Feasibility of Invasive Grass Detection in a Desertscrub Community Using Hyperspectral Field Measurements and Landsat TM Imagery


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Background
- (1)
- From the standpoint of cover, P. ciliare replaces soil. Sub-canopy P. ciliare is less likely to have a profound effect on reflectance of a mixed pixel.
- (2)
- Native grasses do not form dense stands in the upland habitats in which P. ciliare is invading.
- (3)
- P. ciliare is visible from a distance with the human eye at different times of the year.

1.2. Objectives
- (1)
- Identify spectral characteristics that distinguish P. ciliare from uninvaded Arizona Upland cover types throughout the year
- (2)
- Determine best time of year to discriminate between P. ciliare and uninvaded Arizona Upland vegetation
- (3)
- Assess the potential of multi-date analysis to improve upon single-date analysis to discriminate between P. ciliare and uninvaded Arizona Upland vegetation
2. Data and Methods
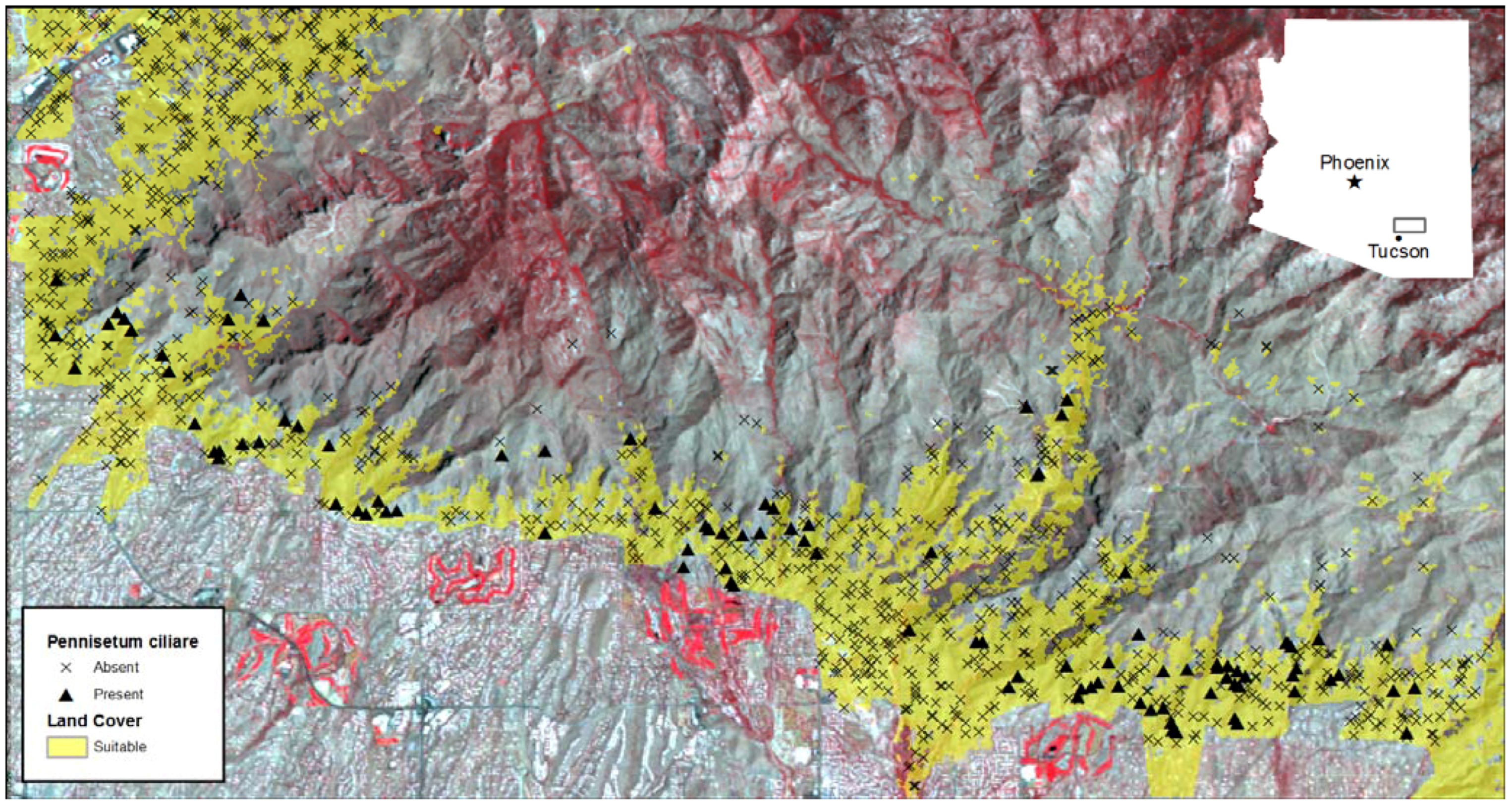
2.1. Study Area

2.2. Field Data Collection—Cover Measurements
2.3. Field Data Collection—Spectral Data Acquisition
2.3. Spectral Separability of Pennisetum ciliare from Native Cover Types
