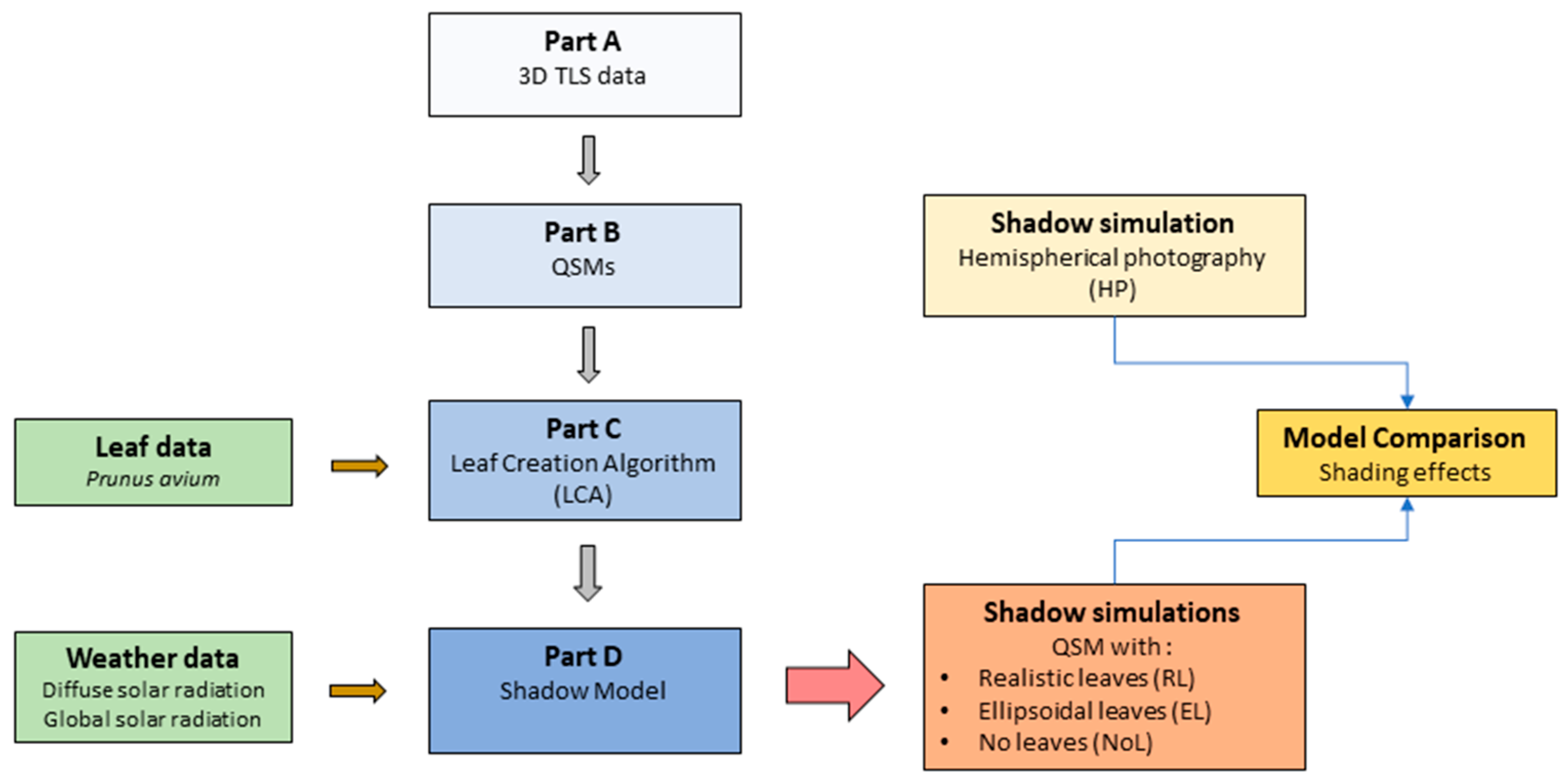
2.2. Overview of the Modelling Steps
The development of the shadow model can be divided in four parts (Parts A–D, see
Figure 1):
Part A: Collection of 3D TLS data, processing and tree segmentation;
Part B: Creation of quantitative structure models (QSMs), representative of tree structures;
Part C: Introduction of the leaf creation algorithm (LCA) and its application;
Part D: Application of the shadow model using solar radiation data.
To update our shadow model, the six wild cherry trees were scanned with a TLS Faro Focus 3D S120 (FARO Technologies, Inc.; Lake Mary, FL, USA) in early spring (April 2019), under windless and leaf-off conditions, for a better visibility of the woody compartments of the trees [
48]. The scanned tree surface is representative of the last growing season. We used a multiple scan approach with four scan positions around each tree, 90° degrees apart (azimuth angle) and at a distance of 10 m from the base of the tree. TLS device sampling parameters were set to
¼ for “resolution” and
4x for “quality”. A minimum of five reflective targets were set out around each tree to merge the multiple scans. The scans were registered using the software Faro Scene 6.2.4.30 (FARO Technologies, Inc.; Lake Mary, FL, USA) with a reported mean point error below 4 mm. Duplicated points were eliminated with a “low” search radius. In the following step, tree point clouds were manually extracted; this step was straightforward as trees had no direct neighbours or hindering obstacles. Lastly, we filtered the point clouds to remove noise points and outliers in CloudCompare v2.10.2 [
49].
The processed and filtered single-tree point clouds served as basis for reconstructing the architecture of trees using the MATLAB implementation of TreeQSM version 2.3 [
37]. In the 3D reconstruction of QSMs, trees are modelled as a hierarchical collection of cylinders (e.g., geometric primitives) fitted to local details of the single-tree point cloud.
