
Weak Environmental Controls of Tropical Forest Canopy Height in the Guiana Shield

 , and
, and
Abstract
:
1. Introduction
- What is the scale and magnitude of spatial autocorrelation of tropical forest heights?
- Are there links between environmental variables and canopy heights in tropical forests and do these links depend on the spatial resolution of the LiDAR-derived maps?
- Are environmental variables important drivers of tropical forest canopies and do they bring useful information in the perspective of predictive mapping?
2. Material and Methods
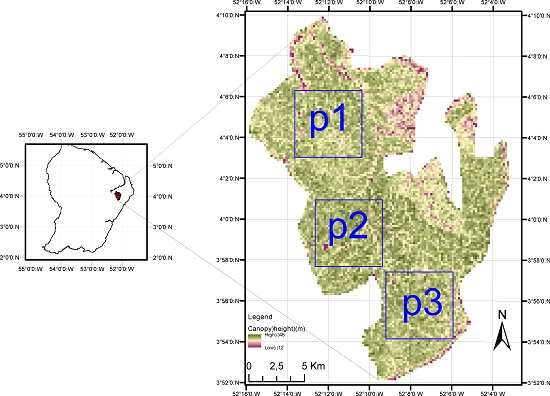
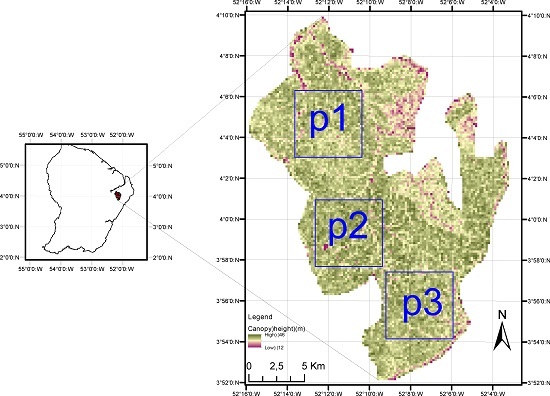
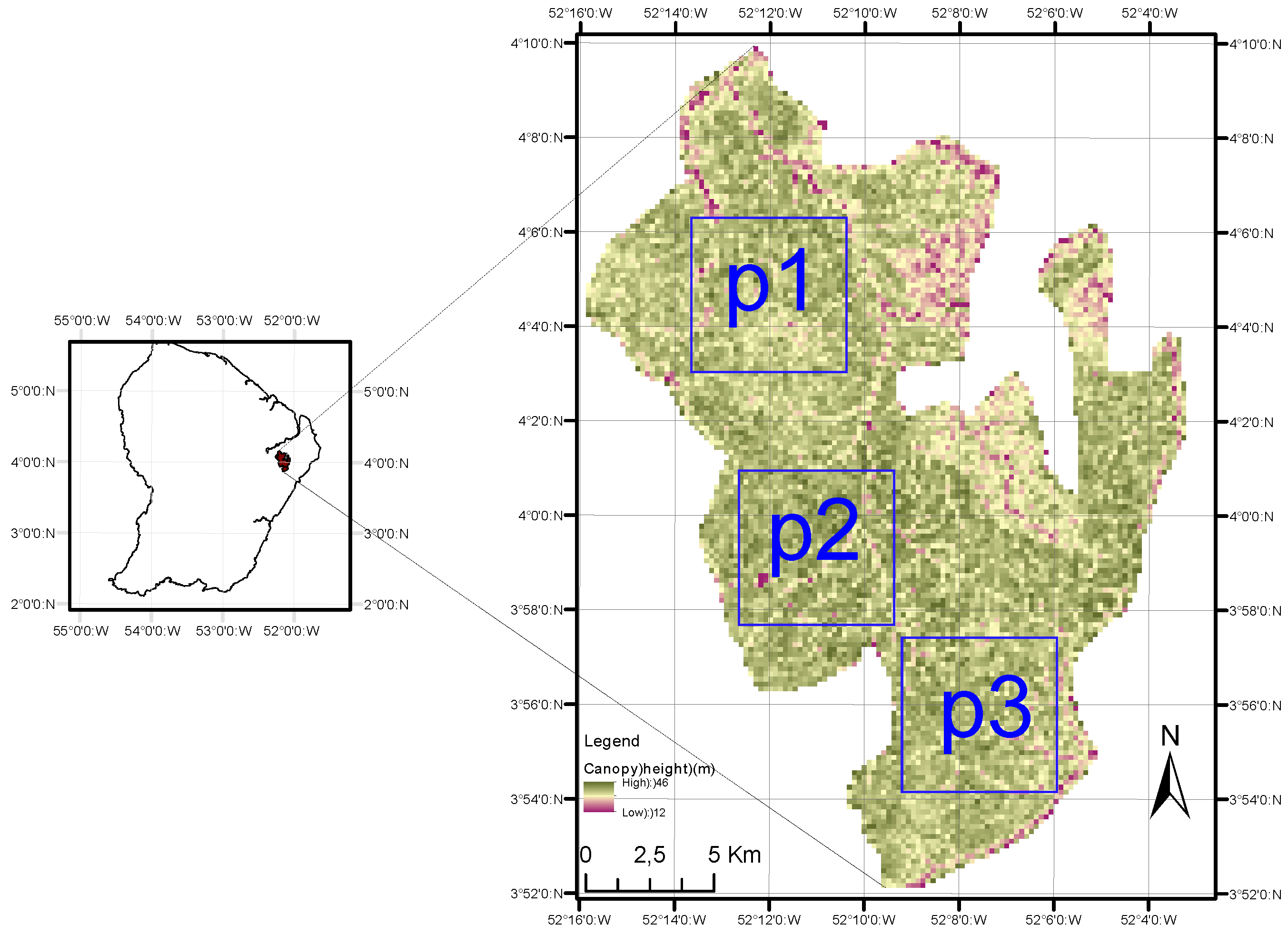
2.1. Study Site
2.2. LiDAR DEM and DCM
2.2.1. Environmental Variables
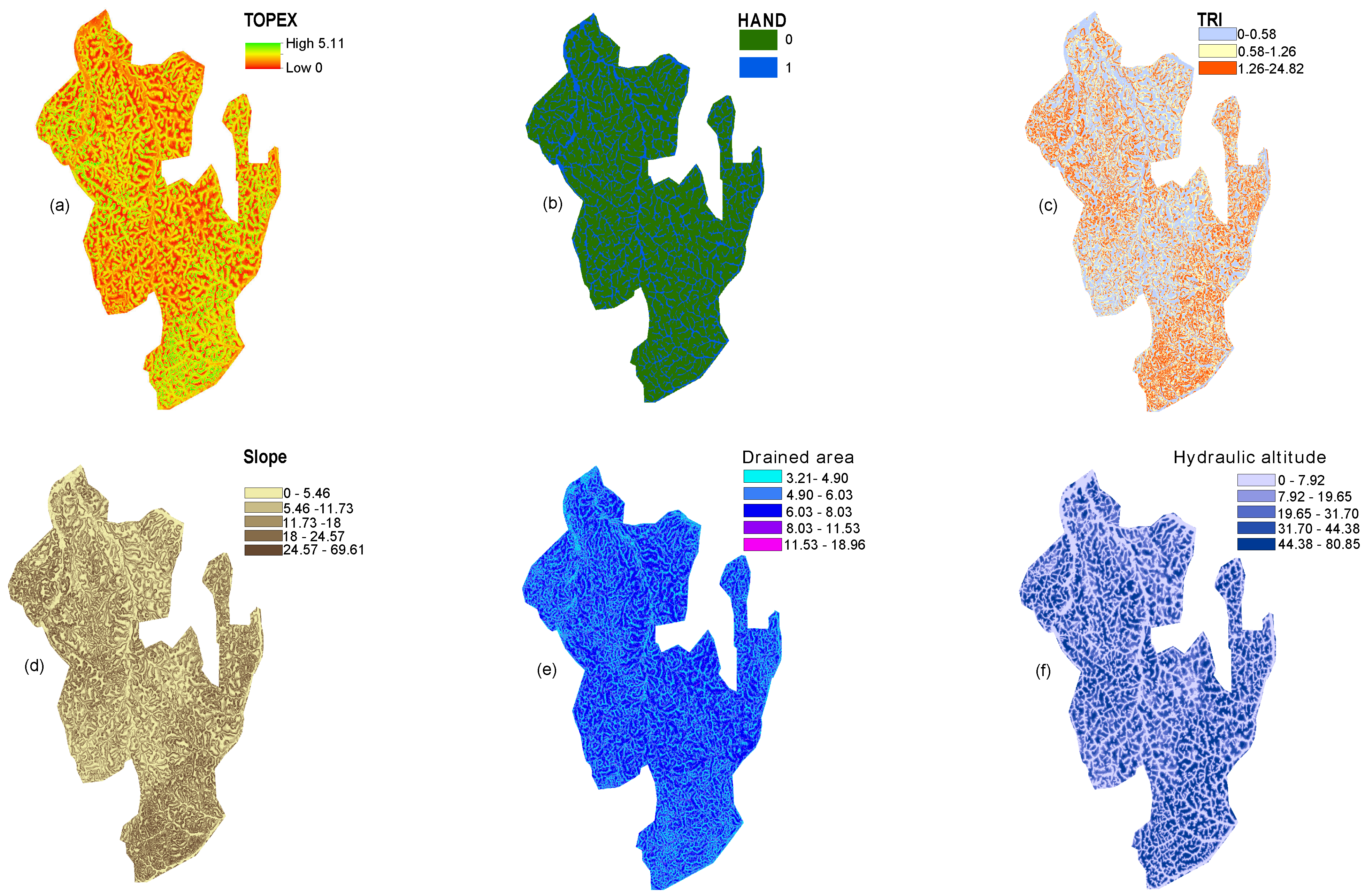
- The Drained Area (DA) was extracted. The DA measures the surface of the hydraulic basin that flows through a cell. A low value indicates cells located on the border between two basins, whereas the highest values indicate cells located downstream.