2.4. Spectral Separability of Mixed Landscapes
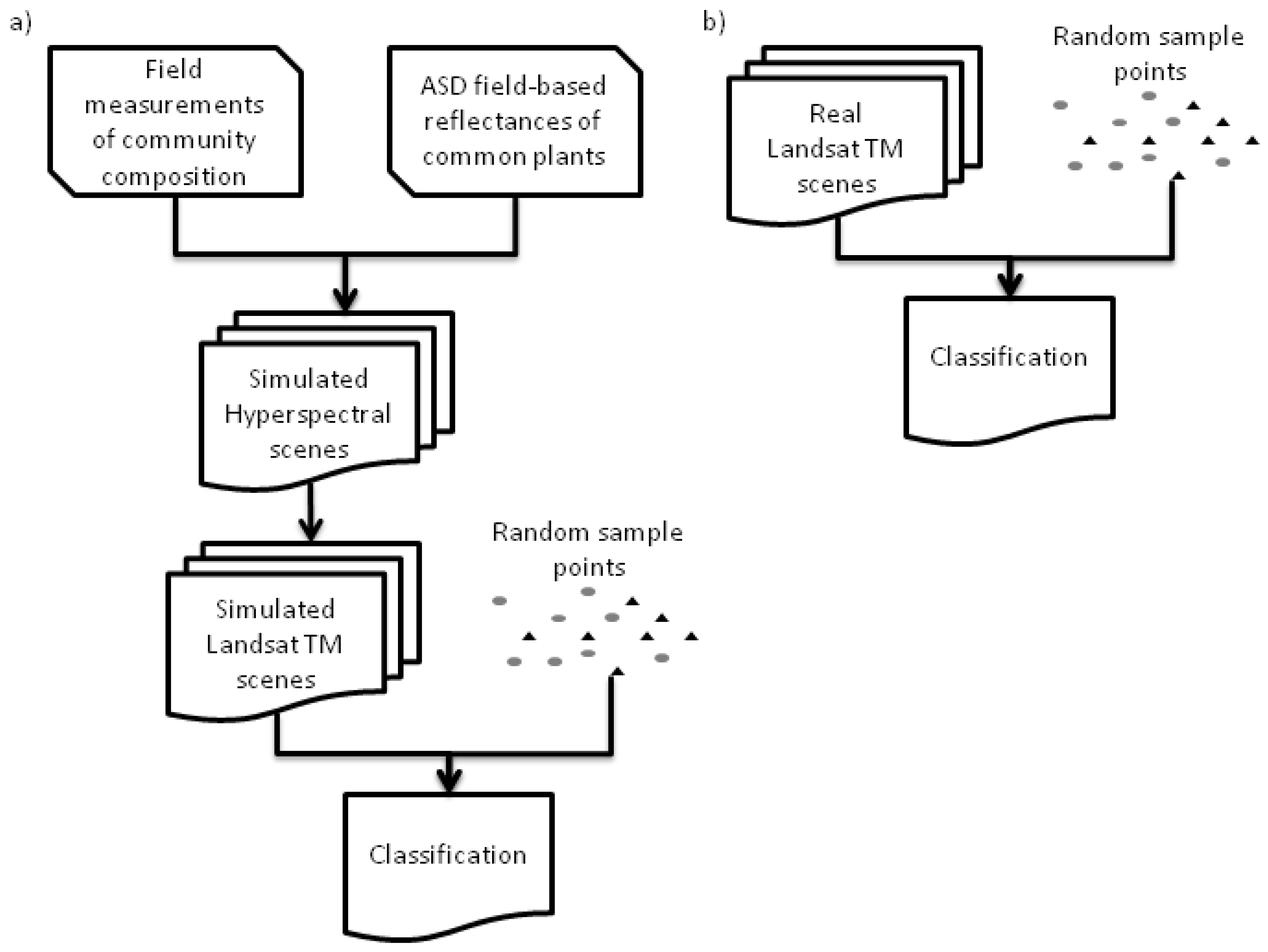
2.5. Simulated Landsat TM Scenes
2.6. Landsat TM Scenes
2.7. Landsat TM Training and Validation Sites
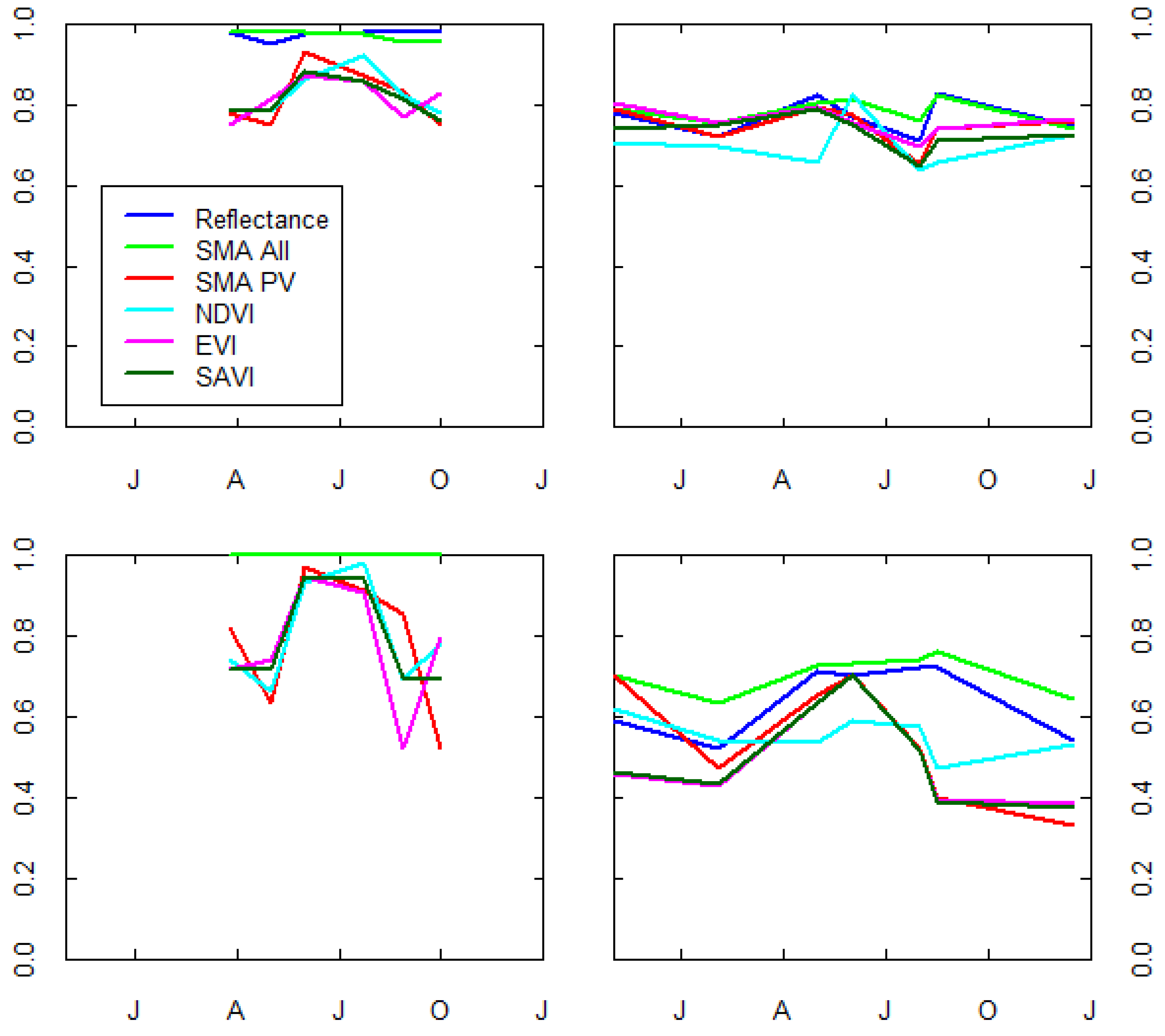
2.8. Scene Classification—Classification Data Models
- Pure reflective (Refl)
- Spectrally unmixed PV, soil, and NPV (SMAAll)
- Spectrally unmixed PV (SMAPV)
- Normalized Vegetation Difference Index (NDVI)
- Soil-adjusted Vegetation Index (SAVI)
- Enhanced Vegetation Index (EVI)
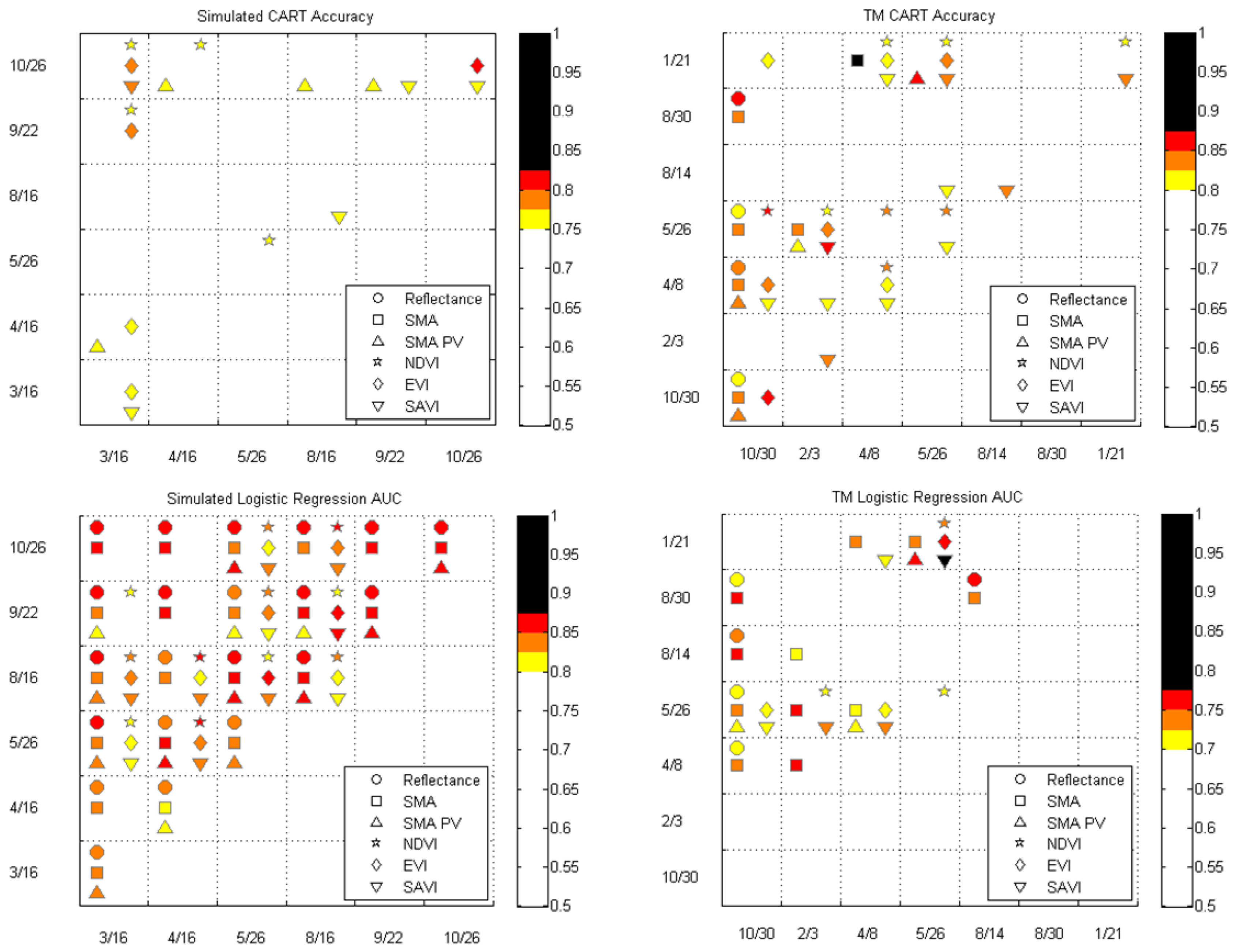
2.9. Classification and Regression Trees (CART)