To optimise QSMs, we tested 32 combinations of key model input parameters (among them, cover patch diameter
d, and relative cylinder length
lcyl) and produced 15 models for each possible combination of inputs to select the best model, define the optimised parameters, and to assess the robustness of the reconstruction method and uncertainty of results [
41,
50]. The mean point-cylinder-model-distance was used as suitable metric for the optimisation [
51]. All QSMs were reconstructed with the same optimised input parameters: in the first cover set
d was 15 cm; for the second cover, the minimum and maximum
d were 1 cm and 5 cm, respectively;
lcyl was 3.5. The uncertainty of QSM-parameters stayed below 10% (CV; coefficient of variation) for each of the six trees. The regular, cylindrical-like, stem base of the trees did not require additional triangulation.
The QSM-derived tree parameters for the scanned trees are presented in
Table 2. Trees were similar in terms of DBH and height, while total tree and branch volume, cylinder and branch counts, revealed the more complex structures hold by trees
Pa_2,
Pa_4 and
Pa_5. We identified
Pa_6 as the least elaborate tree structure. In addition, the count of branches and accumulated branch length are important as realistic leaves are placed along the branch-cylinders by the LCA (details in
Section 2.2 Part C).
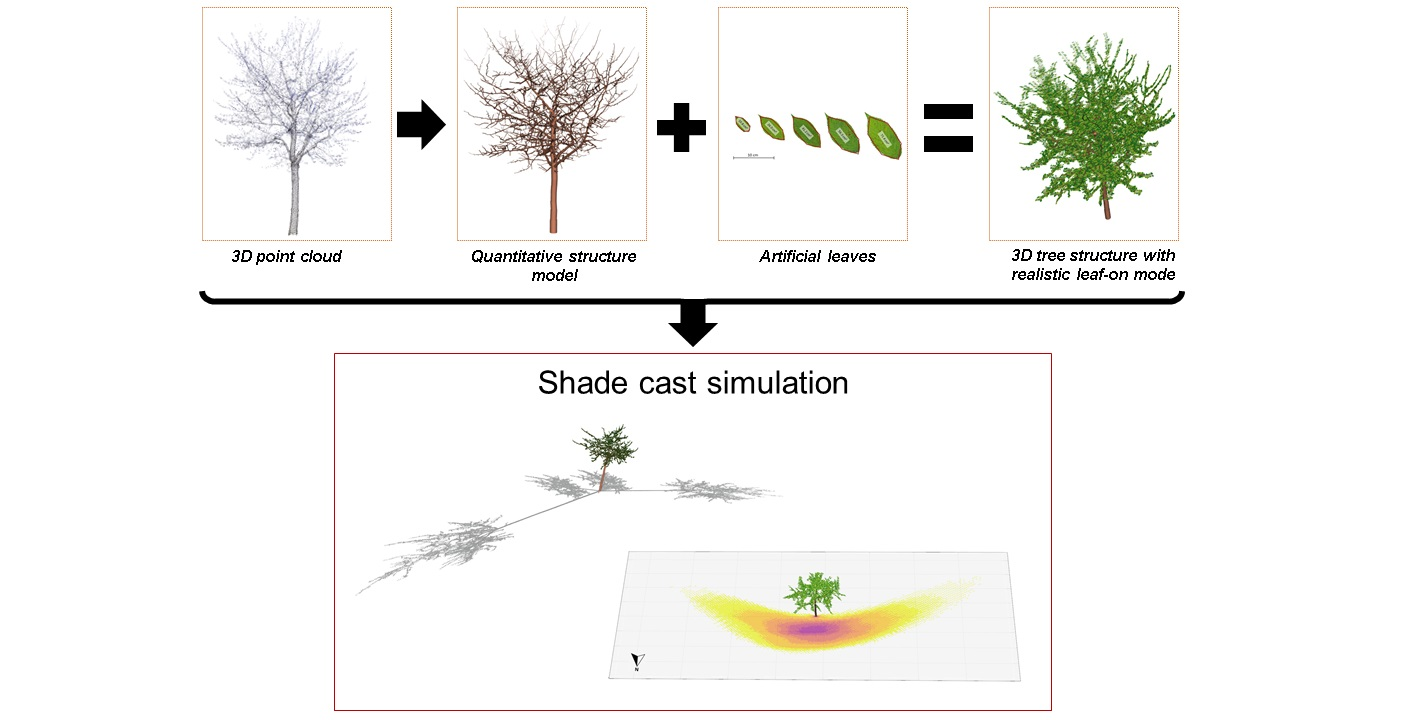
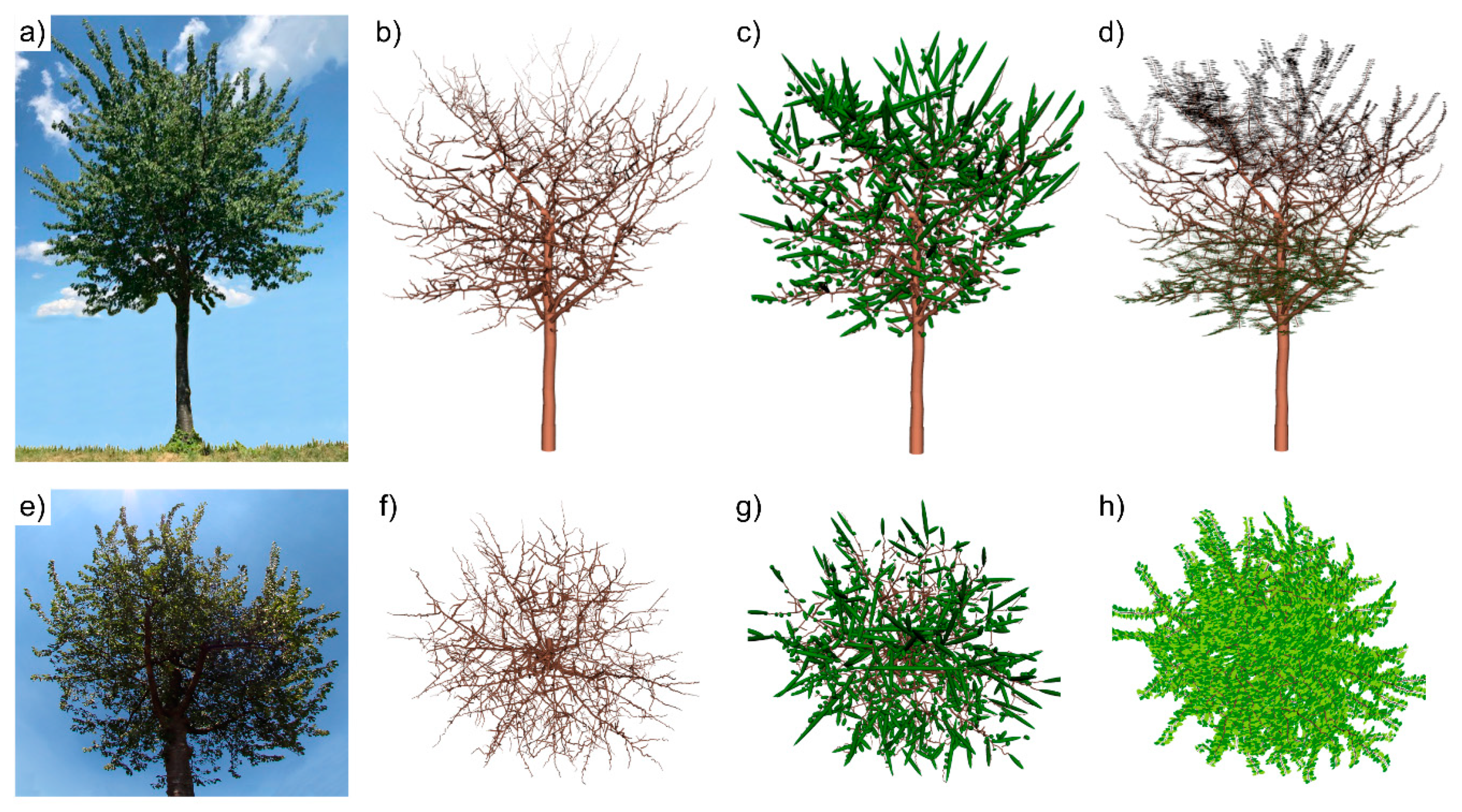
The QSMs play a central role in our shadow model, as they provide the basic topological structure to which virtually created leaves are attached. The processing steps are illustrated in
Figure 2: a photograph of one wild cherry tree (
Figure 2a); the QSM of that tree (
Figure 2b); the modelled leaf-on mode with ellipsoidal leaf-replacements (
Figure 2c); the new leaf-on mode using realistic leaves (
Figure 2d); photograph of the crown for the four models as a worm’s eye view (
Figure 2e); bird’s eye views of the QSM, QSM with ellipsoidal leaf-replacement and with realistic leaves, respectively (
Figure 2f–h). The full description of the new leaf-on mode and the details of the enhanced shadow algorithm are presented in
Section 2.2 Part C and Part D, respectively; the use of these 3D structures for simulating the shading effects is described in
Section 2.3. A video-visualisation of the 3D structure of tree
Pa_5, the QSM and leaf modes is found in
Supplementary File S1.
