep. The water column interior (between 10 m deep and 35 m deep) has the same nominal resolution of the reference experiment (1.6 m). This setup resolves the diurnal cycle of temperature associated with the near-surface layer (e.g.,
Figure 8b) [
64]. During effective heat loss at the sea surface from high winds or cooler air temperature than sea-surface temperature the shallowest ∼20 cm have a negative temperature gradient both during day and night time (
Figure 8b) that was not resolved in Experiment A (
Figure 8a). The absorption of incoming solar radiation takes place over the top several meters of the water column, but heat is released right at the surface which causes the very surface to cool. During daytime, a subsurface temperature maximum develops between 30 and 150 cm (80 cm on average) from the surface. The largest differences between the two experiments concentrate on the top 3 m, below which depth the root mean square error (RMSE) decreases to 50% of the surface value (RMSE = 0.17 °C). The RMSE decrease flattens at 12 m and remains at 10% of the surface value to the bottom.
The development of the very near-surface gradient results in changes to the amplitude of the daily fluctuations of temperature such that the night–day range reduces in Experiment B, making night time minima warmer and day time maxima cooler than in Experiment A as the near-surface gradients limit the exchange of heat with the atmosphere.
pCO2 shows a similar night–day evolution to the temperature profiles with a consistent negative gradient in the top 20 cm and the development of a daytime subsurface maximum coincidental with the temperature maximum.
Experiment B showed that the carbonate system model variables are sensitive to the increased resolution near the surface. The comparison between experiments A and B (
Table 1) shows the largest improvements in 2008, in particular for pH, pCO
2 and DIC (a 10%, 19% and 18% improvement respectively,
Table 1). The effect on the air–sea flux is largest and it is apparent throughout the year save the most stratified periods between June and August. The impact is important and results in a 70% to 100% increase in the net CO
2 uptake at L4. The modeled integrated annual CO
2 flux was 1.8 mo C/m
2/year in Experiment A for 2008 increasing to an integrated annual CO
2 flux of 2.8 mo C/m
2/year in Experiment B for the same year. A similar result is obtained for 2009 with a change from net CO
2 source of 0.3 mo C/m
2/year in Experiment A to net CO
2 source of 0.6 mo C/m
2/year in Experiment B.
3.3. Evaluation of Surface Slicks; Experiment C
Experiment C involved a parameterization of the effects of primary production on the CO
2 transfer velocity (
). This was introduced through a scaling
with the gross primary production excreted by the phytoplankton as a result of its metabolic activity. In practice,
was reduced to a maximum of 70% of its wind dependent value (calculated following Nightingale et al. [
65]) when the gross productivity surpasses the 30% highest values modeled at L4 in the years 2008–2009. The results are summarized in
Table 1 and suggest a negligible impact when compared to Experiment A. The integrated annual CO
2 flux supports these results with changes smaller than 1% for both 2008 and 2009 years.
3.4. Evaluation of Data Assimilation; Experiment D
Results from Experiment D show that assimilation of EO chl-a improves some of the model variables but it also deteriorates bulk variables like total biomass (
Table 1 and
Table A1 in
Appendix A). As expected from previous work [
34] the forecast step always shows a larger variance or ensemble spread than after the analysis step suggesting a reduction in ensemble variance as a result. The modeled mixed layer chl-a (
Figure 9a) shows an improved agreement with observations (
Table 1) but the model fails to capture the spring bloom correctly. In particular, the model solution constructed as the mean of all the model ensembles overestimates chl-a in March and end of April while the overestimation during the observed spring bloom in May improves. The summer evolution of chl-a is better resolved as is the increase seen in September (
Figure 9a).
The improvement seen in chl-a do not however translate to a better fit in modeled total biomass, and the assimilation of chl-a degrades the model solution as a result of the large miss-representation of the spring bloom (not shown). Indeed, when the results between 15 March and 15 April are removed, the correlation for total biomass increases to 0.61 at the 95% significance. One effect of the assimilation is the overestimation of all the nutrient variables during the spring bloom (not shown). This reflect the inability of the model to lower the chl-a field without increasing the nutrient values during that period. The discrepancies and excess nutrients are however corrected during the summer period and have a small impact on the overall correlation with observations (
Table 1).