- The Hydraulic Altitude (HA) was computed from the 3rd order hydraulic system. The HA of a cell is its altitude above the closest stream of its hydraulic basin.
- In order to map terra firme versus floodplain forest, we used a predictive model adapted from the HAND topographic algorithm (Heigh Above the Nearest Drainage ) using a high resolution DTM. The HAND binary index has been developed to model soil water conditions in Amazonia rainforest and proved to provide good proxy of permanent water saturation in other contexts [38]. We classify pixels with HAND ≤ 2 m as floodplain and other pixels as terra firme.
- We computed the slope from the DEM as the maximum rate of change in elevation from a cell to its eight neighboring cells over the distance between them.
- The Terrain Ruggedness Index (TRI) catches the difference between flat and more mountainous landscapes. The TRI was calculated using Arc-GIS 10.2 as the sum of the change in altitude between a grid cell and its eight neighbors.
- We used the Topographic Exposure index (TOPEX) to measure exposure to the wind. TOPEX is a variable that represents the degree of shelter assigned to a location. TOPEX was derived from the quantitative assessment of the horizontal inclination. The values of this index are closely correlated with the wind-shaped index. Exposure is calculated based on the height and distance of the surrounding horizon, which are combined to obtain the inflection angle. We used this angle to quantify topographic exposure. A higher inflection angle is equal to a lower exposure or more shelter [23].
2.3. Data Analysis
2.3.1. Spatial Autocorrelation
2.3.2. Link between Forest Canopy Height and Environmental Drivers at Different Scales
2.3.3. Prediction of Forest Height from Environmental Drivers
- The range is the lag distance at which the sill is reached. The range is the distance where the spatial correlation vanishes and the variogram levels off.
- The sill is the semivariance at the lag distances above which there is no spatial autocorrelation.
- The nugget is the semivariance at a lag distance of zero; it represents the spatial variability below the sampling frequency.