In July 2018, leaves were collected from felled wild cherry trees for generating data to serve as basis for the virtual creation of leaves on overlaying the QSMs in a computer environment. The realistic leaves were an attempt to represent carefully the leaf topology of wild cherry trees, and were used as the RL leaf-on mode for the shadow simulations.
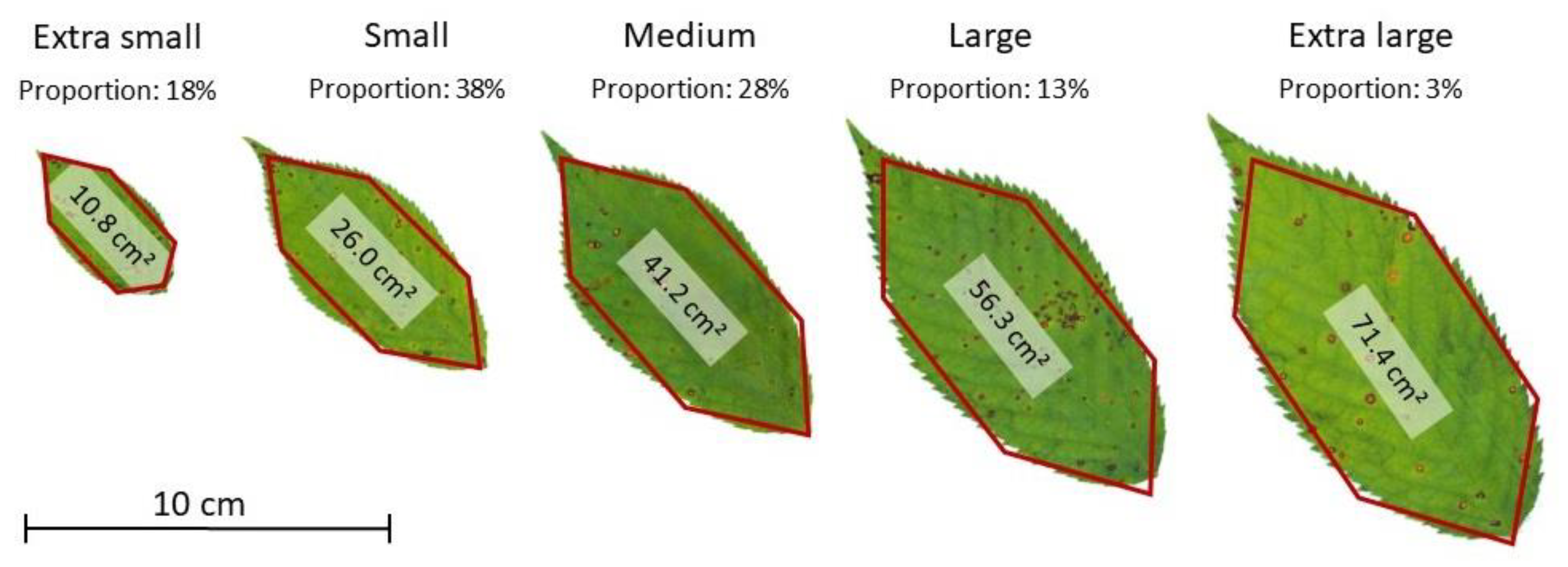
To gain insights in leaf patterns (i.e., shape distribution, size distribution, and spatial distribution within-trees), we sampled 58 branches from eight cherry trees growing in analogous conditions as the six laser-scanned trees. Branches were randomly selected from different tree heights, starting from the crown base to the tree top, and in multiple cardinal directions. We cut these branches at the branch collar, manually removed their leaves, and stored them in separate bins, which were weighed. A subset of 20 branches has been photographed with a scale to determine branch parameters, such as branch collar diameter, branch length and branch specific mass. From the leaf bins, 630 fresh leaves were randomly drawn and scanned with a ScanMaker 9800XL plus (Microtek International, Inc.; Hsinchu City, Taiwan) in a resolution of 600 dpi, to gather information of the leaf dimensions with the software ImageProPlus Version 7.0.1 (Media Cybernetics, Inc.; Rockville, MD, USA). Leaf length, leaf width and leaf area (one-sided area of the leaf blade) were measured. We divided the leaf area observations into five sets of equal length, every 15.1 cm² apart from minimum to maximum values (
Figure S1). In addition, we determined the leaf area distribution, defined five leaf size classes by taking the mean leaf area of each range (extra small, small, medium, large, extra-large) and obtained their proportions, as shown in
Figure 3. Lastly, we computed hexagonal leaf-like shapes with the geometric parameters derived from leaf measurements.
The realistic leaves were initially designed as regular six-sided polygons. We calculated leaf length and leaf width for the leaf classes by averaging minimum and maximum values for each leaf in the scanning procedure. Hence, the average lower leaf edges were at 38% of the leaf length, while the average upper leaf edges at 68%. Both lower and upper edges were placed equidistant within the minimum diameter. The area of the convex hull of the six points was calculated as leaf area. Finally, the geometric edges were adjusted by a ratio, so that the mean leaf, representative of a size class, keeps the corresponding average leaf area (
