
Landscape Structure of Woody Cover Patches for Endangered Ocelots in Southern Texas

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
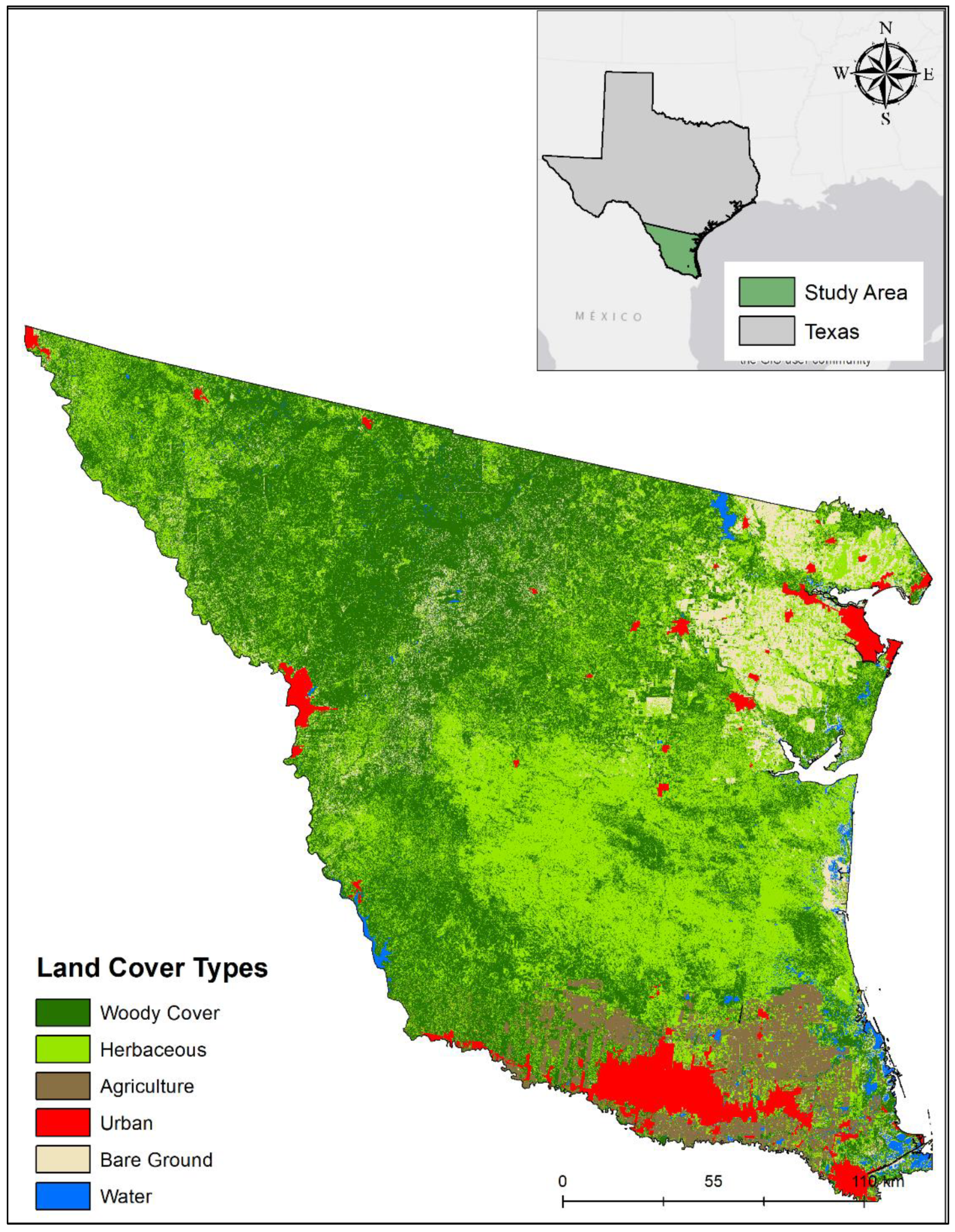
2.1. Study Area
2.2. Ocelot Spatial Data
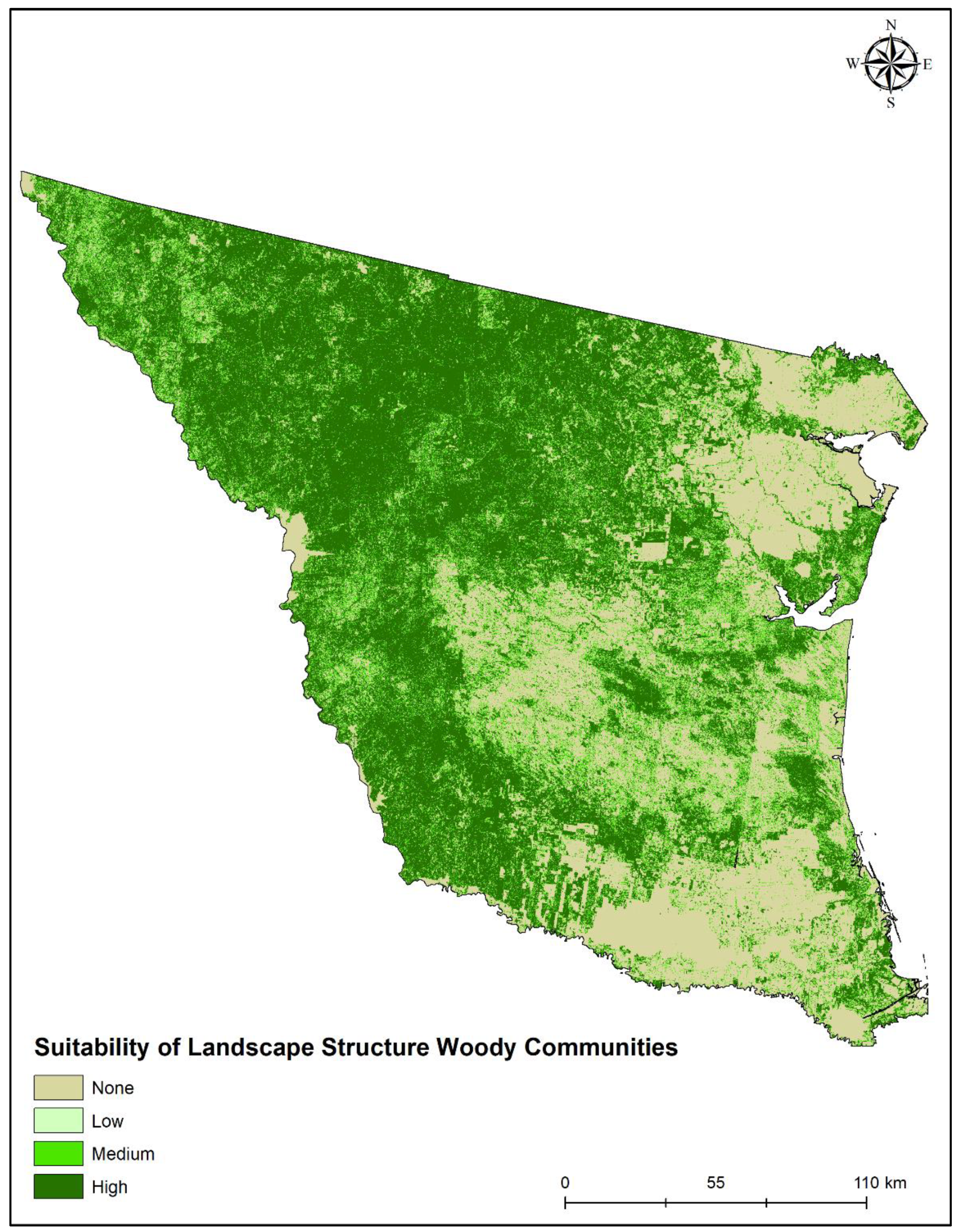
2.3. Landscape Suitability Analyses
2.4. Model Validation
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirzel, A.H.; le Lay, G.; Helfer, V.; Randin, C.; Guisan, A. Evaluating the ability of habitat suitability models to predict species presence. Ecol. Model. 2006, 199, 142–152. [Google Scholar] [CrossRef]
- Turner, M.G.; Gardner, R.G. Introduction to landscape ecology and scale. In Landscape Ecology in Theory and Practice; Turner, M.G., Gardner, R.G., Eds.; Springer: New York, NY, USA, 2015; pp. 1–32. [Google Scholar]
- Rondinini, C.; di Marco, M.; Chiozza, F.; Santulli, G.; Baisero, D.; Visconti, P.; Hoffmann, M.; Schipper, J.; Stuart, S.N.; Tognelli, M.F.; et al. Global habitat suitability models of terrestrial mammals. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Zimmermann, N.F. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Baasch, D.M.; Tyre, A.J.; Millspaugh, J.J.; Hygnstrom, S.E.; Vercauteren, K.C. An evaluation of three statistical methods used to model resource selection. Ecol. Model. 2010, 221, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.M.A.; Lesmeister, D.B.; David, R.J. Resource selection analysis. In Quantitative Analyses in Wildlife Science; Brennan., L.B., Tri, A.N., Marcot, B.G., Eds.; Johns Hopkins University Press in Affiliation with The Wildlife Society: Baltimore, MD, USA, 2021; pp. 199–215. [Google Scholar]
- Poor, E.E.; Scheick, B.K.; Mullinax, J.M. Multiscale consensus habitat modeling for landscape level conservation prioritization. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cianfrani, C.; Le Lay, G.; Hirzel, A.H.; Loy, A. Do habitat suitability models reliably predict the recovery areas of threatened species? J. Appl. Ecol. 2010, 47, 421–430. [Google Scholar] [CrossRef]
- Rondinini, C.; Stuart, S.; Boitani, L. Habitat suitability models and the shortfall in conservation planning for African vertebrates. Conserv. Biol. 2005, 19, 1488–1497. [Google Scholar] [CrossRef]