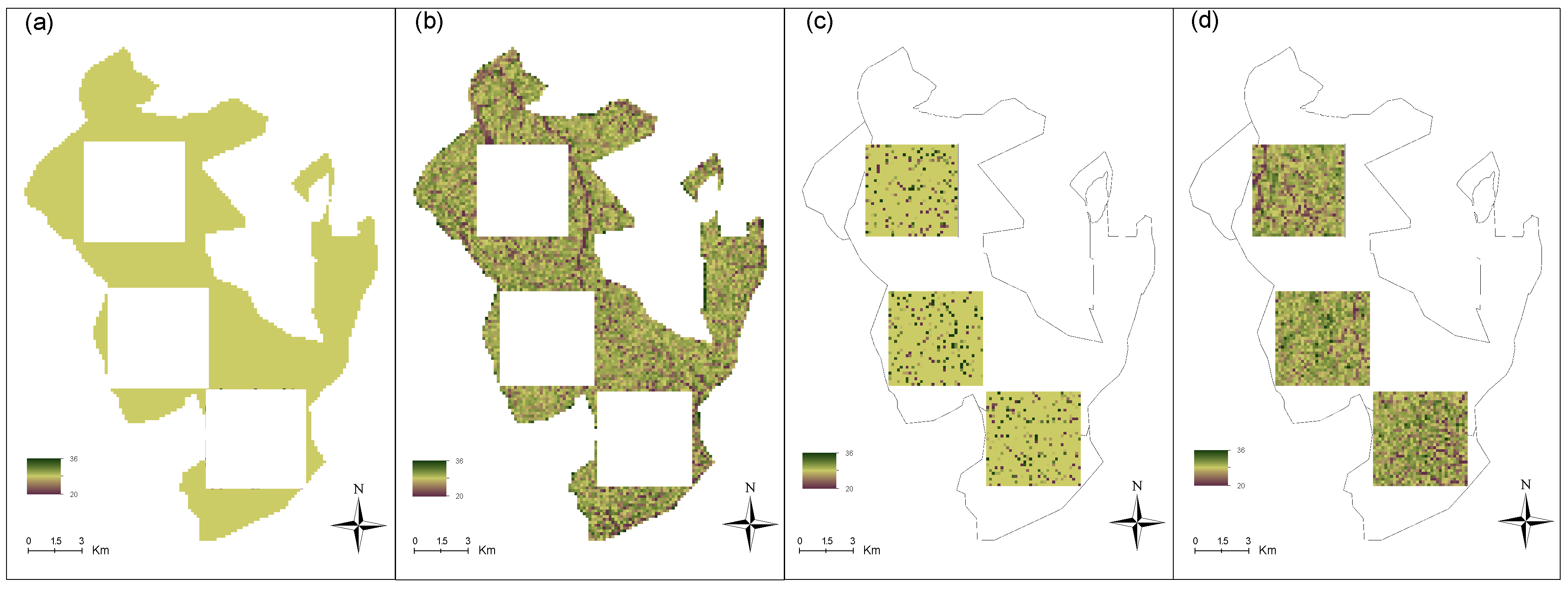
2.3.4. Strategies to build height maps
- Inside the calibration plots, where we compared the prediction with and without environmental covariates (Slope, TOPEX, TRI, HA, DA, HAND)
- Outside the calibration plots, where we compared predictions with environment-only to a null model (no calibration points, no environment)
3. Results
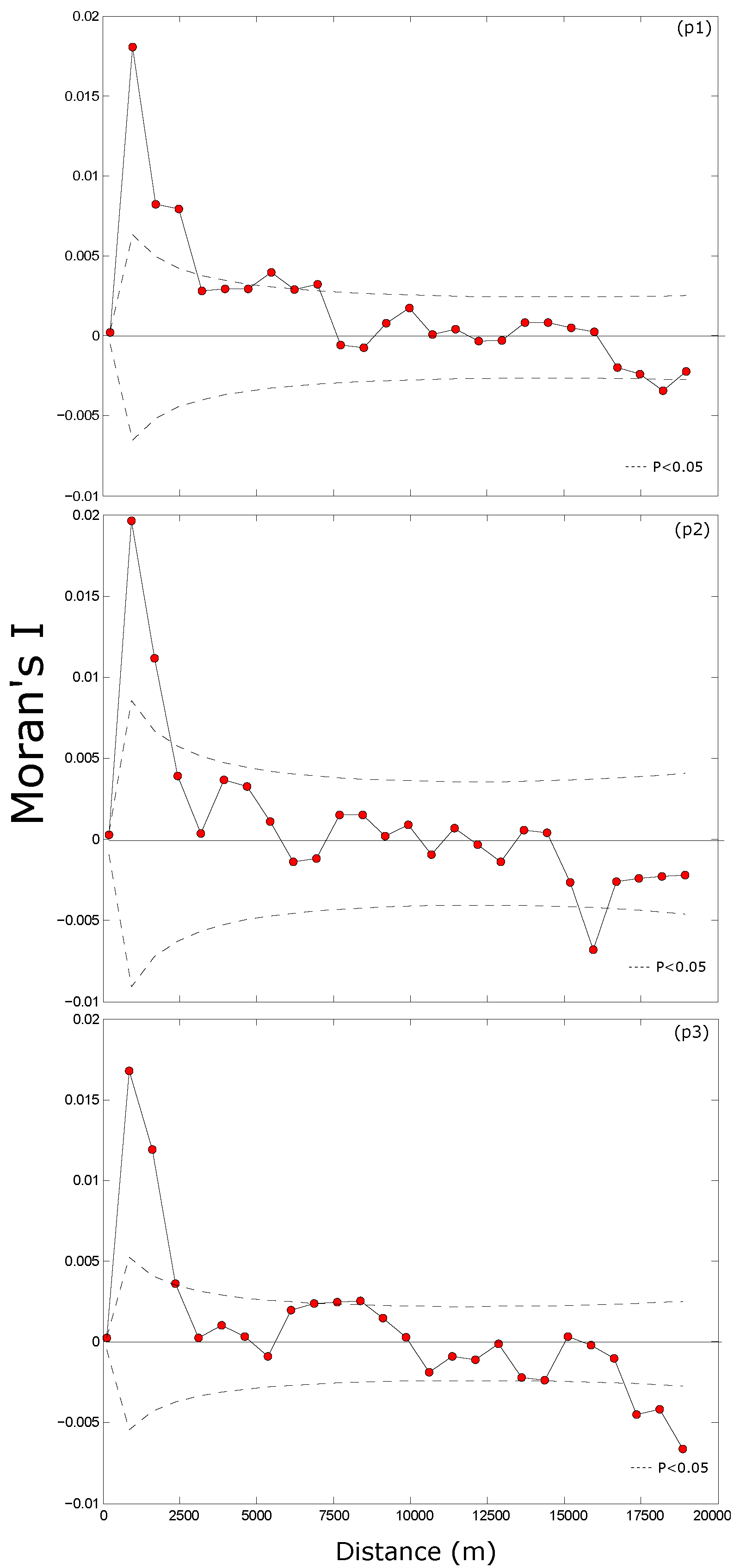
3.1. Spatial Variability of Canopy Height
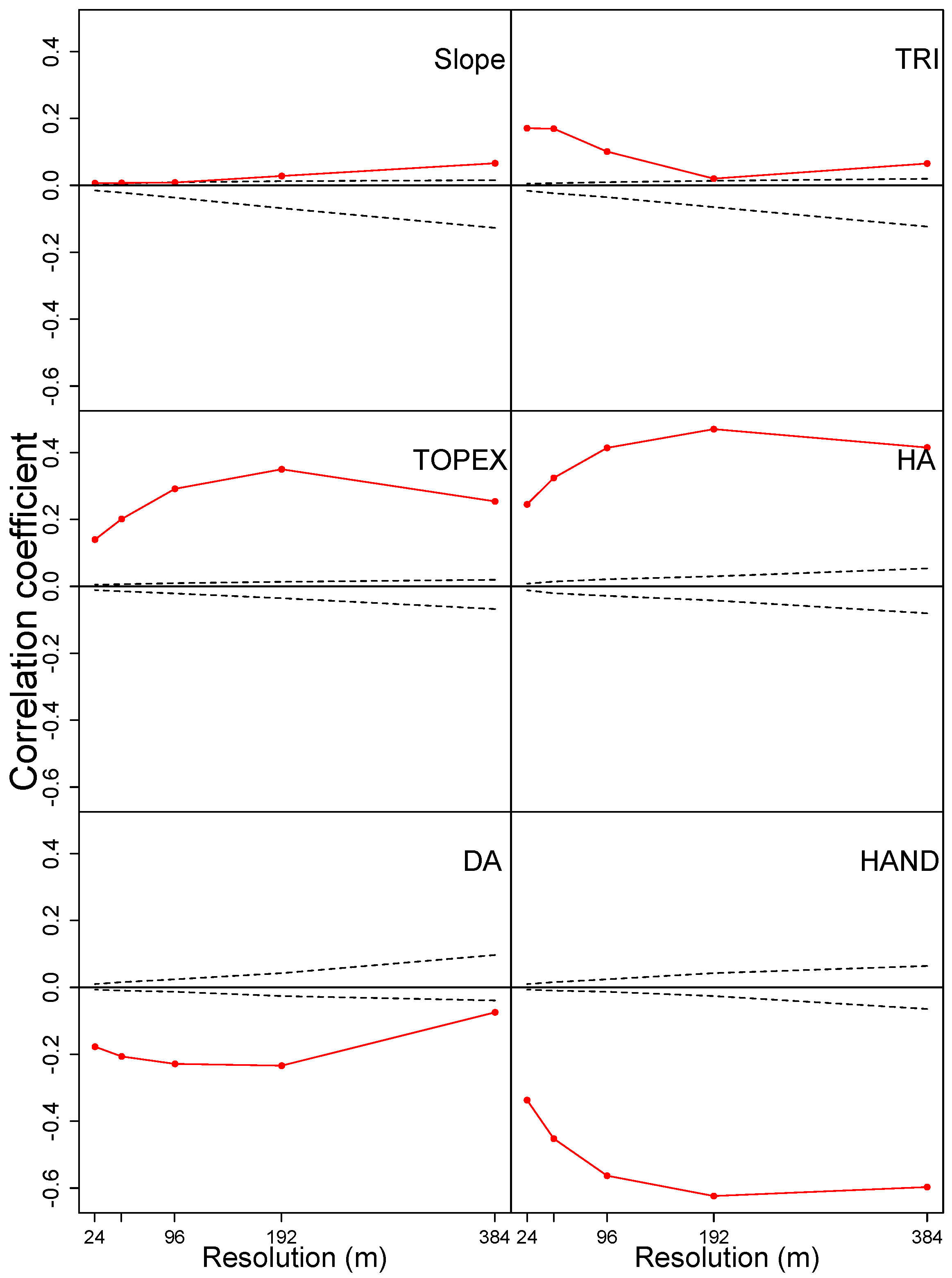
3.2. Environmental Drivers of Canopy Height
3.2.1. Effect of Hydrological Variables
3.2.2. Effect of topographical variables
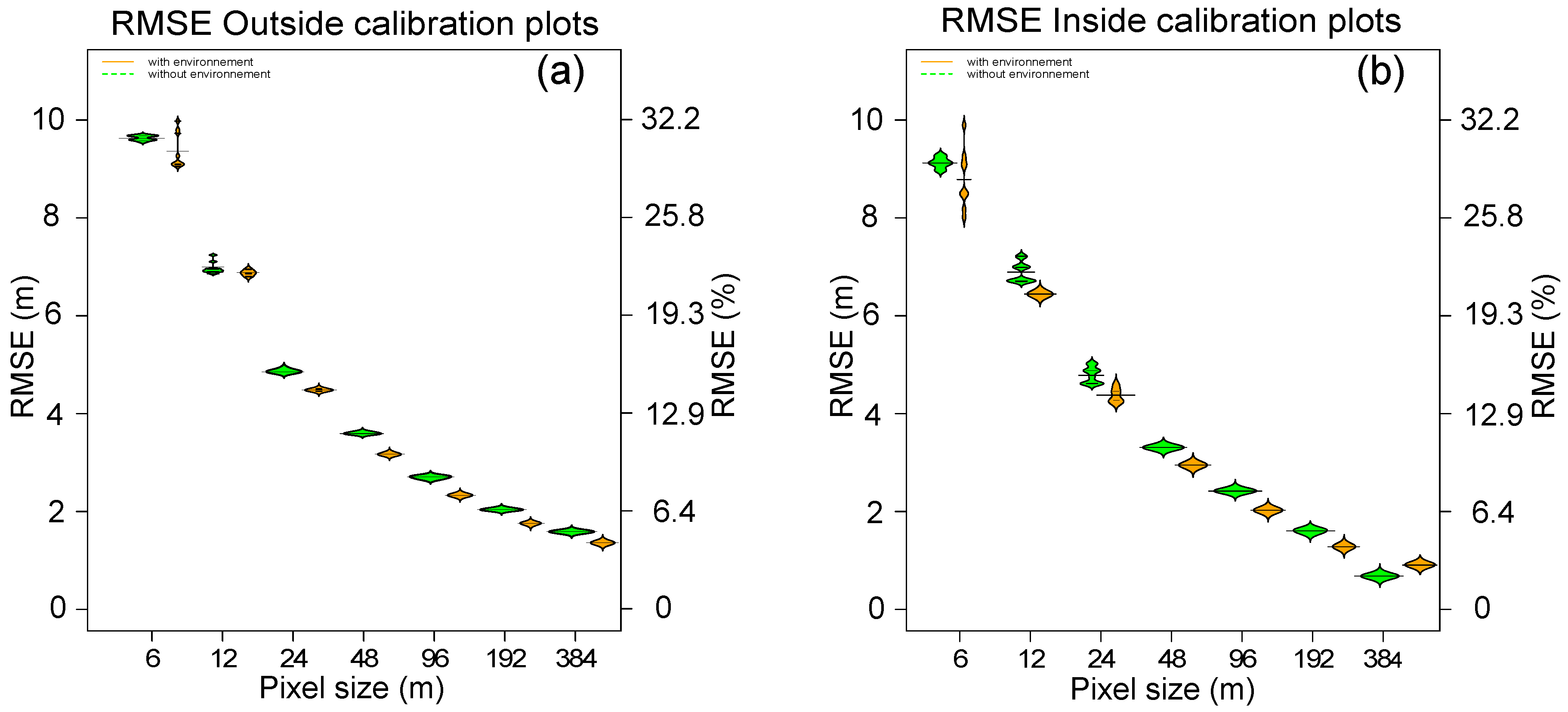
3.3. Mapping Forest Canopy Height
3.3.1. Case 1: Inside Calibration Plots
3.3.2. Case 2: Outside Calibration Plots
4. Discussion
4.1. Spatial Auto-Correlation in Forest Canopy Height
4.1.1. Spatial Structure at Fine Scale
4.1.2. Spatial Structure at Large Scale
4.2. Detecting Environmental Drivers with Multi-Scale Analysis
4.2.1. Observations from Fine to Large Scale
4.2.2. Effects of Specific Environmental Variables
4.3. Mapping Tropical Forest Canopy Height
4.3.1. Mapping Canopy Height with Low-Resolution Remote Sensing Information
4.3.2. Do Environment Variables Really Help to Map Forest Canopy Height?