Table S1). The resulting irregular hexagons can be seen in
Figure 3. The final realistic leaves representatives of the five classes were defined by six geometric points (
Table S2). Leaf geometry, area, length and width, as well as insights on the distribution and proportion of leaves, were used to feed the leaf creation algorithm (LCA).
The LCA was developed to overlay realistic leaves on the top of a QSM generated with
TreeQSM. The distribution of leaves follows the pre-set information on leaf geometry and leaf size classes proportion, the 3D tree-cylinder model properties and the user-defined input parameter “leaf spacing”. On a tree basis, for a range of virtually created leaves with increasing leaf spacing (0.5, 1.0, 1.5, 2.0, 2.5, and 4.0 cm), preliminary results [
52] revealed the leaf spacing of 2 cm being the most suitable to match the estimated total leaf area in first order branches (mean percentual overestimation of 7%;
Figures S2 and S3). The algorithm outputs consist of two data frames: the leaf edge coordinates dataset (six points defining leaf geometry and orientation), in a coordinate system respective to the input QSM, and; an informational table containing leaf attributes (leaf number, size class, area, origin).
The steps imbedded in the LCA are described in Algorithm 1. First order branches assumed a central role in the leaf generation process, while stem-cylinders were prohibited from receiving leaves. According to the insights obtained on leaf distribution and from the starting position of a first order branch, branch-cylinders were foliated if distanced in magnitude equal or greater to 8.47 % of the length of the respective first order branch (
Figure S4). Selected branch-cylinders were given leaf positions according to leaf spacing and an alternate distichous phyllotaxy (right and left arrangement). A virtual petiole (the stalk that attaches the leaf blade to the branches) of 2 cm was set from the initial leaf position. Furthermore, a random leaf area class was selected and the leaf-bottom point was matched with the extended leaf position. The other five leaf-geometry points were calculated by keeping leaf azimuth angle perpendicular to the cylinder direction (left or right direction) and leaf blade parallel to the ground plane. The six leaf-geometry points were presented in XYZ coordinates relative to the input QSM. The leaf edge coordinates output dataset was used as input within the shadow model, jointly with the tree-cylinder model.