- Martínez-Meyer, E.; González-Bernal, A.; Velasco, J.A.; Swetnam, T.L.; González-Saucedo, Z.Y.; Servín, J.; López-González, C.A.; Oakleaf, J.K.; Liley, S.; Heffelfinger, J.R. Rangewide habitat suitability analysis for the Mexican wolf (Canis lupus baileyi) to identify recovery areas in its historical distribution. Div. Distrib. 2021, 27, 642–654. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Beckmann, J.P.; Beier, P.; Bird, B.; Bravo, J.C.; Fisher, K.; Grigione, M.M.; López-González, C.A.; Miller, J.R.; Mormorunni, C.; et al. The case for reintroduction, the jaguar (Panthera onca) in the United States as a model. Conserv. Sci. Pract. 2021. [Google Scholar] [CrossRef]
- Hirzel, A.; Guisan, A. Which is the optimal sampling strategy for habitat suitability modeling. Ecol. Model. 2002, 157, 331–341. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.F. Habitat Suitability and Distribution Models, with Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- McGarigal, K.; Cushman, S. The gradient concept of landscape structure. In Issues and Perspectives in Landscape Ecology; Wiens, J.A., Moss, M.R., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 112–119. [Google Scholar]
- Evans, J.S.; Cushman, S.A. Gradient modeling of conifer species using random forests. Landsc. Ecol. 2009, 24, 673–683. [Google Scholar] [CrossRef]
- Cushman, S.A.; Gutzweiler, K.; Evans, J.S.; McGarigal, K. The gradient paradigm, a conceptual and analytical framework for landscape ecology. In Spatial Complexity, Informatics, and Wildlife Conservation; Cushman, S.A., Huettmann, F., Eds.; Springer: Tokyo, Japan, 2010; pp. 83–108. [Google Scholar]
- Zemanova, M.A.; Perotto-Baldivieso, H.L.; Dickins, E.L.; Gill, A.B.; Leonard, J.P.; Wester, D.B. Impact of deforestation on habitat connectivity thresholds for large carnivores in tropical forests. Ecol. Proc. 2017, 6, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Perotto-Baldivieso, H.L. Essential concepts in landscape ecology for wildlife and natural resource managers. In Wildlife Management and Landscapes, Principles, and Applications; Porter, W.F., Parent, C.J., Stewart, R.A., Williams, D.M., Eds.; Johns Hopkins University Press in Affiliation with The Wildlife Society: Baltimore, MD, USA, 2021; pp. 53–67. [Google Scholar]
- Howery, L.D.; Bailey, D.W.; Laca, E.A. Impact of spatial memory on habitat use. Graz. Livest. Wildl. 1999, 70, 91–100. [Google Scholar]