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Denslow, J.S. Tropical rainforest gaps and tree species diversity. Annu. Rev. Ecol. Syst. 1987, 18, 431–451. [Google Scholar] [CrossRef]
- Clark, D.B.; Palmer, M.W.; Clark, D.A. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology 1999, 80, 2662–2675. [Google Scholar] [CrossRef]
- Andersen, H.E.; McGaughey, R.J.; Reutebuch, S.E. Estimating forest canopy fuel parameters using LIDAR data. Remote Sens. Environ. 2005, 94, 441–449. [Google Scholar] [CrossRef]
- Okuda, T.; Suzuki, M.; Adachi, N.; Quah, E.S.; Hussein, N.A.; Manokaran, N. Effect of selective logging on canopy and stand structure and tree species composition in a lowland dipterocarp forest in peninsular Malaysia. For. Ecol. Manag. 2003, 175, 297–320. [Google Scholar] [CrossRef]
- Brienen, R.; Phillips, O.; Feldpausch, T.; Gloor, E.; Baker, T.; Lloyd, J.; Lopez-Gonzalez, G.; Monteagudo-Mendoza, A.; Malhi, Y.; Lewis, S.; et al. Long-term decline of the Amazon carbon sink. Nature 2015, 519, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molto, Q.; Hérault, B.; Boreux, J.J.; Daullet, M.; Rousteau, A.; Rossi, V. Predicting tree heights for biomass estimates in tropical forests—A test from French Guiana. Biogeosciences 2014, 11, 3121–3130. [Google Scholar] [CrossRef]