| Algorithm 1 Leaf Creation Algorithm (LCA) |
| 1: | Load a tree-cylinder model (QSM) |
| 2: | Define parameter leaf spacing (i.e., 2 cm) |
| 3: | for each first order branch do |
| 4: | define branch section to be foliated (e.g., the first 8.47% of the length of a first order branch have no leaves). |
| 5: | for each branch cylinder to be foliated do |
| 6: | Establish positions for leaves along the directional axis between cylinder start and end, according to the parameter leaf spacing, and evenly distribute them, alternating between left and right side. |
| 7: | Expand the preliminary leaf position with 2 cm (adding a virtual petiole), perpendicular to cylinder direction and horizontal to the ground, respecting the right or left orientation (+90° or −90° from cylinder direction) |
| 8: | for each leaf position do |
| 9: | Randomly select a leaf-size class and match the lower leaf-geometry point with the established leaf position. |
| 10: | Propagate the other five leaf-geometry points by keeping leaf oriented perpendicular to cylinder direction and horizontal to the ground. |
| 11: | end for (step 8) |
| 12: | end for (step 5) |
| 13: | end for (step 3) |
| 14: | return leaf edge coordinates dataframe and leaf attributes table |
A descriptive summary of the leaves created for the shadow simulations (presented in
Section 2.3) is found within