It is, however, noteworthy that while the correlations with observations of all variables in the carbonate system (pH, DIC and pCO
2,
Table 1) do not improve, the bias or RMSE is significantly reduced (
Table A4). The ensemble variance was reduced in the modeled chl-a as a result of the assimilation but it is apparent in
Figure 9b that this is not the case for pCO
2 or DIC and pH (not shown), although after the initial increase during the first three months of the experiment, it is also true that it did not increase monotonically.
4. Discussion
4.1. Ecosystem Models Predictive Capabilities of CO2
We have shown that a 1D approach using GOTM–ERSEM captures the seasonality of the pelagic ecosystem at L4, including the variables driving the air–sea exchange of CO
2 (e.g., year 2008
Table 1). The results from year 2009 (
Table 1) suggest that the model’s ability to simulate the total biomass and nutrients is not a sufficient condition to resolve the surface evolution of DIC and pCO
2. This follows the poorer simulation of chl-a in 2009 and suggests that an adequate simulation of chl-a and the associated plankton production and respiration rates is critical for a better estimation of the CO
2 dynamics. The seasonality and range of simulated DIC and pCO
2 agree with recent observations [
45] and suggest both a physical and biological control. The high production during the spring and summer periods (
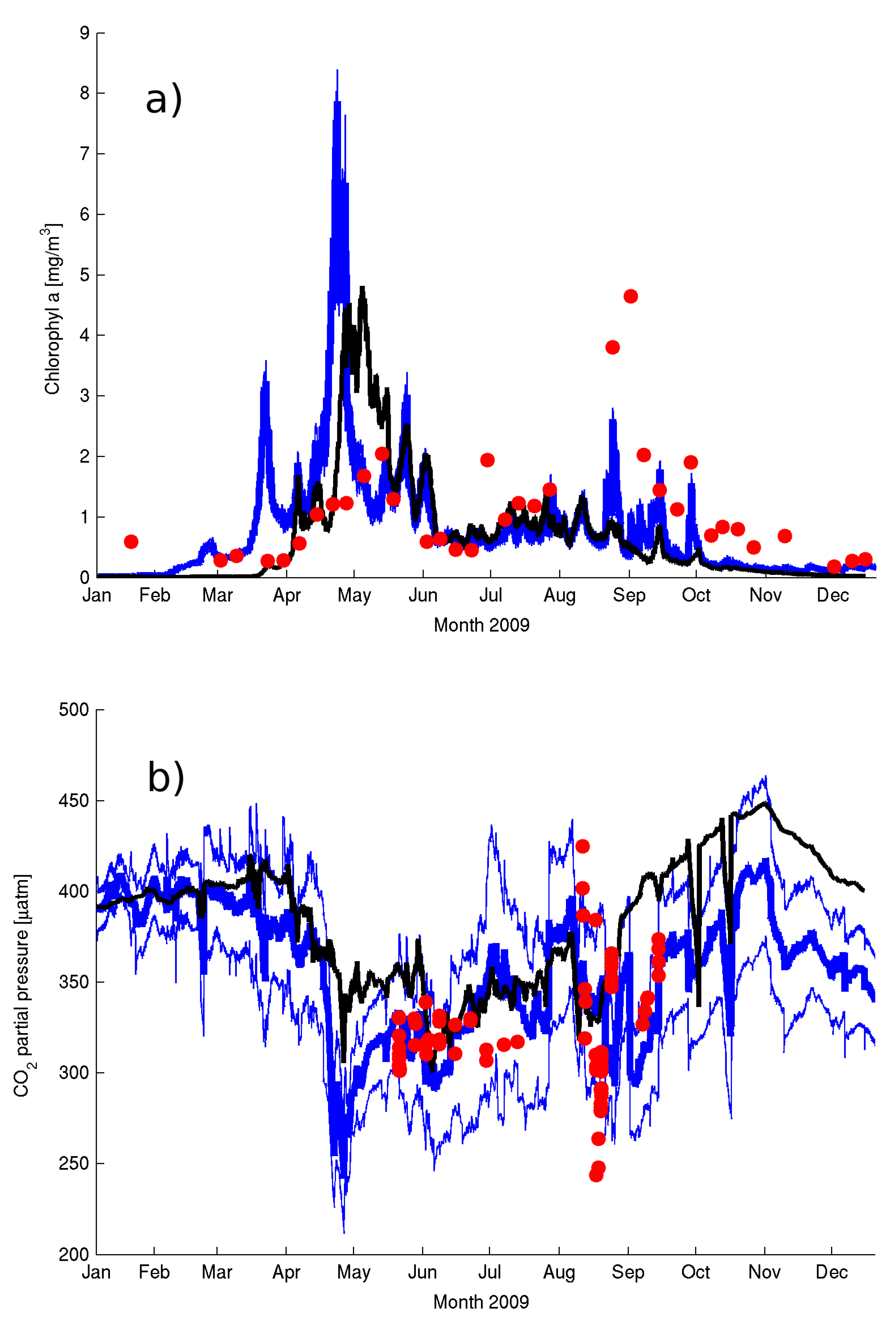
Figure 5) produces undersaturation of CO
2 (
Figure 10a) resulting in a net sink of CO
2 into the waters. It is worth noting that the short-term variability in air–sea CO
2 fluxes is as high as the seasonal signal (
Figure 9a) and that the summer blooms can drive a similar exchange to the longer spring bloom.