2.10. Logistic Regression
2.11. Landsat TM Scene Visualization
3. Analysis of Results
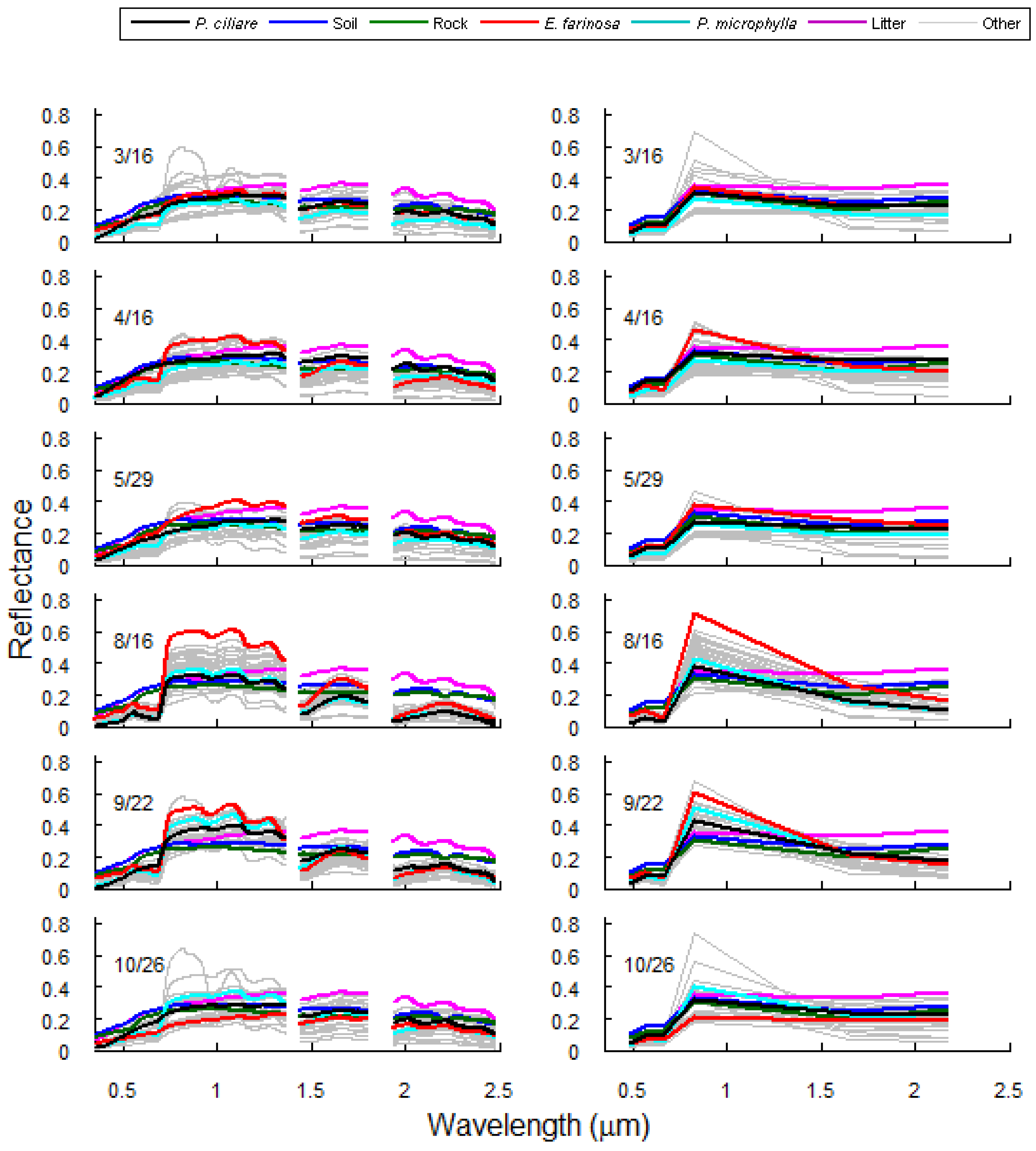
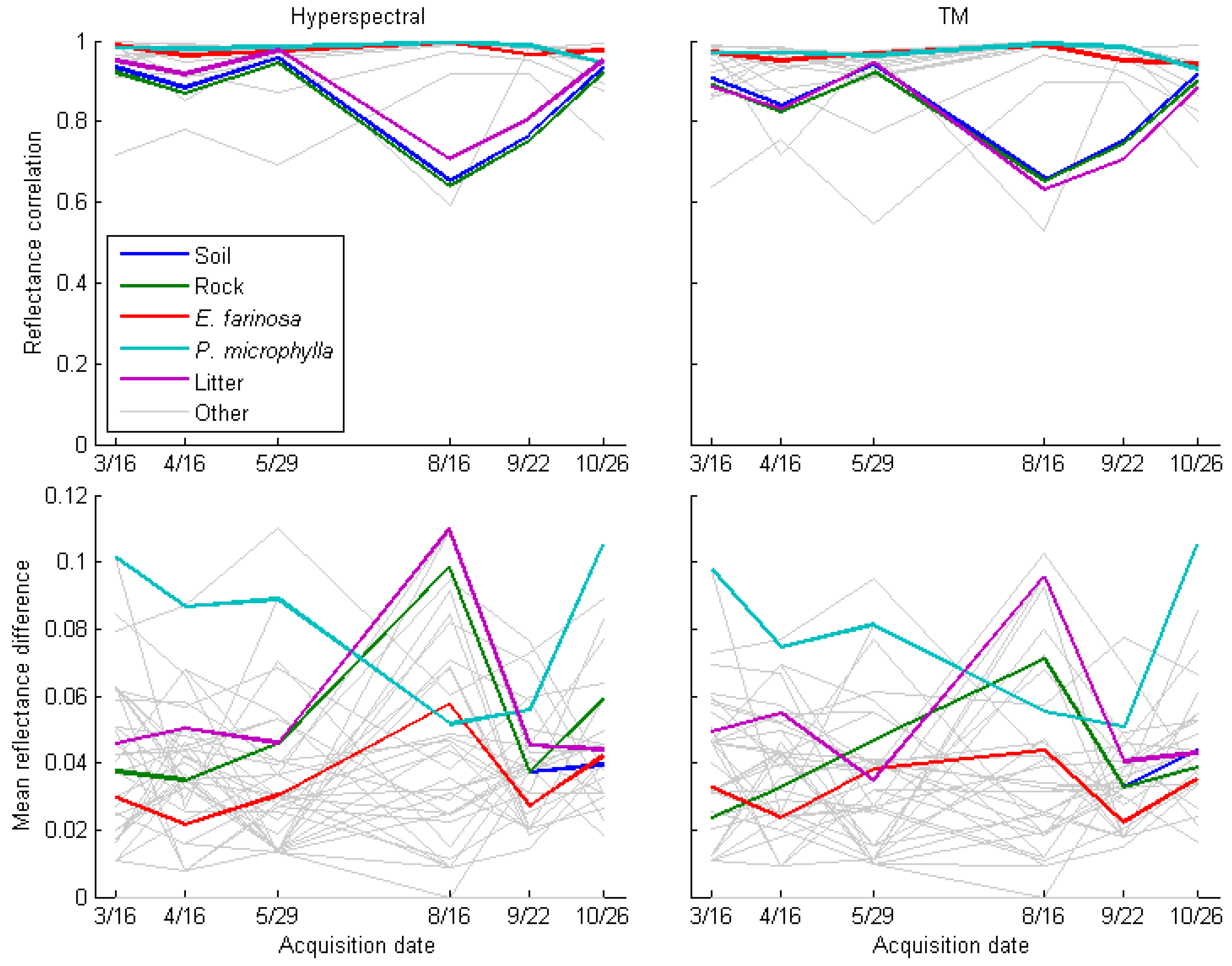
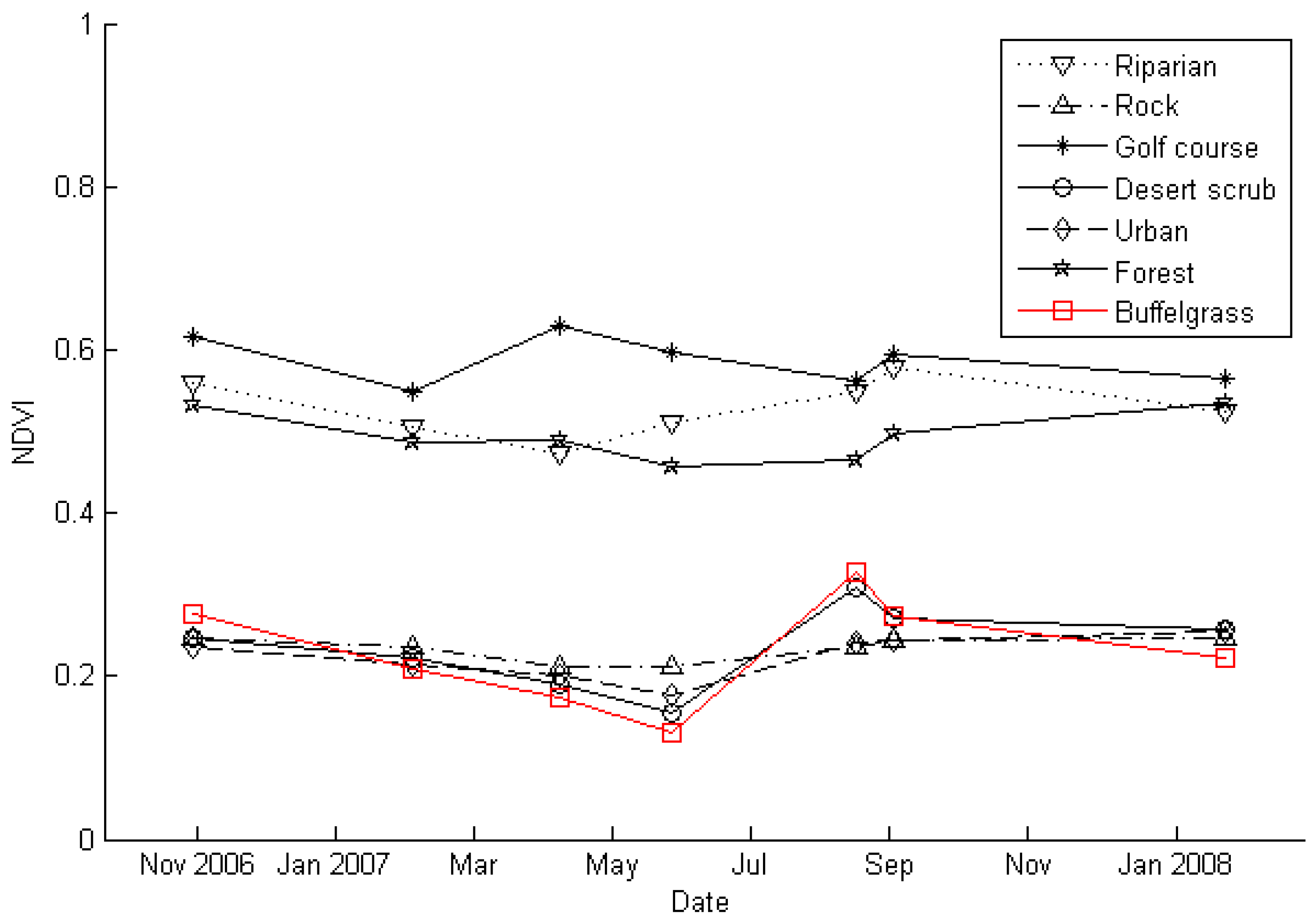
3.1. Spectral Separability of Pennisetum ciliare Over Time
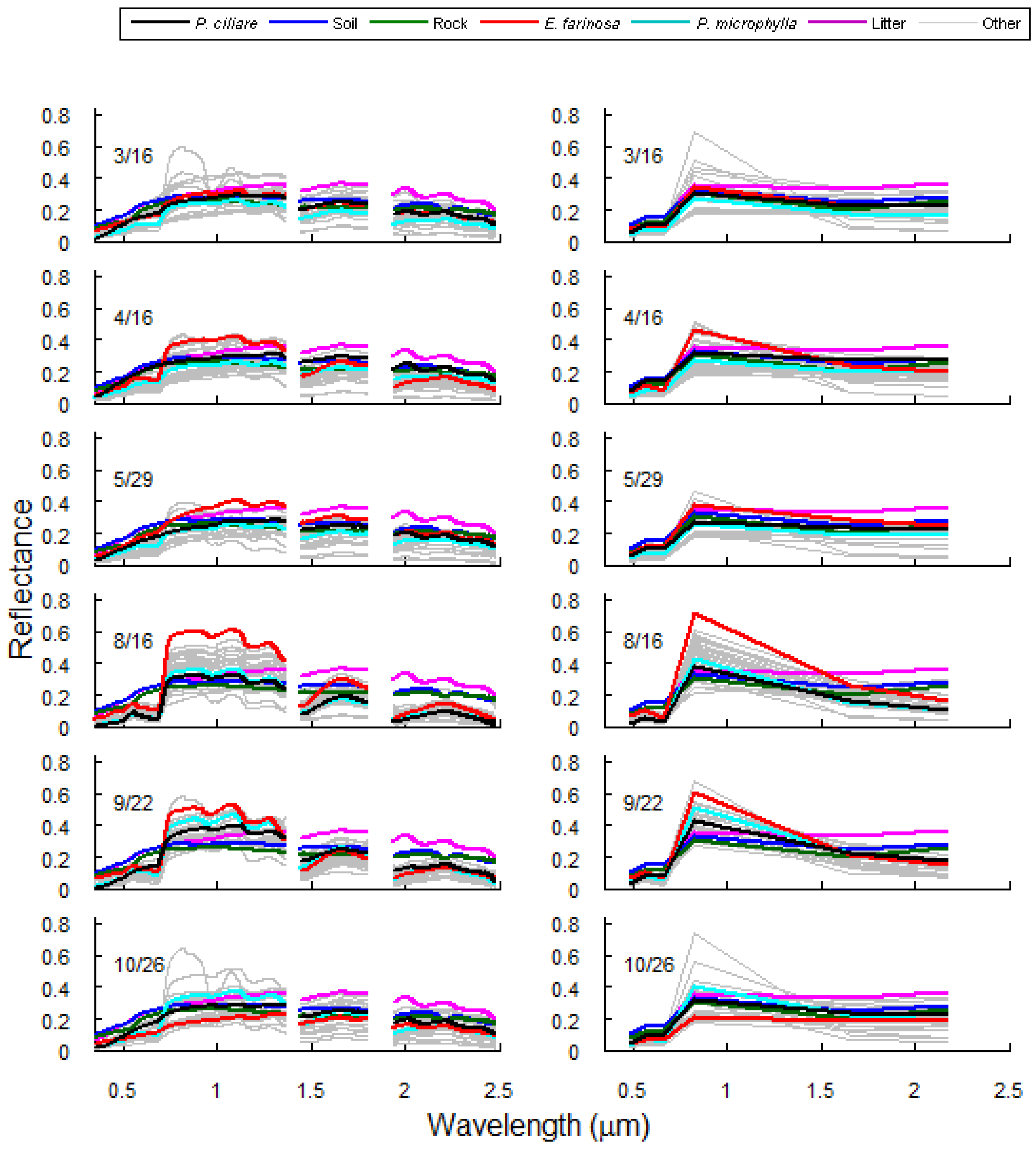
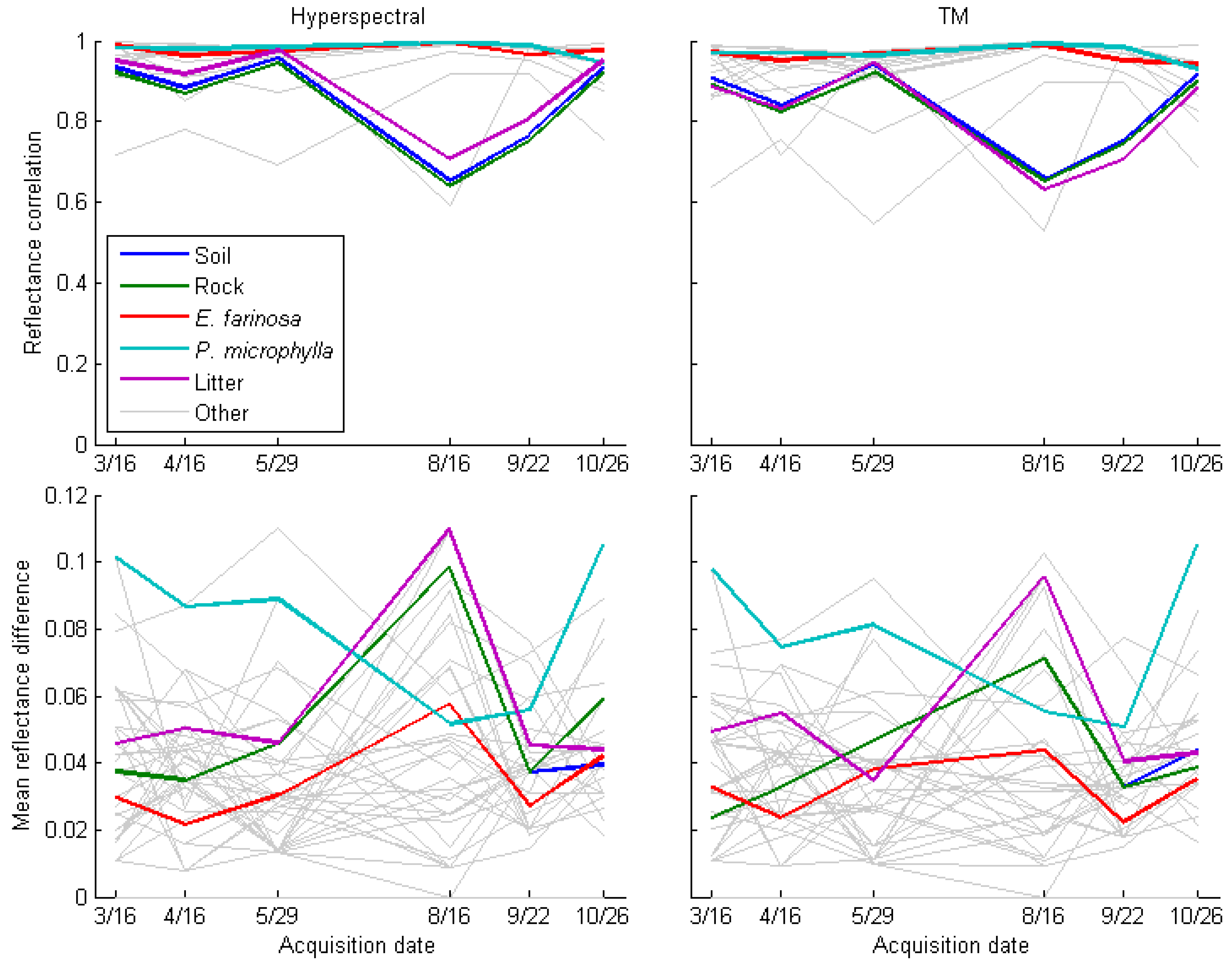

3.2. Mixed Pixel Separability of P. ciliare from Natives in Arizona Upland Landscapes
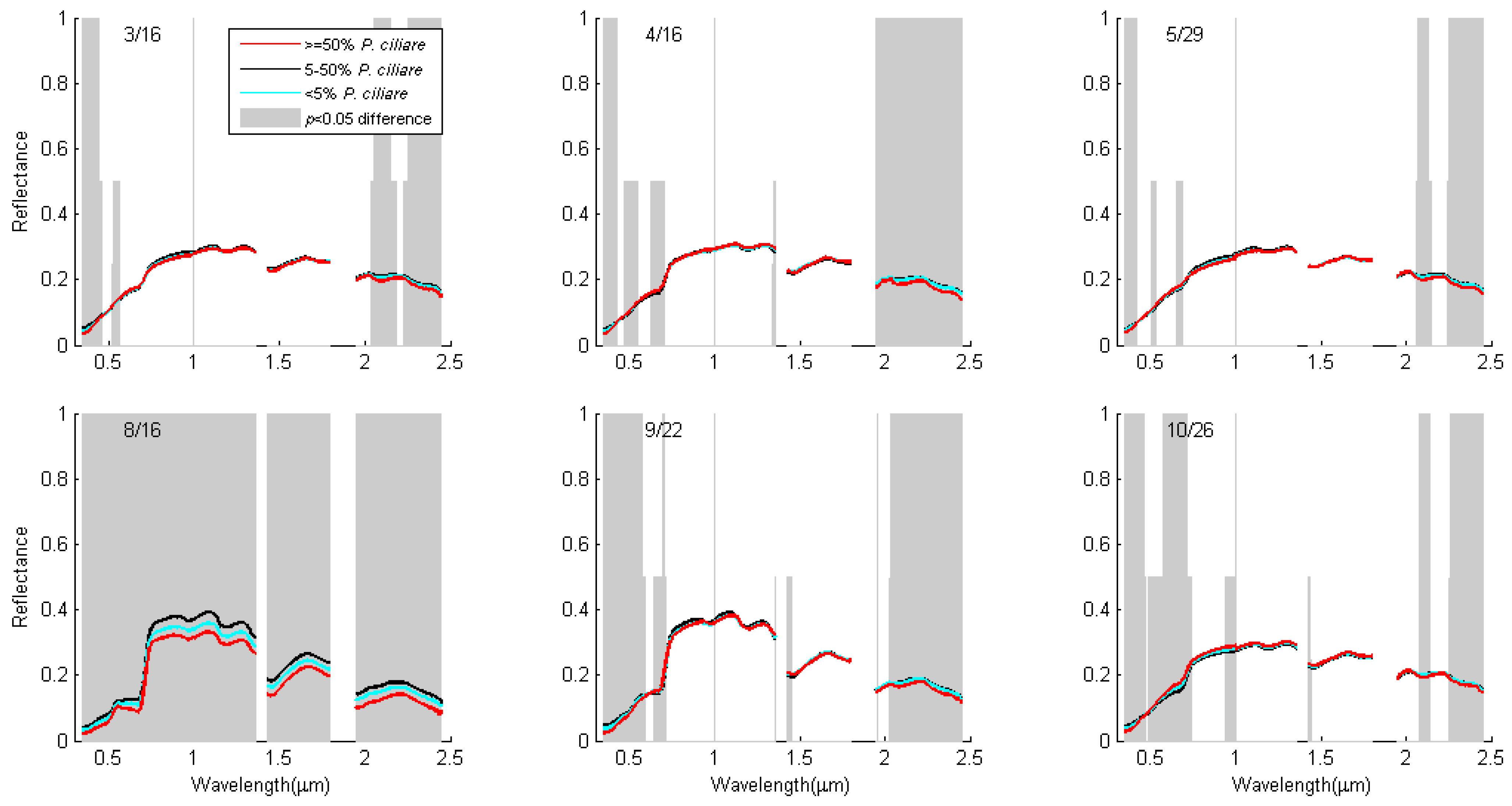
3.3. Single Date Results

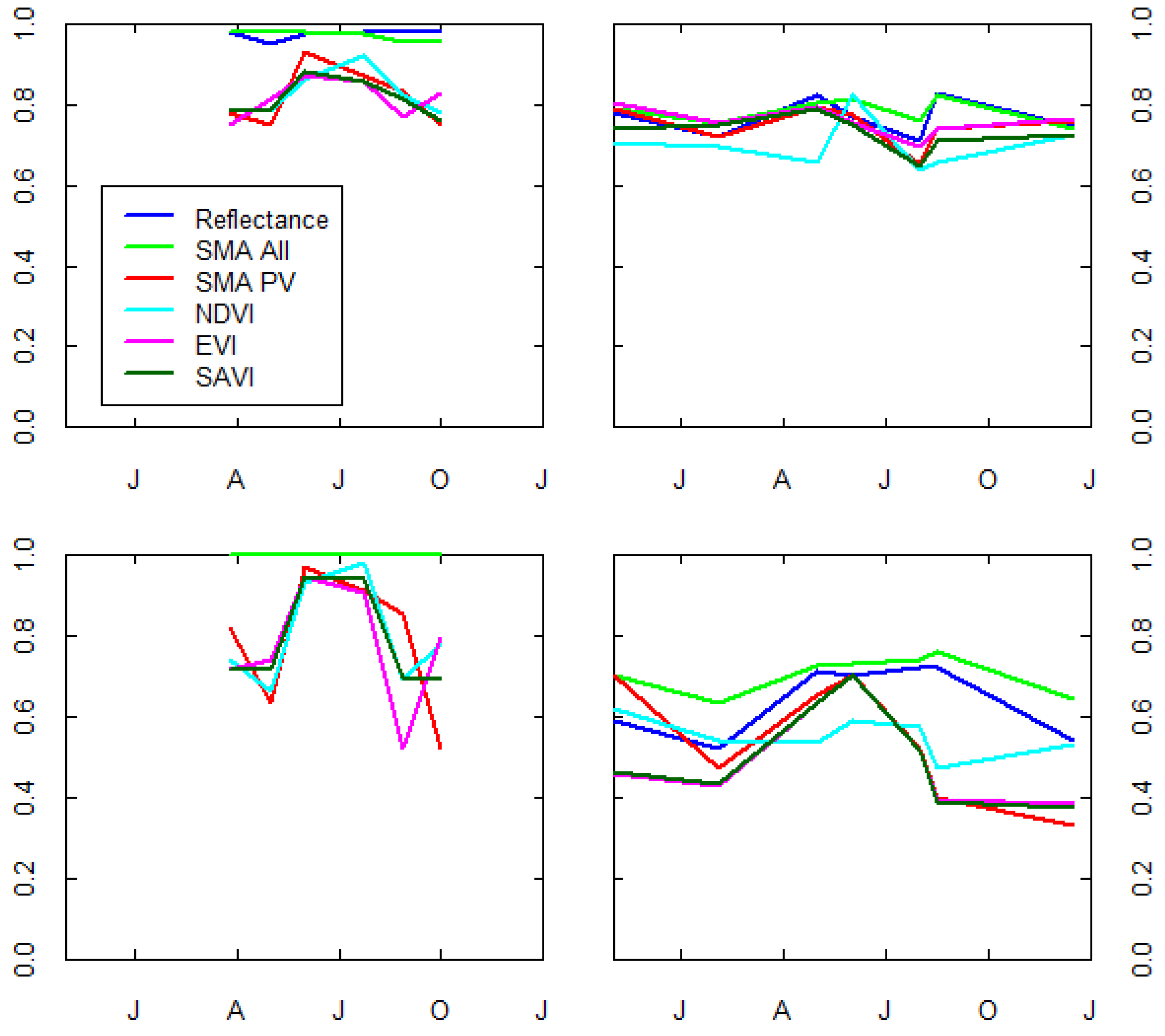
3.4. Multidate Results

4. Discussion