Table 3. The total leaf area correlates well with branch volume and other tree properties (i.e., cumulative branch length), and varied considerably between trees. Likewise, total leaf count ranged from 12,539 (
Pa_6) to 35,194 (
Pa_5). The proportions of leaves found for each leaf-size class confirmed our LCA is not biased.
We refined the shadow model proposed by [
32] in two major aspects: (1) we integrated the output of the LCA to simulate the shading effects of trees (QSMs with realistic leaves); and (2) we expanded the functionalities of the algorithm to be compatible with the data-frame structure and building-logic of TreeQSM (previously based on SimpleTree), including the module related to the creation of ellipsoidal leaves (EL). The updated shadow model includes the realistic leaves following the same principles of the initial method, where cylinders and ellipsoids (as leaf-replacements) are taken as structures to project shadows on the ground under a given sun position.
For simulations of the EL leaf-on mode, we modified the cylinder-radius threshold for ellipsoids, proposed by Rosskopf et al. [
32], from 0.5 to 1.0 cm (algorithm parameter defining where leaf-replacements are to be built upon) and adjusted the length of the longer/major ellipsoids axis, while the minor axis was set to 5.0 cm. Apart of the size limitation, ellipsoids were created for each set of cylinders in a branch order, where the threshold applied. At the given level of implementation, a change in size to simulate leaf growth was not needed as the shade cast was modelled for the leaf attributes given in the month of July 2019.