- Molto, Q.; Rossi, V.; Blanc, L. Error propagation in biomass estimation in tropical forests. Methods Ecol. Evol. 2013, 4, 175–183. [Google Scholar] [CrossRef]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragão, L.E.; Anderson, L.O.; Myneni, R.B.; Nemani, R. Persistent effects of a severe drought on Amazonian forest canopy. Proc. Natl. Acad. Sci. USA 2013, 110, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping forest canopy height globally with spaceborne lidar. J. Geophys. Res. Biogeosci. 2011, 116, G04021. [Google Scholar] [CrossRef]
- Baraloto, C.; Rabaud, S.; Molto, Q.; Blanc, L.; Fortunel, C.; Herault, B.; Davila, N.; Mesones, I.; Rios, M.; Valderrama, E.; et al. Disentangling stand and environmental correlates of aboveground biomass in Amazonian forests. Glob. Chang. Biol. 2011, 17, 2677–2688. [Google Scholar] [CrossRef]
- Guitet, S.; Cornu, J.F.; Brunaux, O.; Betbeder, J.; Carozza, J.M.; Richard-Hansen, C. Landform and landscape mapping, French Guiana (South America). J. Maps 2013, 9, 325–335. [Google Scholar] [CrossRef]
- Ashton, P.S.; Hall, P. Comparisons of structure among mixed dipterocarp forests of north-western Borneo. J. Ecol. 1992, 80, 459–481. [Google Scholar] [CrossRef]
- Wagner, F.; Hérault, B.; Stahl, C.; Bonal, D.; Rossi, V. Modeling water availability for trees in tropical forests. Agric. For. Meteorol. 2011, 151, 1202–1213. [Google Scholar] [CrossRef]