- Cagnacci, F.; Boitani, L.; Powell, R.A.; Boyce, M.S. Animal ecology meets GPS-based radiotelemetry: A perfect storm of opportunities and challenges. Philos. Trans. R. Soc. B 2010, 365, 2157–2162. [Google Scholar] [CrossRef] [Green Version]
- Milleret, C.; Ordiz, A.; Sanz, A.; Uzal, A.; Carricondo-Sanchez, D.; Eriksen, A.; Sand, H.; Wabakken, P.; Wikenros, C.; Åkesson, M.; et al. Testing the influence of habitat experienced during the natal phase on habitat selection later in life in Scandinavian wolves. Sci. Rep. 2021, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Frazier, A.E.; Wang, L. Modeling landscape structure response across a gradient of land cover intensity. Landsc. Ecol. 2013, 28, 233–246. [Google Scholar] [CrossRef]
- Hoechstetter, S.; Walz, U.; Xuan, T.N. Effects of topography and surface roughness in analyses of landscape structure, a proposal to modify the existing set of landscape metrics. Landsc. Online 2008, 3, 1–14. [Google Scholar] [CrossRef]
- Frazier, A.E.; Kedron, P. Comparing forest fragmentation in eastern US forests using patch-mosaic and gradient surface models. Ecol. Inform. 2017, 41, 108–115. [Google Scholar] [CrossRef]
- Mandal, M.; Nilanjana, D.C. Spatial alteration of fragmented forest landscape for improving structural quality of habitat: A case study from Radhanagar Forest Range, Bankura District, West Bengal, India. Geol. Ecol. Landsc. 2020, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Moniem, H.E.M.A.; Holland, J.D. Habitat connectivity for pollinator beetles using surface metrics. Landsc. Ecol. 2013, 28, 1251–1267. [Google Scholar] [CrossRef]
- Hou, W.; Ulrich, W. An integrated approach for landscape contrast analysis with particular consideration of small habitats and ecotones. Nat. Conserv. 2016, 14, 25. [Google Scholar] [CrossRef] [Green Version]
- Mata, J.M. Landscape-Level Tanglehead Dynamics and the Effects on Northern Bobwhite Habitat. Master’s Thesis, Texas A&M University-Kingsville, Kingsville, TX, USA, 17 December 2017. [Google Scholar]
- Hunter, L. Carnivores of the World; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- International Union for the Conservation of Nature. The IUCN Red List of Threatened Species. 2020. Available online: http://www.iucnredlist.org (accessed on 14 January 2021).
- Lombardi, J.V.; Tewes, M.E.; Perotto-Baldivieso, H.L.; Mata, J.M.; Campbell, T.A. Spatial structure of woody cover affects habitat use patterns of ocelots in Texas. Mammal Res. 2020, 65, 555–563. [Google Scholar] [CrossRef]