For modelling the insolation on the ground plane, we used measured data of solar irradiance (global radiation and diffuse radiation) provided by the German Meteorological Service (DWD) [
53] and chose the nearest meteorological station located in Freiburg (48°01′12.0″N 7°49′48.0″E, 236.5 m a.s.l.), about 15 km from the location of the scanned trees. The solar radiation data for July 2019, with a temporal resolution of 10 min, was integrated within the shadow model.
In summary, the updated shadow model was implemented in the open source language R, version 3.5.3 [
54], and is based on key functions of the R packages “sp” [
55,
56] and “insol” [
57]. In addition, we utilised functions of the package “rgl” [
58] for visualisation purposes.
2.4. Comparisons and Analysis of Shading Effects
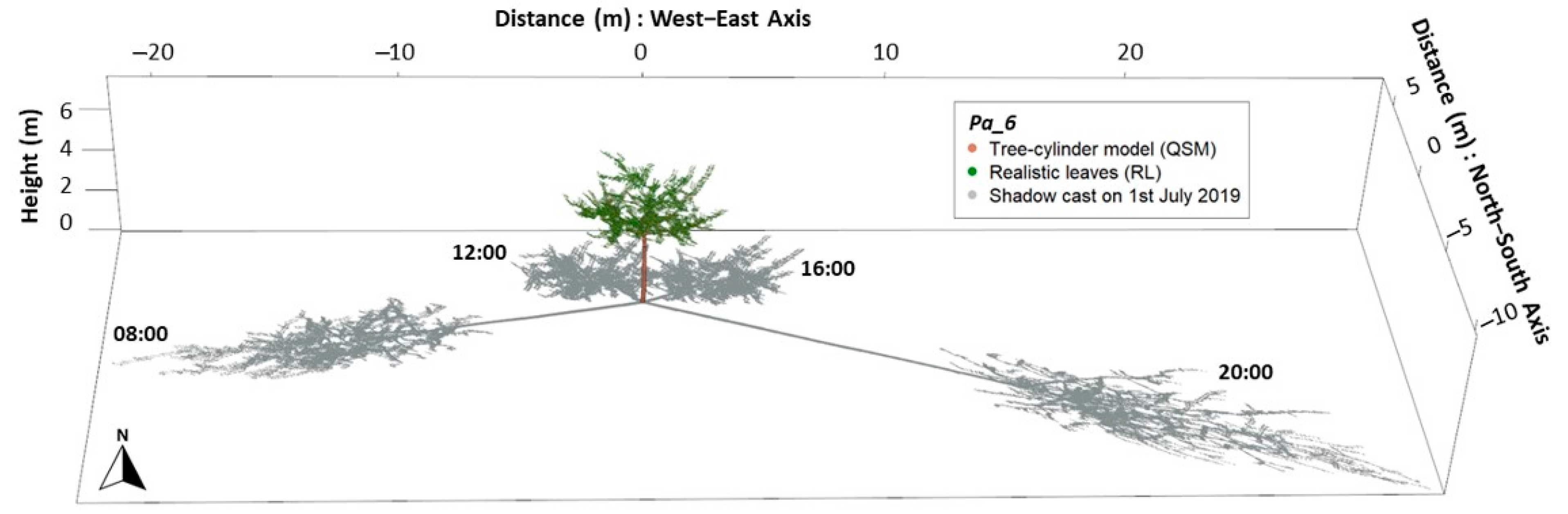
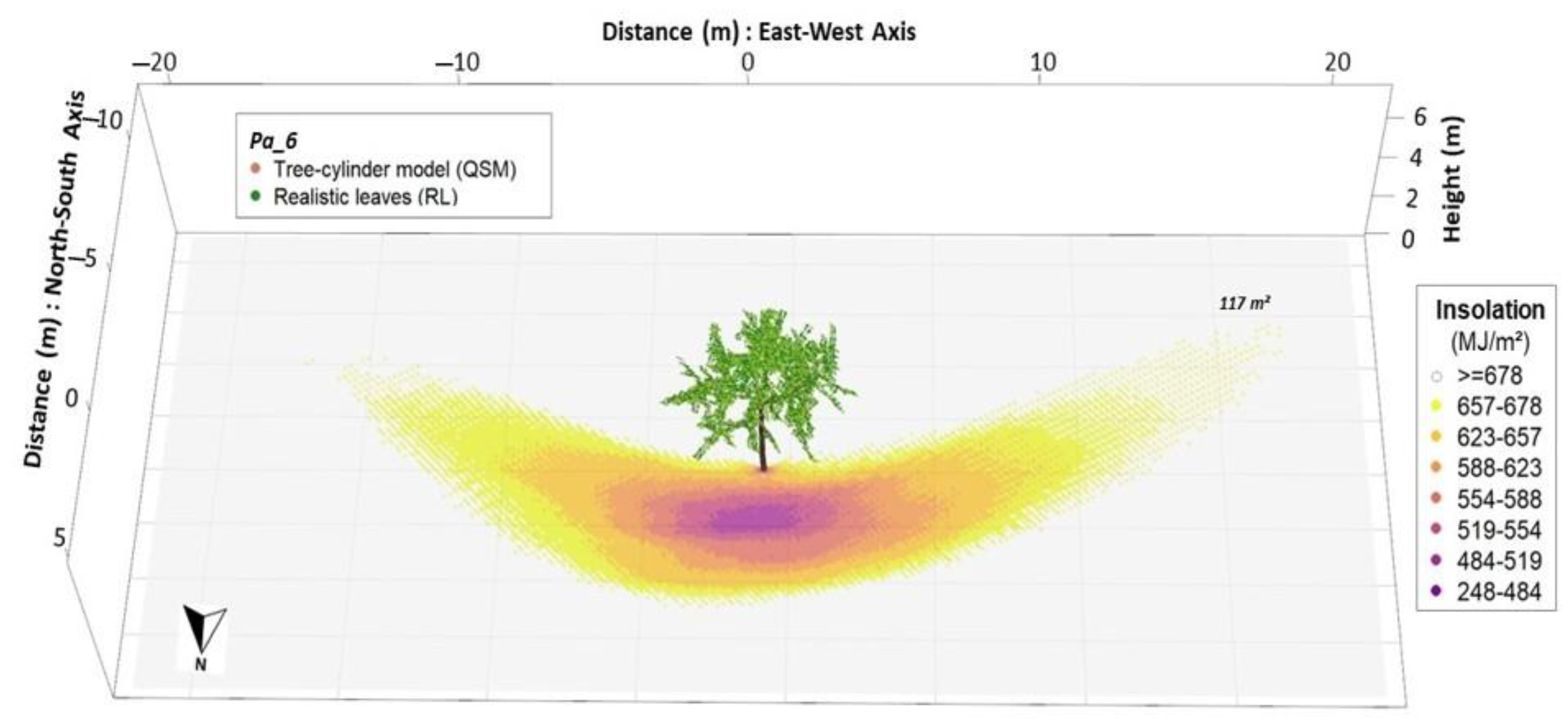
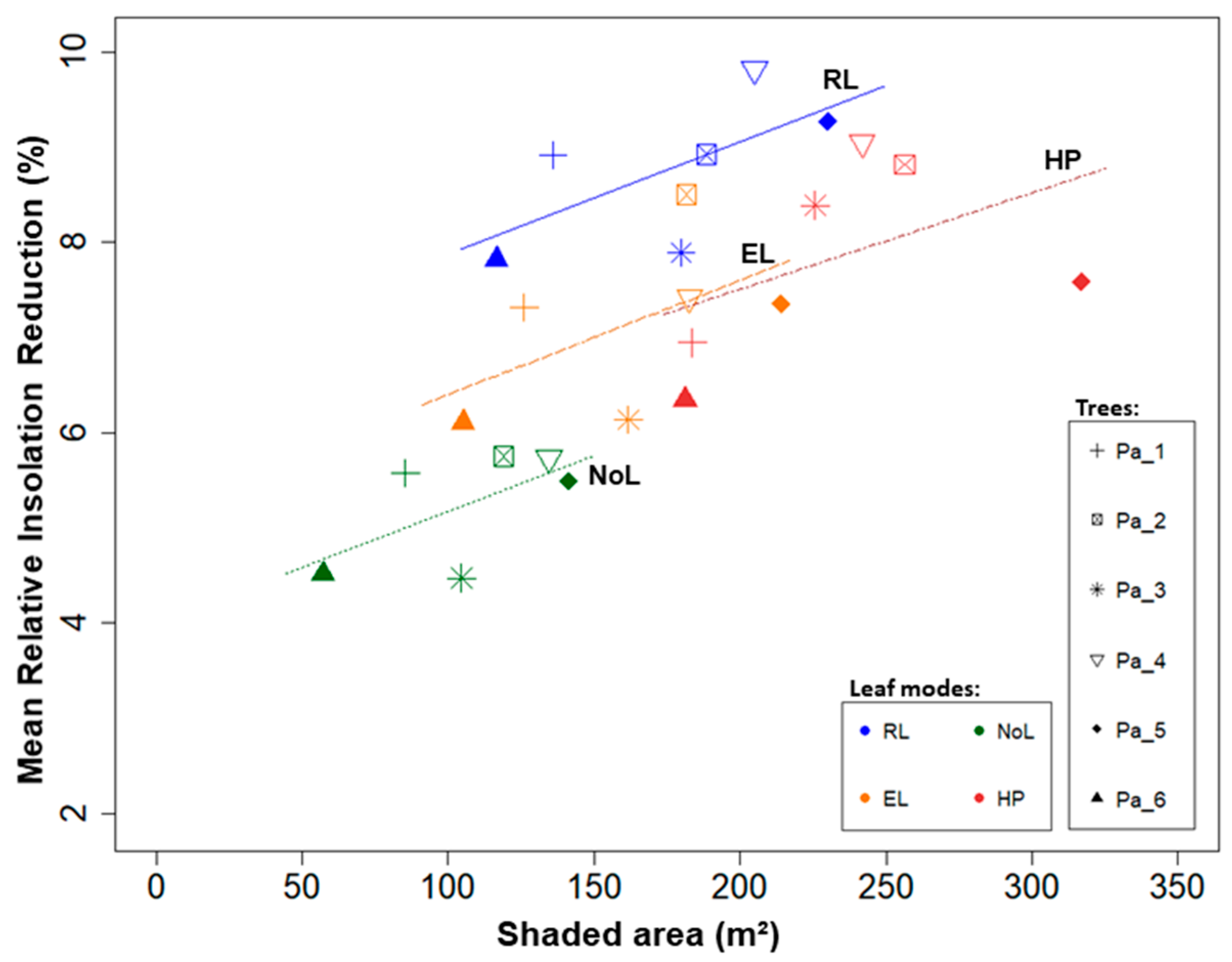
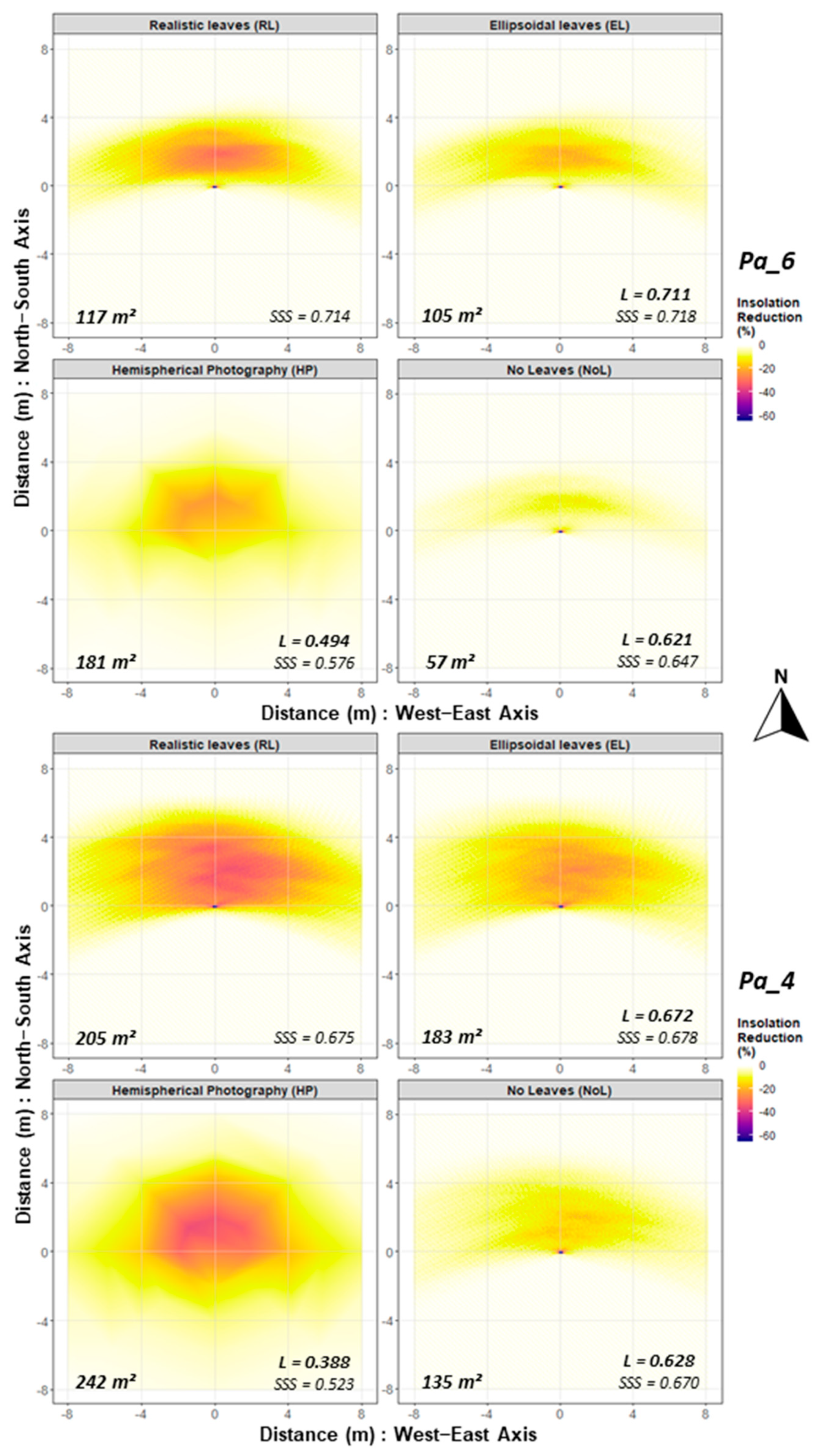
We explored the heterogeneity of the modelled shading effects in terms of monthly insolation, and compared it to full radiation conditions to estimate the shading effect: the total shaded area, and the absolute and relative insolation reduction.