Despite the model’s ability to capture the seasonal cycle on all the variables of the carbonate system the model seems to underestimate the short-term variability seen in the data (
Figure 9b). This can be related to the coarse temporal resolution of the wind forcing (six hourly) (e.g., [
64]) and the effect of lateral advection which is intrinsic in the data and not resolved in the model. At L4, variations in DIC drive the observed variability in the carbonate system and thus pCO
2 as total alkalinity has very low variability [
45]. The model solution suggests that L4 can be considered a neutral site with respect to CO
2 fluxes or a small source (
Table 2) in contrast to the results reported by Kitidis et al. [
45].
4.2. Sensitivity of Modeled CO2 to SST Diurnal Cycle
The increased vertical resolution near the surface in Experiment B enabled the simulation of the diurnal cycle in SST [
64] which overall had a small impact on the ability of GOTM–ERSEM to simulate the bulk properties of the pelagic ecosystem at L4 as observed at 10 m (
Table 1) but a large impact on the air–sea fluxes of CO
2 (
Figure 10a,b). The comparison with SST measured at the L4 buoy yielded a small decrease in correlation (from 0.94 to 0.93 at the 99.9% confidence level) and a small decrease in the bias (see
Table A4). Experiment B did however improve the simulation of pCO
2, DIC and pH in 2008. The largest differences in the carbonate system between experiments A and B in 2009 concentrated in the first 5 months (January to May,
Figure 10a) and in Experiment B resulted in an increased supersaturation and resulting flux to the atmosphere from January to March. The amplitude of the daily variations in the fluxes remained relatively unchanged from June to December (
Figure 10b) driven primarily by the daily fluctuations in productivity. The difference was largest during the low productivity months of January to March and it was partly responsible for the change in value of the net flux estimates.
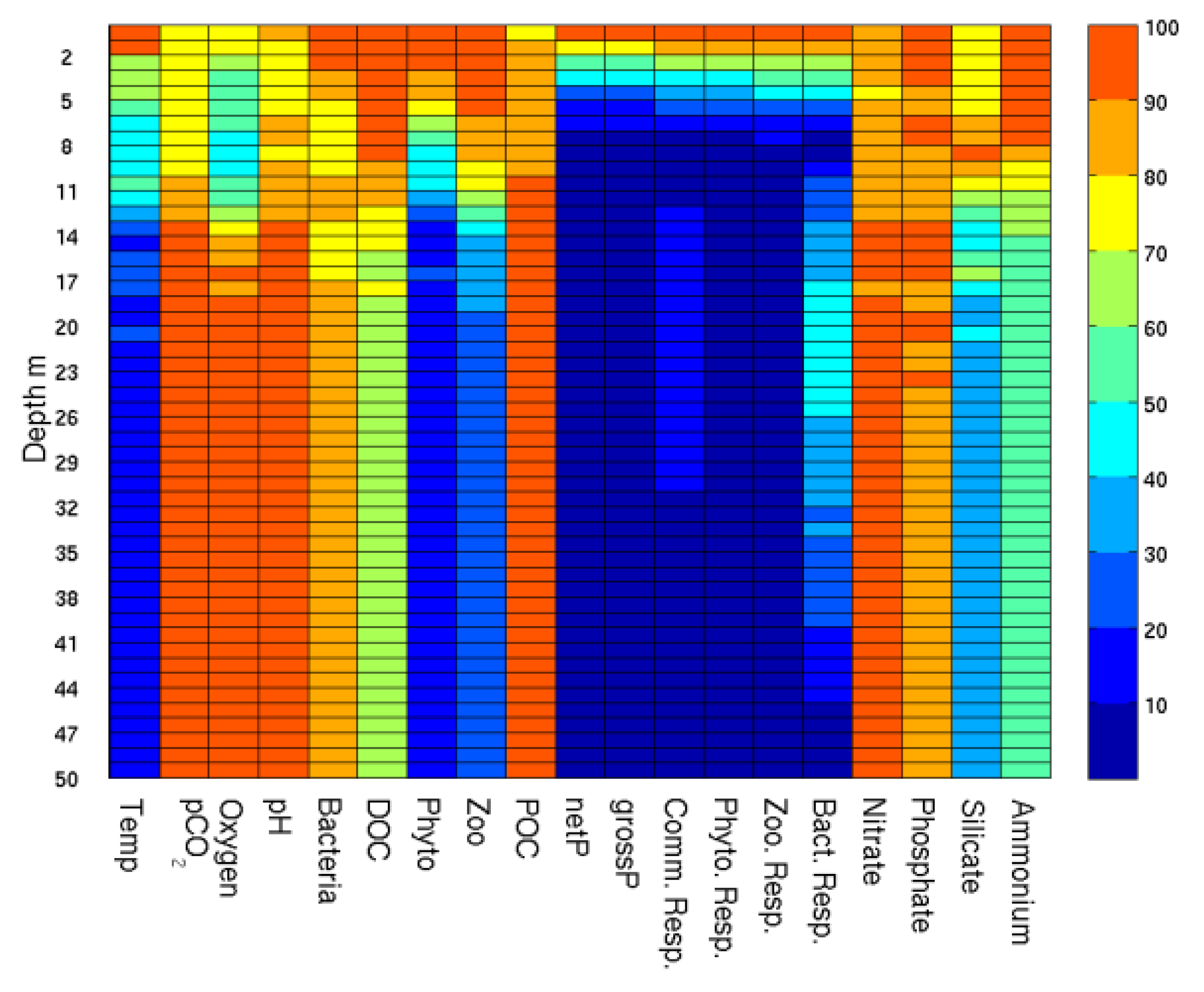
The changes with respect to the reference experiment (A) are mostly located in the top 10 m and decrease with depth in a significant number of variables (
Figure 11). These include rate variables like respiration and production as well as state model variables like biomass of plankton and zooplankton. These differences will be discussed in relation to three periods in 2009 defined as a winter non-growing season (January to March with a consistent gross production to community respiration (G/R) ratio less than one,
