
Crop Water Stress Index as a Proxy of Phenotyping Maize Performance under Combined Water and Salt Stress


Abstract
:1. Introduction
2. Materials and Methods
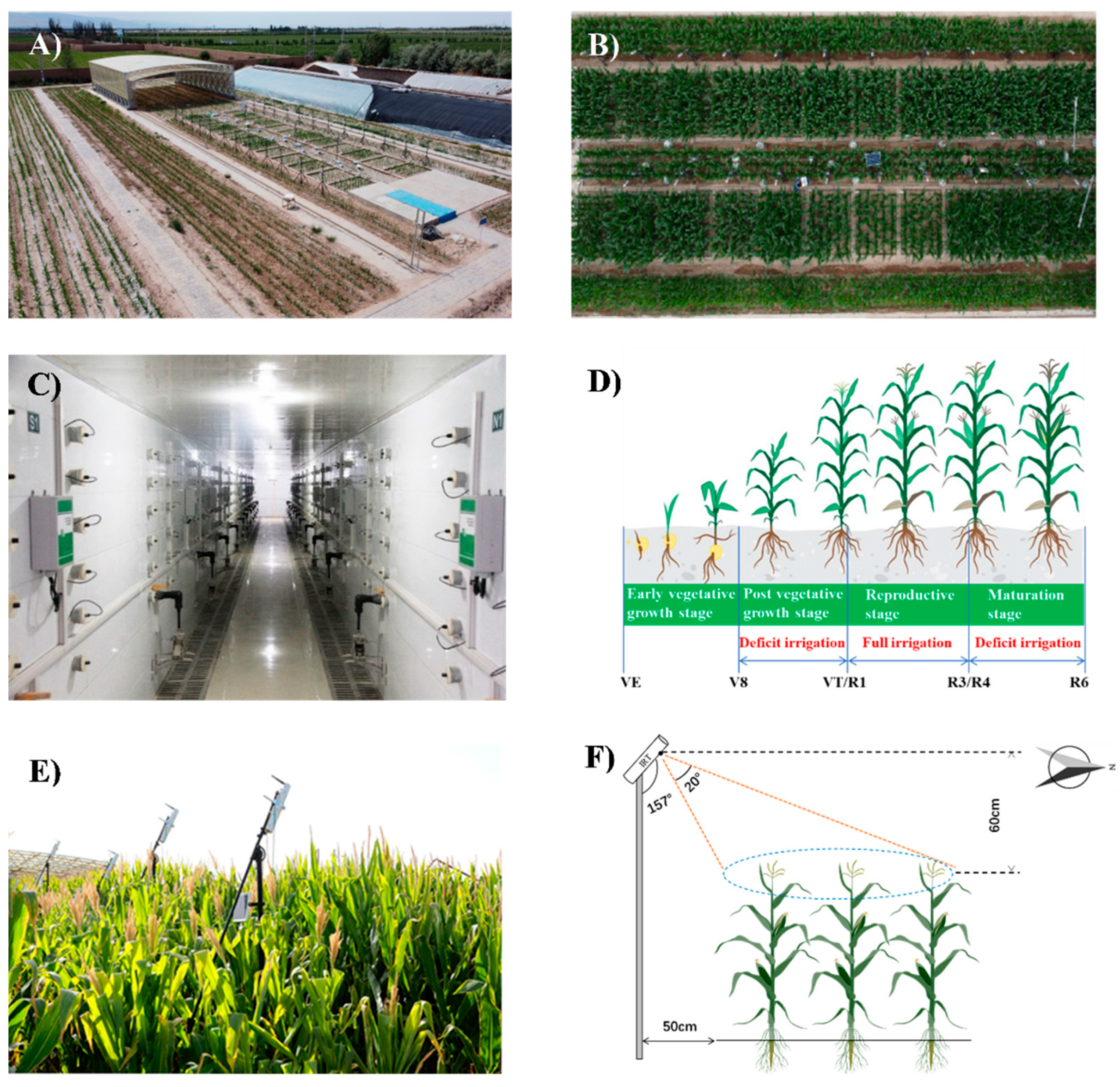
2.1. Experimental Area and Design
2.2. Canopy Temperature and CWSI Calculation
2.3. Gas Exchange and Water Potential Measurements
2.4. Measurement of Leaf Area Index, Effective Leaf Width, Biomass, Yield, and ET
2.5. Data Analysis
3. Results
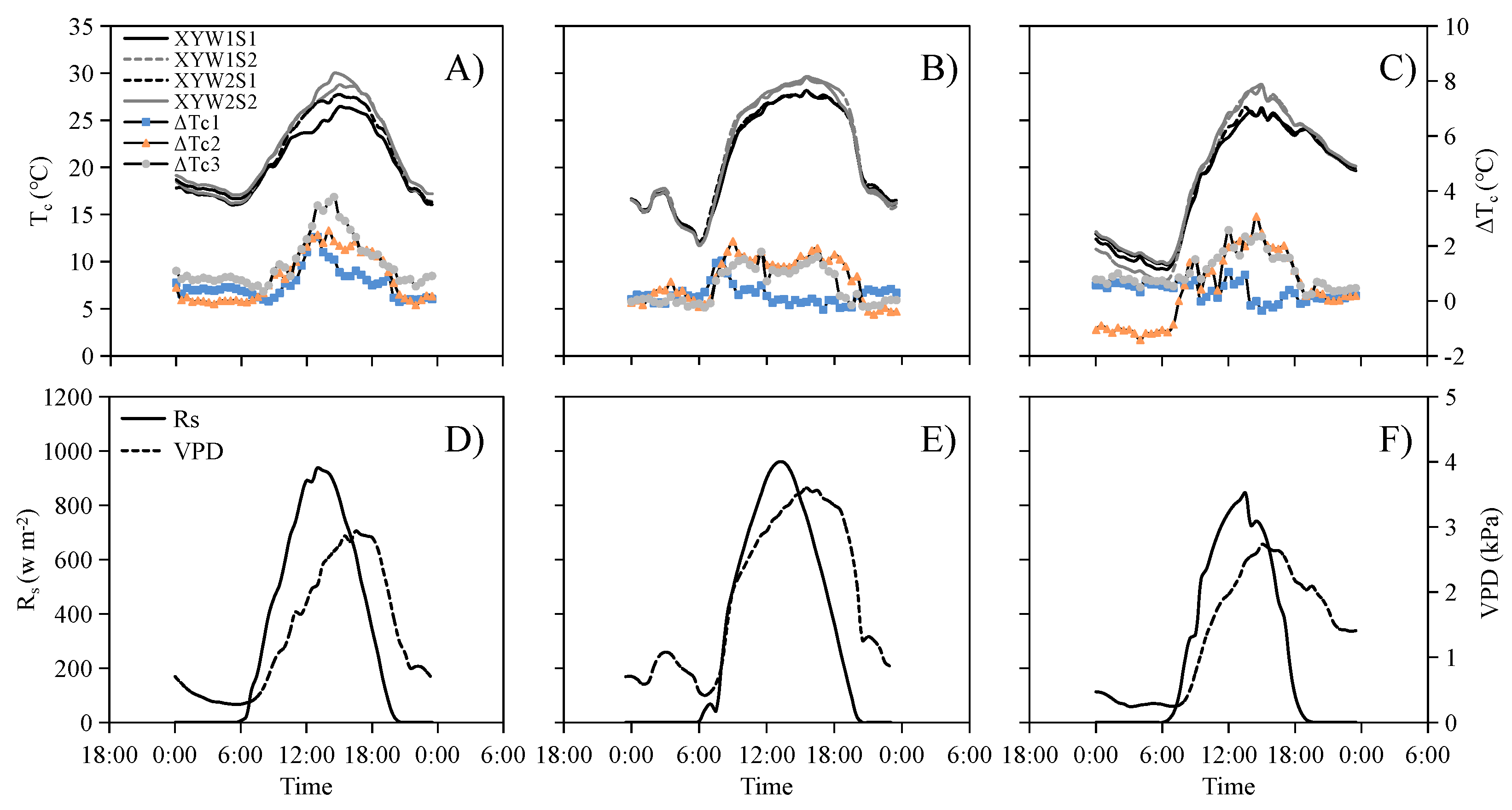
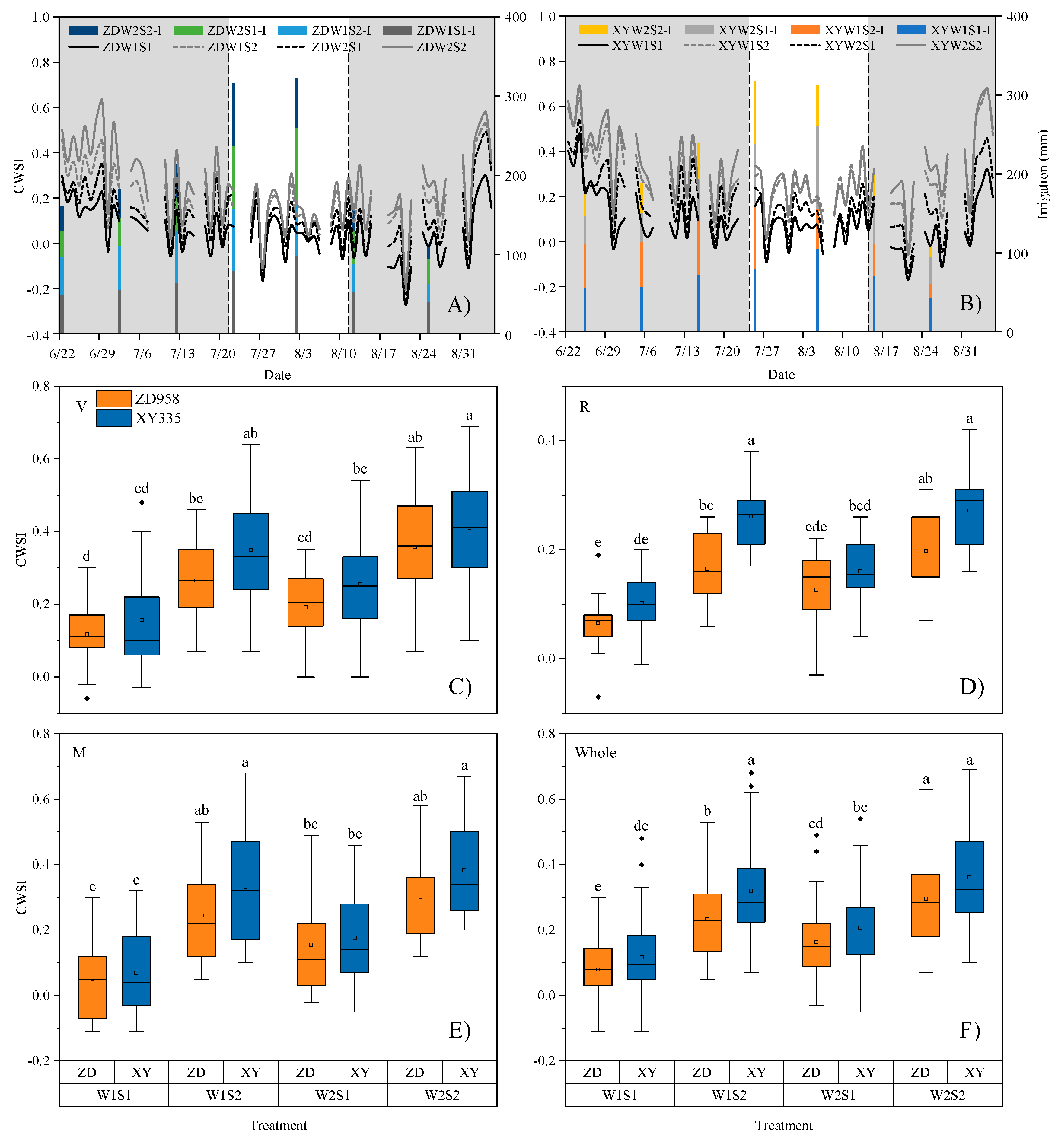
3.1. Tc Can Characterize the Degree of Water and Salt Stress on Maize
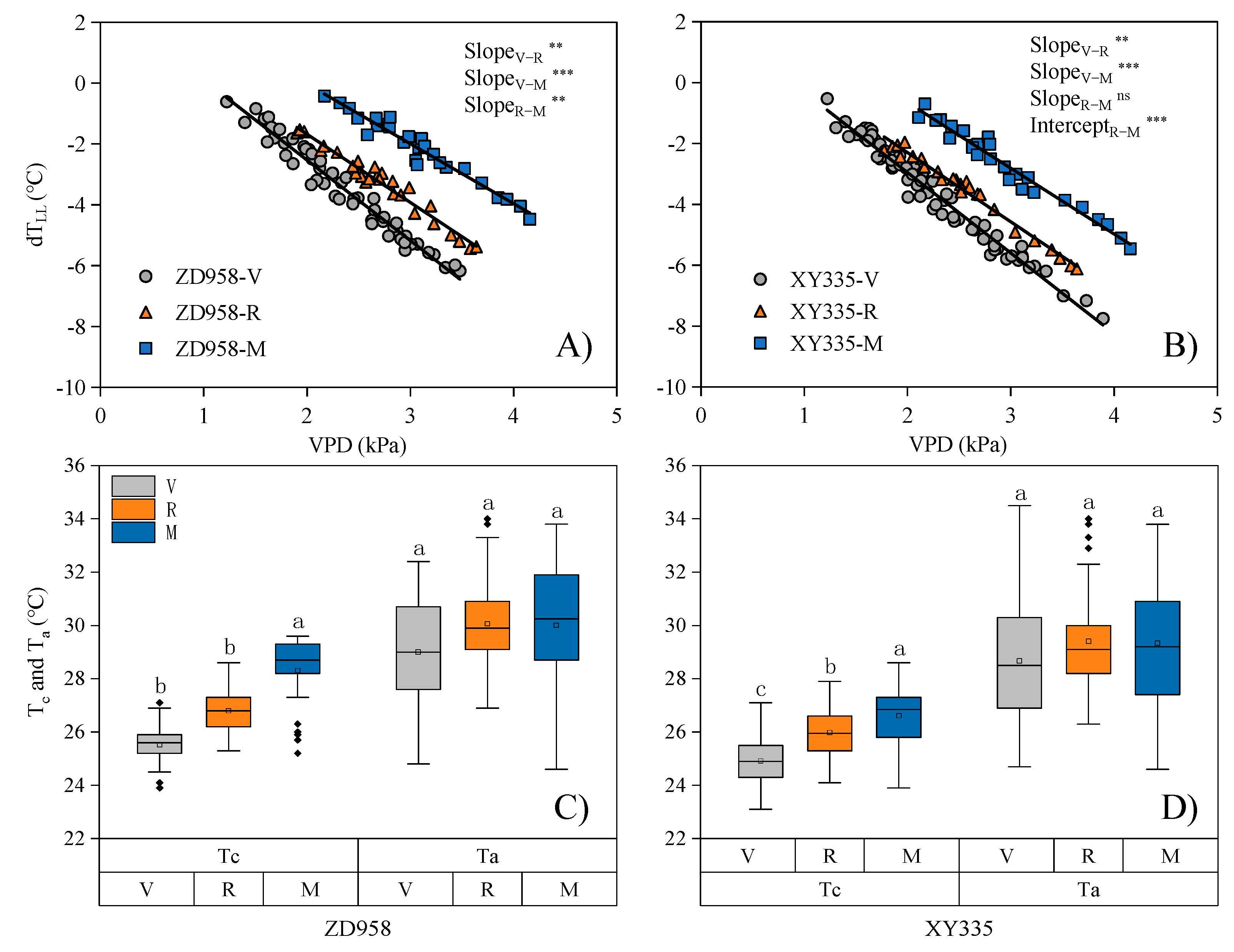
3.2. Growth Stage-Specific NWSBs Are Necessary for Establishing the CWSI Model
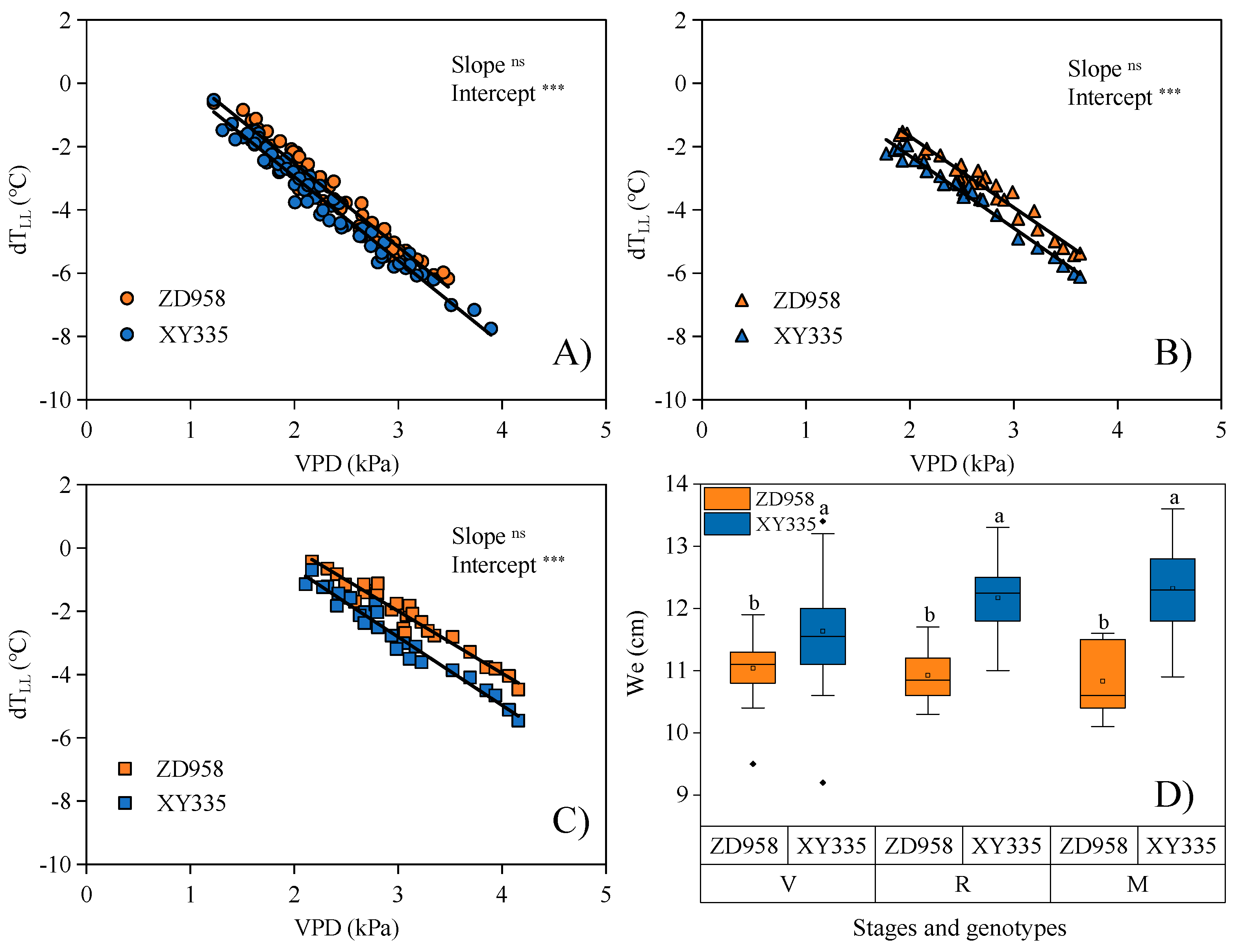
3.3. Significant Differences in NWSB Are Attributed to the Differences in Leaf Morphology between Maize Genotypes
3.4. CWSI Characterizes the Seasonal Dynamics of Maize under Water and Salt Stress
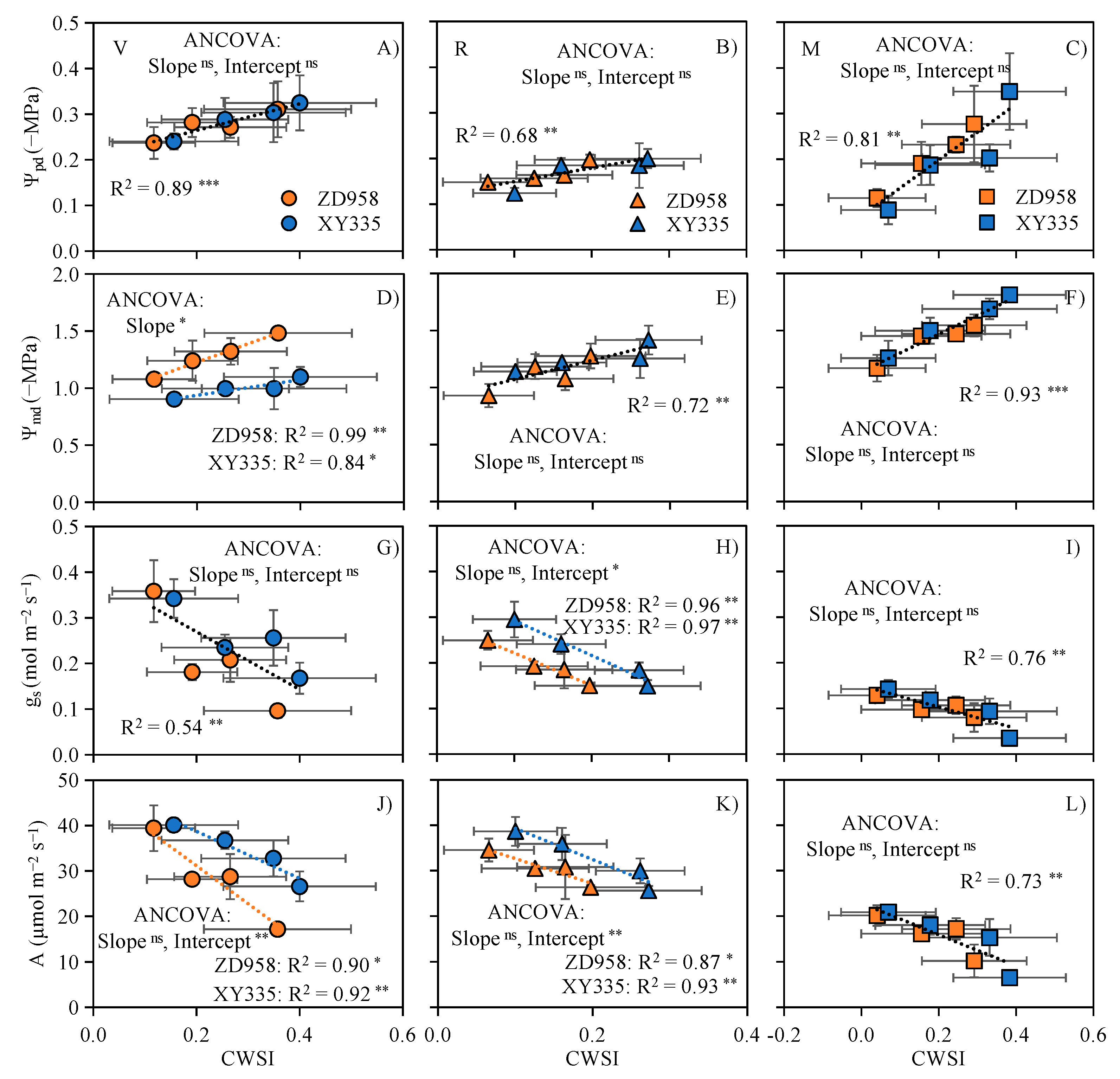
3.5. CWSI Can Diagnose the Physiological Variations of Maize under Water and Salt Stress
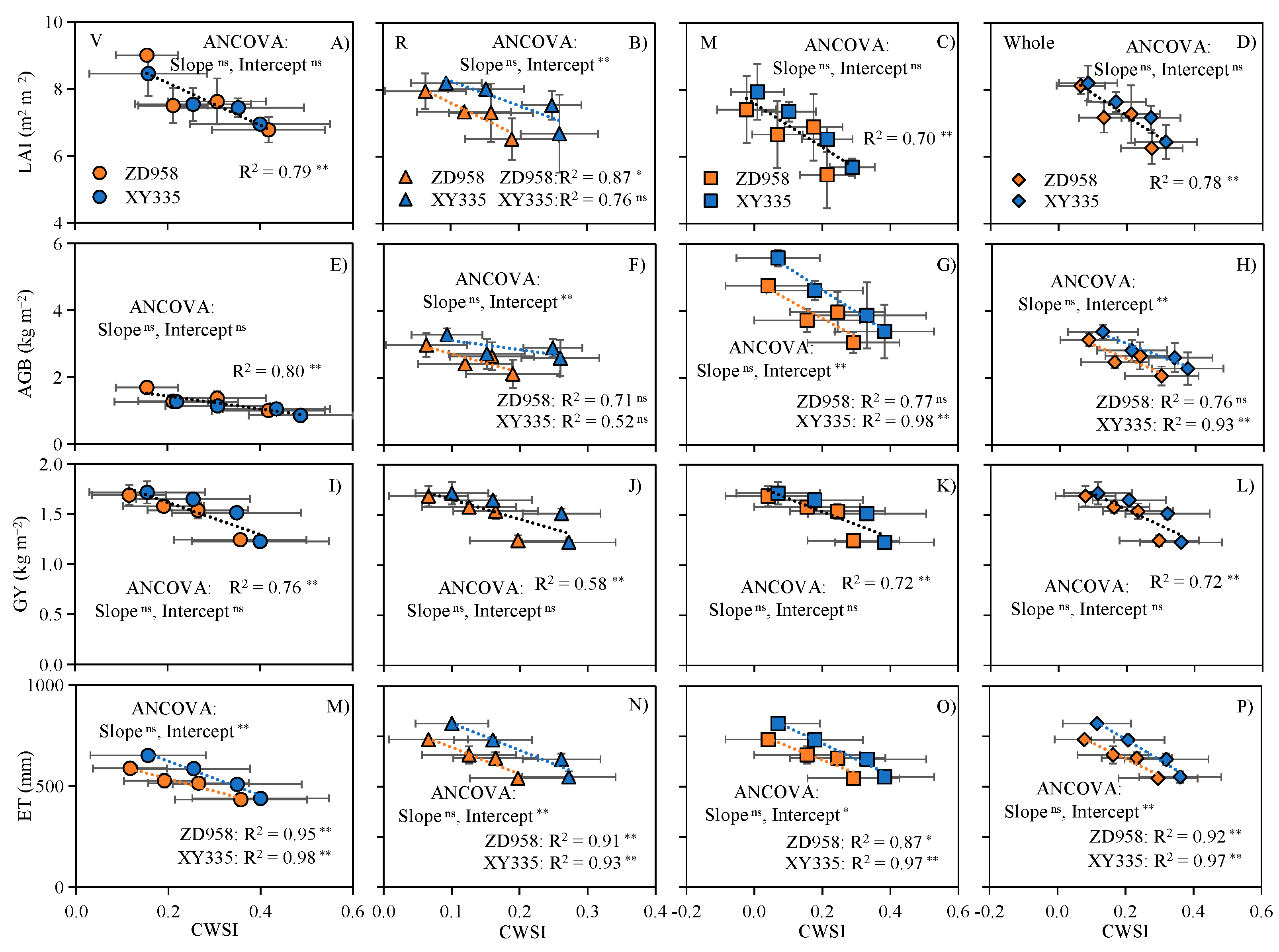
3.6. CWSI Can Predict Maize Growth, Yield, and Water Use under Water and Salt Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alghory, A.; Yazar, A. Evaluation of crop water stress index and leaf water potential for deficit irrigation management of sprinkler-irrigated wheat. Irrig. Sci. 2019, 37, 61–77. [Google Scholar] [CrossRef]
- Kang, S.; Hao, X.; Du, T.; Tong, L.; Su, X.; Lu, H.; Li, X.; Huo, Z.; Li, S.; Ding, R. Improving agricultural water productivity to ensure food security in China under changing environment: From research to practice. Agric. Water Manag. 2017, 179, 5–17. [Google Scholar] [CrossRef]
- Cakir, R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crop. Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Jiang, J.; Feng, S.; Ma, J.; Huo, Z.; Zhang, C. Irrigation management for spring maize grown on saline soil based on SWAP model. Field Crop. Res. 2016, 196, 85–97. [Google Scholar] [CrossRef]
- Hu, Y.; Burucs, Z.; von Tucher, S.; Schmidhalter, U. Short-term effects of drought and salinity on mineral nutrient distribution along growing leaves of maize seedlings. Environ. Exp. Bot. 2007, 60, 268–275. [Google Scholar] [CrossRef]