- Blackburn, A.; Anderson, C.J.; Veals, A.M.; Tewes, M.E.; Wester, D.B.; Young, J.H.; DeYoung, R.W.; Perotto-Baldivieso, H.L. Landscape patterns of ocelot–vehicle collision sites. Landsc. Ecol. 2020, 17, 1–5. [Google Scholar] [CrossRef]
- Wang, B.; Rocha, D.G.; Abrahams, M.I.; Antunes, A.P.; Costa, H.C.; Gonçalves, A.L.; Spironello, W.R.; de Paula, M.J.; Peres, C.A.; Pezzuti, J.; et al. Habitat use of the ocelot (Leopardus pardalis) in Brazilian Amazon. Ecol. Evol. 2019, 9, 5049–5062. [Google Scholar] [CrossRef] [Green Version]
- Satter, C.B.; Augustine, B.C.; Harmsen, B.J.; Foster, R.J.; Sanchez, E.E.; Wultsch, C.; Davis, M.L.; Kelly, M.J. Long-term monitoring of ocelot densities in Belize. J. Wildl. Manag. 2019. [Google Scholar] [CrossRef]
- Janečka, J.E.; Davis, I.; Tewes, M.E.; Haines, A.M.; Caso, A.; Blankenship, T.L.; Honeycutt, R.L. Genetic differences in the response to landscape fragmentation by a habitat generalist, the bobcat and a habitat specialist, the ocelot. Conserv. Genet. 2016, 17, 1093–1108. [Google Scholar] [CrossRef]
- Haines, A.M.; Tewes, M.E.; Laack, L.L.; Horne, J.S.; Young, J.H. A habitat-based population viability analysis for ocelots (Leopardus pardalis) in the United States. Biol. Conserv. 2006, 136, 326–327. [Google Scholar] [CrossRef]
- Tewes, M.E. Conservation Status of the Endangered Ocelot in the United States—A 35-Year Perspective; Texas A&M University-Kingsville: Kingsville, TX, USA, 2019. [Google Scholar]
- Leonard, J.P.; Tewes, M.E.; Lombardi, J.V.; Wester, D.B.; Campbell, T.A. Effects of sun angle, lunar illumination, and diurnal temperature on temporal movement rates of sympatric ocelots and bobcats in South Texas. PLoS ONE 2020, 15, e0231732. [Google Scholar] [CrossRef]
- Schmidt, G.M.; Lewison, R.L.; Swarts, H.M. Identifying landscape predictors of ocelot road mortality. Landsc. Ecol. 2020, 35, 1651–1666. [Google Scholar] [CrossRef]
- Lehnen, S.E.; Sternberg, M.A.; Swarts, H.M.; Sesnie, S.E. Evaluating the population connectivity and targeting conservation action for an endangered cat. Ecosphere 2021, 12, e03367. [Google Scholar] [CrossRef]
- Jackson, V.L.; Laack, L.L.; Zimmerman, E.G. Landscape metrics associated with habitat use by ocelots in South Texas. J. Wildl. Manag. 2005, 69, 733–738. [Google Scholar] [CrossRef]