Figure 10c), the summer growing season from April to August (G/R averaging one or above) and the autumn non-growing season with G/R again less than one.
Most of the variables evaluated changed between 0.1% to 6% except for gross production and community respiration that showed increases between 26 and 34% in all seasons. Experiment B displayed a small increase in temperature in the top 10m throughout the year (0.1%) increasing to 0.5% during the growing season. Total detritus (the sum of the three classes in ERSEM, small, medium and large) decreased in the winter (4%), while on averaged did not change during summer and decreased by 5% in the autumn in Experiment B. Irradiance levels also increased in Experiment B (5% in all seasons) in response to a decrease in biomass of 2% in winter and 6% in autumn).
In general, though, pCO2 increased in Experiment B in both growing and non-growing season by 2%. During the non-growing season, this translates into an increase release of CO2 to the atmosphere in both 2008 and 2009. During the growing season, there is no one specific trend, as the CO2 exchange with the atmosphere depends on both the biological uptake of CO2 as well as the wind. During the two years evaluated, in 2009 there was no overall change while in 2008, there was a larger release of CO2 in Experiment B.
4.3. Sensitivity of Modeled CO2 to Surface Surfactants
The reduction of the gas transfer velocity (
) as a function of sea-surface surfactants had a very small impact on the integrated annual CO
2 flux (
Table 2) and bulk ecosystem variables (
Table 1) in our modeled site compared to the results of experiments B and D (
Section 3.2 and
Section 3.4). This is both a consequence of our choice of parameterization (
Section 3.3) of the production of surfactant material and the environmental characteristics of our site. The large variability in phytoplankton gross production (similar to phytoplankton net production in
Figure 5) implies that the reduction of
is only active during a small proportion of the time (<5% during 2008–2009) between April–September and results in small reductions of
(<1%). The high average wind speed (~8
observed at L4 in 2008–2009 also limits the impact of the surfactants on the annual integrated fluxes. This contrasts with the findings of Pereira et al. [
52] who reported an overall 9% reduction in the net CO
2 flux integrated over the entire Atlantic Ocean basin in 2014. Nonetheless, their analysis showed that at our latitude (although no estimates are presented for the UK shelf) the decreased surfactant suppression was consistently less than 5% except for July–September when it increased to 10%.