- Yuan, C.; Feng, S.; Huo, Z.; Ji, Q. Effects of deficit irrigation with saline water on soil water-salt distribution and water use efficiency of maize for seed production in arid Northwest China. Agric. Water Manag. 2019, 212, 424–432. [Google Scholar] [CrossRef]
- Jones, H.G. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology; Cambridge University Press: New York, NY, USA, 2014. [Google Scholar]
- Ben-Gal, A.; Agam, N.; Alchanatis, V.; Cohen, Y.; Yermiyahu, U.; Zipori, I.; Presnov, E.; Sprintsin, M.; Dag, A. Evaluating water stress in irrigated olives: Correlation of soil water status, tree water status, and thermal imagery. Irrig. Sci. 2009, 27, 367–376. [Google Scholar] [CrossRef]
- Gollan, T.; Turner, N.C.; Schulze, E.D. The responses of stomata and leaf gas exchange to vapour pressure deficits and soil water content III. In the sclerophyllous woody species Nerium oleander. Oecologia 1985, 65, 356–362. [Google Scholar] [CrossRef]
- Ding, R.; Kang, S.; Zhang, Y.; Hao, X.; Tong, L.; Li, S. A dynamic surface conductance to predict crop water use from partial to full canopy cover. Agric. Water Manag. 2015, 150, 1–8. [Google Scholar] [CrossRef]
- Jones, H.G. Plant water relations and implications for irrigation scheduling. Acta Horticul. 1990, 278, 67–76. [Google Scholar] [CrossRef]
- Ballester, C.; Jimenez-Bello, M.A.; Castel, J.R.; Intrigliolo, D.S. Usefulness of thermography for plant water stress detection in citrus and persimmon trees. Agric. Forest Meteorol. 2013, 168, 120–129. [Google Scholar] [CrossRef]
- Zhu, J.; Bo, X.; Liu, L. Improved protocol for the in situ measurement of water potential of plants with a thermocouple psychrometer. Chin. Bull. Bot. 2013, 48, 531–539, (in Chinese with English abstract). [Google Scholar]
- Agam, N.; Cohen, Y.; Berni, J.A.J.; Alchanatis, V.; Kool, D.; Dag, A.; Yermiyahu, U.; Ben-Gal, A. An insight to the performance of crop water stress index for olive trees. Agric. Water Manag. 2013, 118, 79–86. [Google Scholar] [CrossRef]
- Kullberg, E.G.; Dejonge, K.C.; Chavez, J.L. Evaluation of thermal remote sensing indices to estimate crop evapotranspiration coefficients. Agric. Water Manag. 