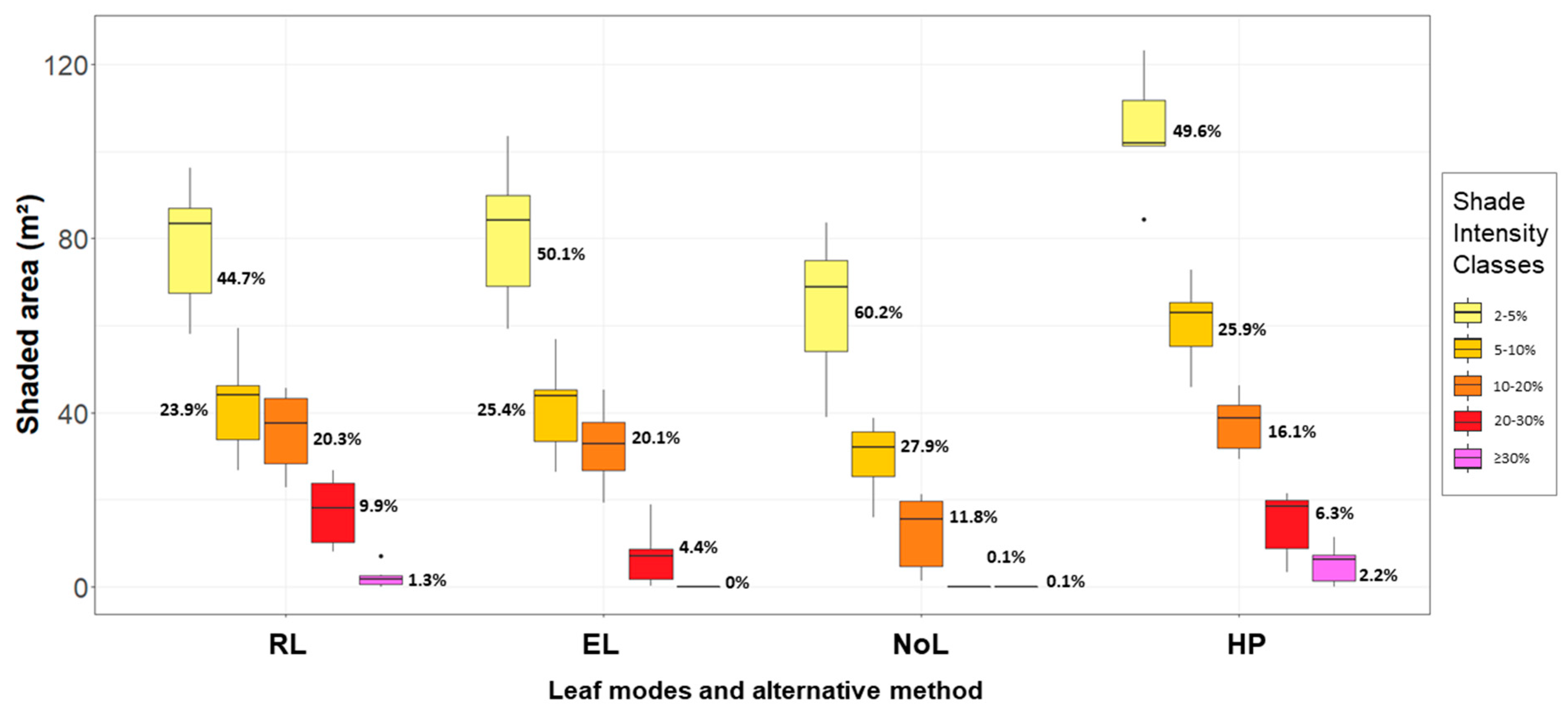
The shaded area was defined as the area sum of grid cell receiving less than 98% of the maximum possible insolation (cells with insolation reduction), whereas the remaining cells were defined under full light conditions (no decrease in insolation). On each shaded area, total insolation reduction was calculated as the sum of differences between the maximum radiant energy potentially available on a grid cell and the actual radiant energy found on it. Furthermore, we investigated the shaded area proportion under different shade intensities (in percent), as defined:
where
x1 = sum of 10-min incoming radiation energy per grid cell over simulation period (31 days), under possible shaded conditions (insolation <98% of the maximum possible insolation);
x2 = sum of 10-min incoming radiation energy per grid cell over simulation period (31 days), under full light conditions (insolation ≥98% of the maximum possible insolation).
Shade intensity values were split in classes of relative insolation reduction, as the intensity of shading have varied effects on the establishment and productivity of agricultural crops [
9]. The five shade intensity classes were 2–5%; 5–10%; 10–20%; 20–30%; and >30% insolation reduction. We calculated the results for each tree under each of the four treatments (RL, EL, NoL, and HP). Lastly, we investigated the spatial heterogeneity for pairs of observations taking RL as references (RL against EL, NoL and HP).
In order to parameterise the bivariate spatial dependence, and to test the similarity of the spatial patterns of the shading effects, we calculated the bivariate association measure
L [
60,
61]. For these spatial analyses, we used a subset data corresponding to a grid area of 128 m² (rectangle of 16 m × 8 m), encompassing the majority of the insolation reduction on ground, from the tree trunk 2 m to the south, 6 m to the north, and 8 m towards west and east directions.
The
L measure, the univariate spatial association measures (SSSx and SSSy) and correlation coefficients associated to it, were estimated with the “spdep” library [
55] in the R environment v 3.5.3 [
54]. For the insolation reduction grid data, neighbours were created with the “cell2nb” function and the type of sharing boundary connectivity was set to “queen”; weights were given with the “nb2listw” function, and the globally standardised coding scheme style (“C”) was chosen [
62]. A permutation test for the
L measure (400 random permutations, “lee.mc” function) of
x and
y (RL and EL, NoL or HP shading effects, respectively) for the given spatial weighting scheme established the rank of the observed statistic and calculated “pseudo
p-values”.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}