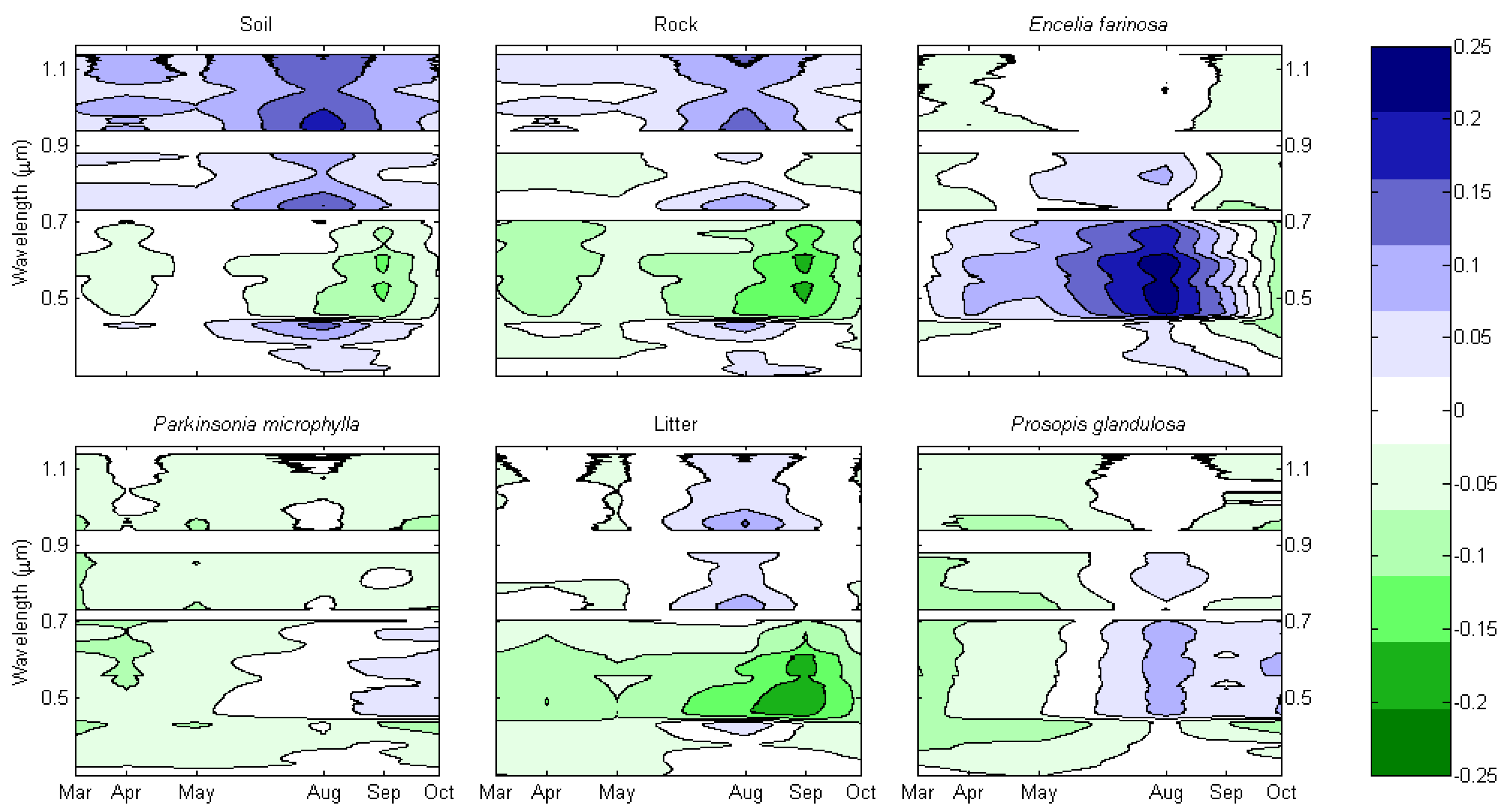
4.1. Invaded Areas are Greener than Uninvaded Areas
4.2. P. ciliare Dries out and Senesces before Native Vegetation
4.3. Invaded Areas are Redder during the Senesced Phase
4.4. Best Dates for Distinguishing P. ciliare from Native Vegetation
5. Concluding Remarks
Acknowledgements
Abbreviations:
| EVI | Enhanced vegetation index |
| MODIS | Moderate Resolution Imaging Spectroradiometer |
| NDVI | Normalized difference vegetation index |
| NPV | Non-photosynthetic vegetation |
| PV | Photosynthetic vegetation |
| SAVI | Soil-adjusted vegetation index |