2017, 179, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, T.C. Plant responses to water stress. Annu. Rev. Plant. Biol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Argyrokastritis, I.G.; Papastylianou, P.T.; Alexandris, S. Leaf water potential and crop water stress index variation for full and deficit irrigated cotton in Mediterranean conditions. Agric. Agric. Sci. Procedia 2015, 4, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Fitriyah, A.; Fatikhunnada, A.; Okura, F.; Nugroho, B.D.A.; Kato, T. Analysis of the drought mitigated mechanism in terraced paddy fields using CWSI and TVDI indices and hydrological monitoring. Sustainability 2019, 11, 6897. [Google Scholar] [CrossRef] [Green Version]
- O’Shaughnessy, S.A.; Evett, S.R.; Colaizzi, P.D.; Howell, T.A. A crop water stress index and time threshold for automatic irrigation scheduling of grain sorghum. Agric. Water Manag. 2012, 107, 122–132. [Google Scholar] [CrossRef] [Green Version]
- Veysi, S.; Naseri, A.A.; Hamzeh, S.; Bartholomeus, H. A satellite based crop water stress index for irrigation scheduling in sugarcane fields. Agric. Water Manag. 2017, 189, 70–86. [Google Scholar] [CrossRef]
- Yuan, G.; Luo, Y.; Sun, X.; Tang, D. Evaluation of a crop water stress index for detecting water stress in winter wheat in the North China Plain. Agric. Water Manag. 2004, 64, 29–40. [Google Scholar] [CrossRef]
- Bellvert, J.; Zarco-Tejada, P.J.; Marsal, J.; Girona, J.; Gonzalez-Dugo, V.; Fereres, E. Vineyard irrigation scheduling based on airborne thermal imagery and water potential thresholds. Aust. J. Grape Wine Res. 2016, 22, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Padilla-Diaz, C.M.; Martinez-Guanter, J.; Fernandez, J.E.; Perez-Ruiz, M. Assessing a crop water stress index derived from aerial thermal imaging and infrared thermometry in super-high density olive orchards. Agric. Water Manag. 2017, 187, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Applicability and limitations of using the crop water stress index as an indicator of water deficits in citrus orchards. Agric. For. Meteorol. 2014, 198, 94–104. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Lopez-Lopez, M.; Espadafor, M.; Orgaz, F.; Testi, L.; Zarco-Tejada, P.; Lorite, I.J.; Fereres, E. Transpiration from canopy temperature: Implications for the assessment of crop yield in almond orchards. Eur. J. Agron. 2019, 105, 78–85. [Google Scholar] [CrossRef]
- Jackson, S.H. Relationships between normalized leaf water potential and crop water stress index values for acala cotton. Agric. Water Manag. 1991, 20, 109–118. [Google Scholar] [CrossRef]