4.4. Sensitivity of Modeled CO2 to the Assimilation of chl-a
The assimilation of chl-a had a significant impact in the carbonate system but mostly by reducing the bias in DIC, pH or pCO
2. The correlations deteriorated for all three variables but the concentration of the observations around the May to September months precludes a definitive assessment of the effect of surface chl-a assimilation on simulating the seasonality of the carbonate system. Experiment D also corrected the simulation of chl-a and the nutrients silicate and phosphate (
Table 1).
Similar to Experiment B, Experiment D had a significant impact on the top 10 m averaged G and R, particularly during the winter (G increased 450%, R 20%) and in autumn (G increased 60% R 23%). Off all the variables evaluated, only plankton biomass had similar changes with a 900% increase in winter compared to 70% increase in the autumn. The second largest corrections corresponded to the zooplankton biomass, which decreased in winter (28%) and in summer (10%) while it increased in autumn (53%). Generally, the growing season saw more modest changes across all variables between 3 and 12%. It is worth remembering that although the percentage increases are significant they represent a small number as they correspond to the non-growing seasons (winter in particular) when biomass levels and carbon uptake and respired are very small. This is a further indication that winter ecosystem dynamics and autumn ones are the least well captured by this 1D model approach.
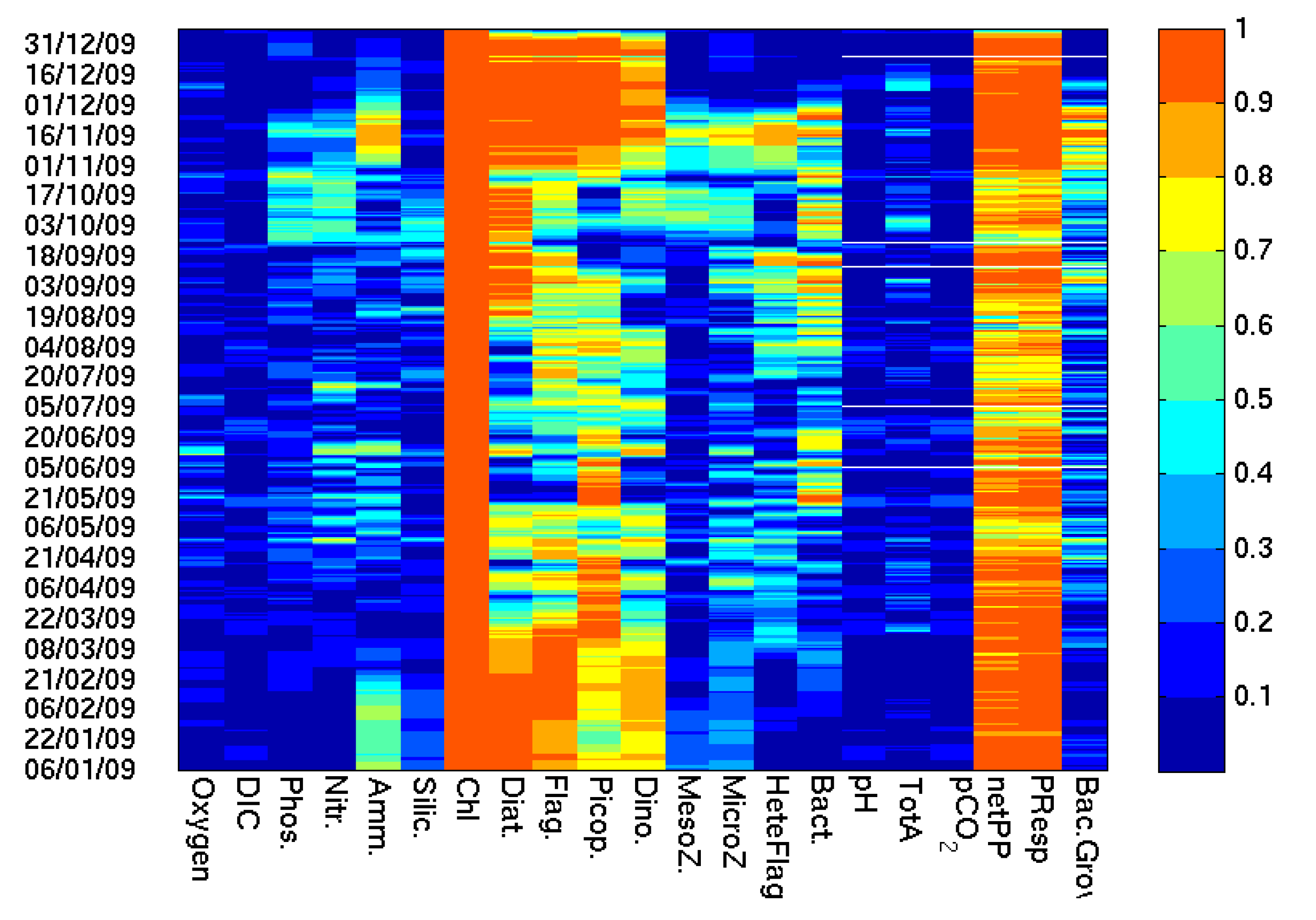
One way of visualizing how the information associated with chl-a is spread across all other model variables during the analysis step is to calculate the time evolution of the cross-correlation matrix among all of the ensemble members (
Figure 12). Chl-a is generally always correlated with model estimates of phytoplankton production and respiration and its correlation with the biomass of the functional types (including phytoplankton and zooplankton) varies in time. For example, chl-a is highly correlated with diatoms during the spring bloom, while the correlation with the picoplankton increases during the short-lived blooms of this group during the summer period. Equally, the correlations between chl-a and DIC, pH or pCO
2 are relatively low and indicates strong non-linear dynamics and feedback between chl-a or biomass and the carbonate chemistry. This suggest that direct corrections to these variables were relatively small in the EnKF analysis. Assimilation of chl-a indirectly corrected these variables in the forecast step, through the re-initialization of the other model variables in the analysis. The low correlations also explain why we do not see a reduction in the ensemble spread of these variables (e.g.,
Figure 9b) The corrections to near-surface pCO
2 (