| SMA | Spectral mixture analysis |
References
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass fire cycle, and global change. Ann. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. Bioscience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Mclaughlin, S.P.; Bowers, J.E. Effects of wildfire on a Sonoran Desert plant community. Ecology 1982, 63, 246–248. [Google Scholar] [CrossRef]
- Burquez-Montijo, A.M.; Miller, M.E.; Yrizar, A.M.; Tellman, B. Mexican grasslands, thornscrub, and the transformation of the Sonoran Desert b invasive exotic buffelgrass (Pennisetum ciliare). In Invasive Exotic Species in the Sonoran Region; Tellman, B., Ed.; The University of Arizona Press-The Arizona-Sonora Desert Museum: Tucson, AZ, USA, 2002; pp. 126–146. [Google Scholar]
- Williams, D.G.; Baruch, Z. African grass invasion in the Americas: Ecosystem consequences and the role of ecophysiology. Biol. Invasions 2000, 2, 123–130. [Google Scholar] [CrossRef]
- Van Devender, T.R.; Felger, R.S.; Burquez-Montijo, A. Exotic Plants in the Sonoran Desert Region, Arizona and Sonora. In Proceedings of the California Exotic Pest Plant Council Symposium, Concord, CA, USA, 11 December 1997; Volume 3, pp. 10–15.