- Leslie, D.M., Jr. An International Borderland of Concern: Conservation of Biodiversity in the Lower Rio Grande Valley; US Geological Survey: Stillwater, OK, USA, 2016. [Google Scholar]
- Tremblay, T.A.; White, W.A.; Raney, J.A. Native woodland loss during the mid-1900s in Cameron County, Texas. Southwest Nat. 2005, 50, 479–519. [Google Scholar] [CrossRef]
- Lombardi, J.V.; Perotto–Baldivieso, H.L.; Tewes, M.E. Land cover trends in South Texas (1987–2050): Potential implications for wild felids. Remote Sens. 2020, 12, 659. [Google Scholar] [CrossRef] [Green Version]
- García, R.S.; Botero-Cañola, S.; Sánchez-Giraldo, C.; Solari, S. Habitat use and activity patterns of Leopardus pardalis (Felidae) in the Northern Andes, Antioquia, Columbia. Biodiversity 2019, 20, 5–19. [Google Scholar] [CrossRef]
- Tewes, M.E.; Everett, D.D. Status and distribution of the endangered ocelot and jaguarundi in Texas. In Cats of the World: Biology, Conservation, and Management; Miller, S.D., Everett, D.D., Eds.; National Wildlife Federation: Washington, DC, USA, 1986; pp. 147–158. [Google Scholar]
- Connolly, A.R. Defining Habitat for the Recovery of Ocelots (Leopardus pardalis) in the United States. Master’s Thesis, Texas State University, San Marcos, TX, USA, 2009. [Google Scholar]
- Griffith, G.; Bryce, S.; Omerik, J.; Rogers, A. Eco-Regions of Texas; Texas Commission on Environmental Quality: Austin, TX, USA, 2007. [Google Scholar]
- Shindle, D.B.; Tewes, M.E. Immobilization of wild ocelots with tiletamine and zolazepam in southern Texas. J. Wildl. Dis. 2000, 36, 546–550. [Google Scholar] [CrossRef] [Green Version]
- Perotto-Baldivieso, H.L.; Cooper, S.M.; Cibils, A.F.; Figueroa-Pagán, M.; Udaeta, K.; Black-Rubio, C.M. Detecting autocorrelation problems from GPS collar data in livestock studies. Appl. Anim. Behav. Sci. 2012, 136, 117–125. [Google Scholar] [CrossRef]
- US Geological Survey Global Visualization Viewer. Available online: https://glovis.usgs.gov/ (accessed on 20 June 2019).
- Guthery, F.S. Slack in the configuration of habitat patches for northern bobwhites. J. Wildl. Manag. 1999, 63, 245–250. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Neel, M.C.; Ene, E. FRAGSTATS v4.2: Spatial Pattern Analysis Program for Categorical and Continuous Maps. University of Massachusetts. 2015. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 15 October 2020).