- Idso, S.B. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Taghvaeian, S.; Comas, L.; Dejonge, K.C.; Trout, T.J. Conventional and simplified canopy temperature indices predict water stress in sunflower. Agric. Water Manag. 2014, 144, 69–80. [Google Scholar] [CrossRef]
- Irmak, S.; Haman, D.Z.; Bastug, R. Determination of crop water stress index for irrigation timing and yield estimation of corn. Agron. J. 2000, 92, 1221–1227. [Google Scholar] [CrossRef]
- Gardner, B.R.; Nielsen, D.C.; Shock, C.C. Infrared thermometry and the crop water stress index. I. History, theory, and baselines. J. Prod. Agric. 1992, 5, 462–466. [Google Scholar] [CrossRef]
- Aladenola, O.; Madramootoo, C. Response of greenhouse-grown bell pepper (Capsicum annuum L.) to variable irrigation. Can. J. Plant Sci. 2014, 94, 303–310. [Google Scholar] [CrossRef]
- Bellvert, J.; Marsal, J.; Girona, J.; Gonzalez-Dugo, V.; Fereres, E.; Ustin, S.L.; Zarco-Tejada, P.J. Airborne thermal imagery to detect the seasonal evolution of crop water status in peach, nectarine and saturn peach orchards. Remote Sens. 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Rud, R.; Cohen, Y.; Alchanatis, V.; Levi, A.; Brikman, R.; Shenderey, C.; Heuer, B.; Markovitch, T.; Dar, Z.; Rosen, C.; et al. Crop water stress index derived from multi-year ground and aerial thermal images as an indicator of potato water status. Precis. Agric. 2014, 15, 273–289. [Google Scholar] [CrossRef]
- Xu, J.; Lv, Y.; Liu, X.; Dalson, T.; Yang, S.; Wu, J. Diagnosing crop water stress of rice using infra-red thermal imager under water deficit condition. Int. J. Agric. Biol. 2016, 18, 565–572. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, G.; Meng, Y.; Chen, B.; Wang, Y.; Zhou, Z. Changes of Related Physiological Characteristics of Cotton Under Salinity Condition and the Construction of the Cotton Water Stress Index. China Agric. Sci. 2013, 46, 3768–3775, (in Chinese with English abstract). [Google Scholar]
- Fan, Y.; Ding, R.; Kang, S.; Hao, X.; Du, T.; Tong, L.; Li, S. Plastic mulch decreases available energy and evapotranspiration and improves yield and water use efficiency in an irrigated maize cropland. Agric. Water Manag. 2017, 179, 122–131. [Google Scholar] [CrossRef]
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development; Iowa State University: Ames, IA, USA, 2011. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements; FAO: Rome, Italy, 1998; FAO Irrigation and Drainage Paper No. 56. [Google Scholar]