- Rogstad, A. The Buffelgrass Strategic Plan; Arizona-Sonora Desert Museum: Tucson, AZ, USA, 2008. [Google Scholar]
- Brenner, J. Structure, Agency, and the Transformation of the Sonoran Desert by Buffelgrass (Pennisetum ciliare): An Application of Land Change Science. Ph.D. Thesis, Clark University, Worcester, MA, USA, 2009. [Google Scholar]
- Franklin, K.A.; Lyons, K.; Nagler, P.L.; Lampkin, D.; Glenn, E.P.; Molina-Freaner, F.; Markow, T.; Huete, A.R. Buffelgrass (Pennisetum ciliare) land conversion and productivity in the plains of Sonora, Mexico. Biol. Conser. 2006, 127, 62–71. [Google Scholar] [CrossRef]
- Olsson, A.D.; Marsh, S.E.; Orr, B.J.; Guertin, D.P. Development of a Spatial Decision Support System for Buffelgrass Management for the Tucson Mountain Sector of Pima County; Pima County Natural Resources Parks and Recreation: Tucson, AZ, USA, 2009. [Google Scholar]
- Turner, R.M.; Brown, D.E. Sonoran Desert scrub. In Biotic Communities: Southwestern United States and Northwestern Mexico; Brown, D.E., Ed.; University of Utah Press: Salt Lake, UT, USA, 1994; pp. 181–221. [Google Scholar]
- Huete, A.R. A Soil-Adjusted Vegetation Index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Shreve, F.; Wiggins, I.L. Vegetation and Flora of the Sonoran Desert; Stanford University Press: Stanford, CA, USA, 1964. [Google Scholar]
- de la Fuente, E.G.; Solis, J.D.; Fitzmaurice, A.S.; Encinia, F.B.; Tristan, V.V.; Grant, W.E. Growth rate pattern of buffelgrass [Pennisetum ciliare L. (Link.) Syn. Cenchrus ciliaris L.] in Tamaulipas, Mexico. Tecnica Pecuaria En Mexico 2007, 45, 1–17. [Google Scholar]
- Olsson, A.D.; McClaran, M.P.; Marsh, S.E. Transformation rate and extent of undisturbed Sonoran desert scrub by invasive buffelgrass. Biol. Invasions 2011. in review. [Google Scholar]
- Moody, M.E.; Mack, R.N. Controlling the spread of plant invasions—The importance of Nascent Foci. J. Appl. Ecol. 1988, 25, 1009–1021. [Google Scholar] [CrossRef]
- Daughtry, C.S.T. Discriminating crop residues from soil by shortwave infrared reflectance. Agron. J. 2001, 93, 125–131. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Jackson, R.D.; Slater, P.N.; Pinter, P.J. Discrimination of growth and water-stress in wheat by various vegetation indexes through clear and turbid atmospheres. Remote Sens. Environ. 1983, 13, 187–208. [Google Scholar] [CrossRef]
- Tucker, C.J.; Newcomb, W.W.; Los, S.O.; Prince, S.D. Mean and inter-year variation of growing-season Normalized Difference Vegetation Index for the Sahel 1981–1989. Int. J. Remote Sens. 1991, 12, 1133–1135. [Google Scholar] [CrossRef]
- WRCC. Arizona Climate Summaries; Western Regional Climate Center: Reno, NV, USA, 2010. [Google Scholar]
- Lowry, J.; Ramsey, R.D.; Thomas, K.; Schrupp, D.; Sajwaj, T.; Kirby, J.; Waller, E.; Schrader, S.; Falzarano, S.; Langs, L.; et al. Mapping moderate-scale land-cover over very large geographic areas within a collaborative framework: A case study of the Southwest Regional Gap Analysis Project (SWReGAP). Remote Sens. Environ. 2007, 108, 59–73. [Google Scholar] [CrossRef]
- Bowers, J.E.; Bean, T.; Turner, R.M. Two decades of change in distribution of plants at the desert laboratory, Tucson, Arizona. Madrono 2006, 53, 252–263. [Google Scholar] [CrossRef]
- Chavez, P.S. Image-based atmospheric corrections revisited and improved. Photogramm. Eng. Remote Sensing 1996, 62, 1025–1036. [Google Scholar]
- Beyer, H.L. Hawth’s Analysis Tools v 3.27 for ArcGIS 9.x. 2004. Available online: http://www.spatialecology.com/htools (accessed on 13 October 2011).
- Vermote, E.F.; Tanre, D.; Deuze, J.L.; Herman, M.; Morcrette, J.J. Second Simulation of the Satellite Signal in the Solar Spectrum, 6S: An overview. IEEE Trans. Geosci. Remote Sens. 1997, 35, 675–686. [Google Scholar] [CrossRef]
- Adams, J.B.; Smith, M.O. Spectral mixture modeling: A new analysis of rock and soil types at the Viking lander 1 suite. J. Geophys. Res. 1986, 91, 8098–8112. [Google Scholar] [CrossRef]
- Penrose, R.; Todd, J.A. On best approximate solution of linear matrix equations. Math. Proc. Cambridge Phil. Soc. 1956, 52, 17–19. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Breiman, L. Classification and Regression Trees; Wadsworth International Group: Belmont, CA, USA, 1984. [Google Scholar]
- Ramsey, F.L.; Schafer, D.W. The Statistical Sleuth: A Course in Methods of Data Analysis, 2nd ed.; Duxbury Press: Pacific Grove, CA, USA, 2002. [Google Scholar]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conser. 1997, 24, 38–49. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Olsson, A.D.; Van Leeuwen, W.J.D.; Marsh, S.E. Feasibility of Invasive Grass Detection in a Desertscrub Community Using Hyperspectral Field Measurements and Landsat TM Imagery. Remote Sens. 2011, 3, 2283-2304. https://0-doi-org.brum.beds.ac.uk/10.3390/rs3102283
Olsson AD, Van Leeuwen WJD, Marsh SE. Feasibility of Invasive Grass Detection in a Desertscrub Community Using Hyperspectral Field Measurements and Landsat TM Imagery. Remote Sensing. 2011; 3(10):2283-2304. https://0-doi-org.brum.beds.ac.uk/10.3390/rs3102283
Chicago/Turabian StyleOlsson, Aaryn D., Willem J.D. Van Leeuwen, and Stuart E. Marsh. 2011. "Feasibility of Invasive Grass Detection in a Desertscrub Community Using Hyperspectral Field Measurements and Landsat TM Imagery" Remote Sensing 3, no. 10: 2283-2304. https://0-doi-org.brum.beds.ac.uk/10.3390/rs3102283