- Paolino, R.M.; Royle, J.A.; Versiani, N.F.; Rodrigues, T.F.; Pasqualotto, N.; Krepschi, V.G.; Chiarello, A.G. Importance of riparian forest corridors for the ocelot in agricultural landscapes. J. Mammal. 2018, 99, 874–884. [Google Scholar] [CrossRef]
- Perotto-Baldiviezo, H.L.; Thurow, T.L.; Smith, C.T.; Fisher, R.F.; Wu, X.B. GIS-based spatial analysis and modeling for landslide hazard assessment in steeplands, Southern Honduras. Agric. Ecosyst. Environ. 2004, 103, 165–176. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Recovery Plan for the Ocelot (Leopardus pardalis), First Revision; Southwest Region, U.S. Fish and Wildlife Service: Albuquerque, NM, USA, 2016. [Google Scholar]
- Laack, L.L.; Tewes, M.E.; Haines, A.M.; Rappole, J.H. Reproductive life history of ocelots Leopardus pardalis in southern Texas. Acta Ther. 2005, 50, 505–514. [Google Scholar] [CrossRef]
- Booth-Binczik, S.D.; Bradley, R.D.; Thompson, C.W.; Bender, L.C.; Huntley, J.W.; Harvey, J.A.; Laack, L.L.; Mays, J.L. Food habits of ocelots and potential for competition with bobcats in southern Texas. Southwest Nat. 2013, 1, 403–410. [Google Scholar] [CrossRef]
- Santos, F.; Carbone, C.; Wearn, O.R.; Rowcliffe, J.M.; Espinosa, S.; Lima, M.G.; Ahumada, J.A.; Gonçalves, A.L.; Trevelin, L.C.; Alvarez-Loayza, P.; et al. Prey availability and temporal partitioning modulate felid coexistence in Neotropical forests. PLoS ONE 2019, 14, e0213671. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Reis, M.B.; Nichols, J.D.; Chiarello, A.G.; Ribeiro, M.C.; Setz, E.Z. Landscape use and co-occurrence patterns of Neotropical spotted cats. PLoS ONE 2017, 12, e0168441. [Google Scholar] [CrossRef]
- Cruz, P.; Iezzi, M.E.; de Angelo, C.; Varela, D.; Di Bitetti, M.S.; Paviolo, A. Effects of human impacts on habitat use, activity patterns and ecological relationships among medium and small felids of the Atlantic Forest. PLoS ONE 2018, 13, e0200806. [Google Scholar] [CrossRef]
- Massara, R.L.; de Oliveira-Paschoal, A.M.; Bailey, L.L.; Doherty, P.F., Jr.; de Frias, B.M.; Chiarello, A.G. Effect of humans and pumas on the temporal activity of ocelots in protected areas of Atlantic Forest. Mamm. Biol. 2018, 92, 86–93. [Google Scholar] [CrossRef]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Zeller, K.A.; Wattles, D.W.; DeStefano, S. Evaluating methods for identifying large mammal road crossing locations: Black bears as a case study. Landsc. Ecol. 2020, 35, 1799–1808. [Google Scholar] [CrossRef]
- Cerqueira, R.C.; Leonard, P.B.; da Silva, L.G.; Bager, A.; Clevenger, A.P.; Jaeger, J.A.; Grilo, C. Potential movement corridors and high road-kill likelihood do not spatially coincide for felids in Brazil: Implications for road mitigation. Environ. Manag. 2021, 67, 412–423. [Google Scholar] [CrossRef]
- Young, D.; Perotto-Baldivieso, H.L.; Brewer, T.; Homer, R.; Santos, S.A. Monitoring British upland grazing ecosystems with the use of landscape structure as an indicator for state-and-transition models. Rangel. Ecol. Manag. 2014, 67, 380–388. [Google Scholar] [CrossRef]
- Perotto-Baldivieso, H.L.; Tapaneeyakul, S.; Pearson, Z.J. Using geospatial technologies in wildlife studies. In Wildlife Techniques Manual, 8th ed.; Research: Applications of Spatial Technologies in Wildlife Research; Silvy, N.J., Ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2020; Volume I, pp. 495–510. [Google Scholar]