- Taghvaeian, S.; Chavez, J.L.; Hansen, N.C. Infrared thermometry to estimate crop water stress index and water use of irrigated maize in northeastern Colorado. Remote Sens. 2012, 4, 3619–3637. [Google Scholar] [CrossRef] [Green Version]
- Dejonge, K.C.; Taghvaeian, S.; Trout, T.J.; Comas, L.H. Comparison of canopy temperature-based water stress indices for maize. Agric. Water Manag. 2015, 156, 51–62. [Google Scholar] [CrossRef]
- Steele, D.D.; Stegman, E.C.; Gregor, B.L. Field comparison of irrigation scheduling methods for corn. Trans. ASAE 1994, 37, 1197–1203. [Google Scholar] [CrossRef]
- Leigh, A.; Sevanto, S.; Close, J.D.; Nicotra, A.B. The influence of leaf size and shape on leaf thermal dynamics: Does theory hold up under natural conditions? Plant Cell Environ. 2017, 40, 237–248. [Google Scholar] [CrossRef]
- Mcdonald, P.G.; Fonseca, C.R.; Overton, J.M.; Westoby, M. Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? Funct. Ecol. 2003, 17, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Taghvaeian, S.; Chavez, J.L.; Bausch, W.C.; Dejonge, K.C.; Trout, T.J. Minimizing instrumentation requirement for estimating crop water stress index and transpiration of maize. Irrig. Sci. 2014, 32, 53–65. [Google Scholar] [CrossRef]
- Yazar, A.; Howell, T.A.; Dusek, D.A.; Copeland, K.S. Evaluation of crop water stress index for LEPA irrigated corn. Irrig. Sci. 1999, 18, 171–180. [Google Scholar] [CrossRef]
- Han, M.; Zhang, H.; Dejonge, K.C.; Comas, L.H.; Gleason, S. Comparison of three crop water stress index models with sap flow measurements in maize. Agric. Water Manag. 2018, 203, 366–375. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X.; Wang, W.; Ran, H.; Song, T.; Guo, Y. Evaluation of the crop water stress index as an indicator for the diagnosis of grapevine water deficiency in greenhouses. Horticulturae 2020, 6, 864. [Google Scholar] [CrossRef]
- Zhang, L.; Niu, Y.; Han, W.; Liu, Z. Establishing method of crop water stress index empirical model of field maize. Trans. Chin. Soc. Agric. Mach. 2018, 49, 233–239, (in Chinese with English abstract). [Google Scholar]
- Cui, X.; Xu, L.; Yuan, G.; Wang, W.; Luo, Y. Crop water stress index model for monitoring summer maize water stress based on canopy surface temperature. Trans. Chin. Soc. Agric. Eng. 2005, 21, 22–24, (in Chinese with English abstract). [Google Scholar]
- Nielsen, D.C. Non water-stressed baselines for sunflowers. Agric. Water Manag. 1994, 26, 265–276. [Google Scholar] [CrossRef]