Figure 10b) resulting from the assimilation of chl-a were significant across the seasonal cycle except for June to August when all experiments agree in their simulation of chl-a and primary production. The corrections were as high as 100 μatm and tended to reduce the values of near-surface pCO
2 resulting in net sink of CO
2 of 1.3 molCm
−2 year
−1 in agreement with the estimates of Kitidis et al. [
45]. The largest correction with a significant impact on the annual estimates relates to the primary production corrections in September to December.
5. Conclusions
We have shown that the 1D GOTM–ERSEM model has skill in simulating both the pelagic ecosystem and the variables related to the dynamics of CO2 at the coastal station L4, although the skill varies from year to year. Resolving the near-surface temperature gradients induced by the diurnal heating cycle (Experiment B) has the most efficient (lowest computational cost and skill requirements) impact at short, seasonal and annual time scales and results in improvements across the evaluated variables that are comparable to the results from the assimilation of chl-a Experiment (D). The correlation of carbonate system variables (DIC, pCO2 and pH) improve by up to 19% while annually integrated values of CO2 exchange with the atmosphere changed by up to 50% with respect to the reference simulation, increasing the net source nature of L4. At the L4 site, the introduction of a dependence of the CO2 gas transfer velocity on gross production as a proxy for the production of surface slicks (Experiment C) has a non-significant impact on the dynamics of CO2, including the air–sea annual exchange estimates.
The assimilation of surface chl-a (Experiment D) has an overall mixed effect on the ability of the model to reproduce the observations. As expected the simulation of chl-a improves and while the correlation for nutrients do not significantly change, their bias decreases. The effect on the carbonate system variables is similar; the bias decreases for DIC and pH but so do the correlations. The largest impact of assimilation is on the net air–sea gas exchange where the results indicate the potential for L4 to be a weak net source of carbon to the Atmosphere (1.3 ± 1.7 molCm−2 year−1). That chl-a assimilation can degrade the simulation of some variables possibly relates to the requirement for succinct parameter changes to successfully simulate the specific years considered in this work.
The results of the three sensitivity experiments indicate that resolving the temperature profile near the surface appears the most important aspect of those investigated here for a skilled simulation of the biogeochemistry at L4. This result indicates that this finer vertical scale should be included in future shelf sea models used in air–sea flux modeling studies.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