- Horne, J.S.; Haines, A.M.; Tewes, M.E.; Laack, L.L. Habitat partitioning by sympatric ocelots and bobcats: Implications for recovery of ocelots in southern Texas. Southwest Nat. 2009, 54, 119–126. [Google Scholar] [CrossRef]
- Blackburn, A.; Heffelfinger, L.J.; Veals, A.M.; Tewes, M.E.; Young, J.H., Jr. Cats, cars, and crossings: The consequences of road networks for the conservation of an endangered felid. Glob. Ecol. Conserv. 2021, 27, e01582. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Landscape Metric | Prediction | Citation |
|---|---|---|
| Landscape Shape Index | Ocelots use a range of values with lower shape values on the landscape | Jackson et al., 2005, Lombardi et al., 2020 |
| Percent Landscape (%) | Ocelots use a range of higher percentage of woody cover on the landscape | Tewes and Everett 1986, Jackson et al., 2005, Haines et al., 2006, Connally 2009, Massara et al., 2018, Paolino et al., 2018, Wang et al., 2019, Lombardi et al., 2020, Blackburn et al., 2020 |
| Patch Density (patches/100 ha) | Ocelots use a range of lower patch densities, indicative of large patches on the landscape | Wang et al., 2019, Blackburn et al., 2020 |
| Largest Patch Index (%) | Ocelots use a range of larger woody patch indices on the landscape | Tewes and Everett 1986, Jackson et al., 2005, Haines et al., 2006, Connally 2009, Massara et al., 2018, Paolino et al., 2018, Wang et al., 2019, Lombardi et al., 2020, Blackburn et al., 2020 |
| Edge Density (m/ha) | Ocelots use a range of lower edge densities, indicative of a forest -interior species on the landscape | Garcia-R et al., 2019 |
| Mean Patch Area (ha) | Ocelots use the largest mean patch areas on the landscape | Jackson et al., 2005, Lombardi et al., 2020, Wang et al., 2019 |
| Aggregation Index | Ocelots use a range of larger aggregation indices on the landscape | Blackburn et al., 2020 |
| Landscape Metric | Criterion | Score |
|---|---|---|
| Landscape Shape Index | 0–1.07 | 0 |
| 1.07–2.25 | 1 | |
| 2.25–3.37 | 0 | |
| Percent Landscape (%) | 0–41.3 | 0 |
| 41.3–100 | 1 | |
| Patch Density (patches/100 ha) | 0–22.6 | 0 |
| 22.7–72.4 | 1 | |
| 72.5–306.5 | 0 | |
| Largest Patch Index (%) | 0–34.3 | 0 |
| 34.4–100 | 1 | |
| Edge Density (m/ha) | 0–191.5 | 1 |
| 191.6–551.7 | 0 | |
| Mean Patch Area (ha) | 0–0.86 | 0 |
| 0.87–4.4 | 1 | |
| Aggregation Index | 0–54 | 0 |
| 55–98 | 1 |
| Year | High | Medium | Low | None | χ2 |
|---|---|---|---|---|---|
| Expected | 84.8 | 6.92 | 4.1 | 4.18 | |
| Observed 2013 | 90.9 | 2.2 | 4.5 | 2.4 | 4.42 |
| Observed 2014 | 65 | 15.8 | 13.8 | 5.4 | 39.6 * |
| Observed 2015 | 87.9 | 8.1 | 1.83 | 6.5 | 2.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombardi, J.V.; Perotto-Baldivieso, H.L.; Sergeyev, M.; Veals, A.M.; Schofield, L.; Young, J.H.; Tewes, M.E. Landscape Structure of Woody Cover Patches for Endangered Ocelots in Southern Texas. Remote Sens. 2021, 13, 4001. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13194001
Lombardi JV, Perotto-Baldivieso HL, Sergeyev M, Veals AM, Schofield L, Young JH, Tewes ME. Landscape Structure of Woody Cover Patches for Endangered Ocelots in Southern Texas. Remote Sensing. 2021; 13(19):4001. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13194001
Chicago/Turabian StyleLombardi, Jason V., Humberto L. Perotto-Baldivieso, Maksim Sergeyev, Amanda M. Veals, Landon Schofield, John H. Young, and Michael E. Tewes. 2021. "Landscape Structure of Woody Cover Patches for Endangered Ocelots in Southern Texas" Remote Sensing 13, no. 19: 4001. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13194001