- Gontia, N.K.; Tiwari, K.N. Development of crop water stress index of wheat crop for scheduling irrigation using infrared thermometry. Agric. Water Manag. 2008, 95, 1144–1152. [Google Scholar] [CrossRef]
- Fritschen, L.J.; Bavel, C.H.M.V. Energy balance as affected by height and maturity of sudangrass. Agron. J. 1964, 56, 201–204. [Google Scholar] [CrossRef]
- Idso, S.B.; Reginato, R.J.; Clawson, K.L.; Anderson, M.G. On the stability of non-water-stressed baselines. Agric. For. Meteorol. 1984, 32, 177–182. [Google Scholar] [CrossRef]
- Smith, W.K. Temperatures of desert plants: another perspective on the adaptability of leaf size. Science 1978, 201, 614–616. [Google Scholar] [CrossRef] [PubMed]
- Meron, M.; Tsipris, J.; Orlov, V.; Alchanatis, V.; Cohen, Y. Crop water stress mapping for site-specific irrigation by thermal imagery and artificial reference surfaces. Precis. Agric. 2010, 11, 148–162. [Google Scholar] [CrossRef]
- O’Shaughnessy, S.A.; Evett, S.R.; Colaizzi, P.D.; Howell, T.A. Using radiation thermography and thermometry to evaluate crop water stress in soybean and cotton. Agric. Water Manag. 2011, 98, 1523–1535. [Google Scholar] [CrossRef]
- Jones, H.G. Stomatal control of photosynthesis and transpiration. J. Exp. Bot. 1998, 49, 387–398. [Google Scholar] [CrossRef]
- Sezen, S.M.; Yazar, A.; Dasgan, Y.; Yucel, S.; Akyildiz, A.; Tekin, S.; Akhoundnejad, Y. Evaluation of crop water stress index (CWSI) for red pepper with drip and furrow irrigation under varying irrigation regimes. Agric. Water Manag. 2014, 143, 59–70. [Google Scholar] [CrossRef]
- Kirnak, H.; Irik, H.A.; Unlukara, A. Potential use of crop water stress index (CWSI) in irrigation scheduling of drip-irrigated seed pumpkin plants with different irrigation levels. Sci. Hortic. 2019, 256, 108608. [Google Scholar] [CrossRef]
- Munns, R. Physiological processes limiting plant growth in saline soils: some dogmas and hypotheses. Plant Cell Environ. 1993, 16, 15–24. [Google Scholar] [CrossRef]
- Braunworth, W.S.; Mack, H.J. The possible use of the crop water-stress index as an indicator of evapotranspiration deficits and yield reductions in sweet corn. J. Am. Soc. Hortic. Sci. 1989, 114, 542–546. [Google Scholar]
- Colak, Y.B.; Yazar, A.; Colak, I.; Akca, H.; Duraktekin, G. Evaluation of crop water stress index (CWSI) for eggplant under varying irrigation regimes using surface and subsurface drip systems. Agric. Agric. Sci. Procedia 2015, 4, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.C. Scheduling irrigations for soybeans with the crop water-stress index (CWSI). Field Crop Res. 1990, 23, 103–116. [Google Scholar] [CrossRef]
- Smith, R.C.G.; Barrs, H.D.; Fischer, R.A. Inferring stomatal-resistance of sparse crops from infrared measurements of foliage temperature. Agric. For. Meteorol. 1988, 42, 183–198. [Google Scholar] [CrossRef]
- Egea, G.; Nortes, P.A.; Gonzalez-Real, M.M.; Baille, A.; Domingo, R. Agronomic response and water productivity of almond trees under contrasted deficit irrigation regimes. Agric. Water Manag. 2010, 97, 171–181. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Fereres, E. Establishing an almond water production function for California using long-term yield response to variable irrigation. Irrig. Sci. 2017, 35, 169–179. [Google Scholar] [CrossRef]
- Kang, J.; Hao, X.; Zhou, H.; Ding, R. An integrated strategy for improving water use efficiency by understanding physiological mechanisms of crops responding to water deficit: Present and prospect. Agric. Water Manag. 2021, 255, 107008. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Whole Growth Season (1 May–25 September) | Study Period (22 June–5 September) |
|---|---|---|
| Mean air temperature (°C) | 19.51 | 21.18 |
| Maximum air temperature (°C) | 27.22 | 29.19 |
| Minimum air temperature (°C) | 12.05 | 13.64 |
| Vapor pressure (kPa) | 1.22 | 1.22 |
| Minimum relative humidity (%) | 31.91 | 34.48 |
| Wind speed (m s−1) | 0.62 | 0.40 |
| Solar radiation (W m−2) | 232.53 | 232.82 |
| No. | Slope | Intercept | Growth Stage | Sources | R2 | n | VPD Range (kPa) | Variety | Location | Latitude and Longitude | Climate |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | −2.64 | 2.75 | V | This study | 0.96 | 53 | 1.2~3.5 | ZD958 | Shiyanghe Experimental Station of China Agricultural University, Wuwei City, Gansu Province | Lat. 37°52′ N, Long. 102°50′ E | The altitude is 1581 m, the average annual temperature is 8 °, and the average annual precipitation is 164 mm. |
| −2.24 | 2.81 | R | 0.96 | 29 | 1.9~3.6 | ||||||
| −1.96 | 3.89 | M | 0.95 | 26 | 2.2~4.2 | ||||||
| −2.62 | 2.27 | V | 0.97 | 67 | 1.2~3.9 | XY335 | |||||
| −2.28 | 2.25 | R | 0.98 | 26 | 1.8~3.6 | ||||||
| −2.16 | 3.66 | M | 0.96 | 26 | 2.1~4.2 | ||||||
| 1 | −1.90 | 2.73 | R3–R4 | Taghvaeian et al. [42] | 0.98 | 6 | 1.5~4.0 | DKC52–60, Dekalb® | Greeley, Colorado | Lat. 40°46′ N, Long. 103°2′ W | The altitude is 1166 m, the average annual temperature is 11.5 °C, and the average annual precipitation is 373 mm. |
| 2 | −1.99 | 3.04 | R–M | Taghvaeian et al. [47] | 0.97 | 12 | 1.5~4.0 | DKC52–59, Dekalb® | Greeley, Colorado | Lat. 40°26′ N, Long. 104°38′ W | The altitude is 1425 m, the average annual temperature is 11.5 °C, and the average annual precipitation is 373 mm. |
| 3 | −1.97 | 3.11 | Idso et al. [30] | 0.97 | 97 | 0.8~4.5 | Tempe, Arizona | Lat. 33°25′ N, Long. 111°56′ W | The altitude is 430 m, the average annual temperature is 16.6 °C, and the average annual precipitation is 211 mm. | ||
| 4 | −0.86 | 1.39 | R–M | Imark [32] | 0.92 | 28 | 1.0~5.5 | var. Antbey | Antalya, Turkey | Lat. 36°55’ N, Long. 34°55′ E | The altitude is 12 m, and the average annual precipitation is 1068 mm. |
| 5 | −2.56 | 1.06 | V | Yazar et al. [48] | 0.93 | 1.2~3.2 | Pioneer 3245 | Bushland, Texas | Lat. 35°11′ N, Long. 102°06′ W | The altitude is 1170 m. | |
| 6 | −1.97 | 3.43 | Han et al. [49] | 0.82 | 1.0~4.0 | Greeley, Colorado | Lat. 40°26′ N, Long. 104°38′ W | The altitude is 1427 m, the average annual temperature is 11.5 °C, and the average annual precipitation is 373 mm. | |||
| 7 | −1.79 | 2.34 | DeJonge et al. [43] | 0.97 | DCK52–04, Dekalb® | Greeley, Colorado | Lat. 40°26′ N, Long. 104°38′ W | The altitude is 1427 m, the average annual temperature is 11.5 °C, and the average annual precipitation is 373 mm. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.; Liao, Q.; Gao, S.; Kang, S.; Du, T.; Ding, R. Crop Water Stress Index as a Proxy of Phenotyping Maize Performance under Combined Water and Salt Stress. Remote Sens. 2021, 13, 4710. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13224710
Gu S, Liao Q, Gao S, Kang S, Du T, Ding R. Crop Water Stress Index as a Proxy of Phenotyping Maize Performance under Combined Water and Salt Stress. Remote Sensing. 2021; 13(22):4710. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13224710
Chicago/Turabian StyleGu, Shujie, Qi Liao, Shaoyu Gao, Shaozhong Kang, Taisheng Du, and Risheng Ding. 2021. "Crop Water Stress Index as a Proxy of Phenotyping Maize Performance under Combined Water and Salt Stress" Remote Sensing 13, no. 22: 4710. https://0-doi-org.brum.beds.ac.uk/10.3390/rs13224710