- Wagner, F.; Rossi, V.; Stahl, C.; Bonal, D.; Herault, B. Water availability is the main climate driver of neotropical tree growth. PLoS ONE 2012, 7, e34074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detto, M.; Muller-Landau, H.C.; Mascaro, J.; Asner, G.P. Hydrological networks and associated topographic variation as templates for the spatial organization of tropical forest vegetation. PLoS ONE 2013, 8, e76296. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.B.; Clark, D.A. Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. Manag. 2000, 137, 185–198. [Google Scholar] [CrossRef]
- Lobo, E.; Dalling, J. Effects of topography, soil type and forest age on the frequency and size distribution of canopy gap disturbances in a tropical forest. Biogeosciences 2013, 10, 6769–6781. [Google Scholar] [CrossRef]
- Lan, G.; Hu, Y.; Cao, M.; Zhu, H. Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China. For. Ecol. Manag. 2011, 262, 1507–1513. [Google Scholar] [CrossRef]
- Ferry, B.; Morneau, F.; Bontemps, J.D.; Blanc, L.; Freycon, V. Higher treefall rates on slopes and waterlogged soils result in lower stand biomass and productivity in a tropical rain forest. J. Ecol. 2010, 98, 106–116. [Google Scholar] [CrossRef]
- Thomas, S.C.; Martin, A.R.; Mycroft, E.E. Tropical trees in a wind-exposed island ecosystem: Height-diameter allometry and size at onset of maturity. J. Ecol. 2015, 103, 594–605. [Google Scholar] [CrossRef]
- Gleason, S.M.; Williams, L.J.; Read, J.; Metcalfe, D.J.; Baker, P.J. Cyclone effects on the structure and production of a tropical upland rainforest: Implications for life-history tradeoffs. Ecosystems 2008, 11, 1277–1290. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Asner, G.P.; Morton, D.C.; Anderson, L.O.; Saatchi, S.S.; Espírito-Santo, F.D.; Palace, M.; Souza, C. Regional ecosystem structure and function: Ecological insights from remote sensing of tropical forests. Trends Ecol. Evol. 2007, 22, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Ruel, J.; Pin, D.; Spacek, L.; Cooper, K.; Benoit, R. The estimation of wind exposure for windthrow hazard rating: Comparison between Strongblow, MC2, Topex and a wind tunnel study. Forestry 1997, 70, 253–266. [Google Scholar] [CrossRef]
- Palace, M.; Sullivan, F.B.; Ducey, M.; Herrick, C. Estimating tropical forest structure using a terrestrial lidar. PLoS ONE 2016, 11, e0154115. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Mascaro, J.; Muller-Landau, H.C.; Vieilledent, G.; Vaudry, R.; Rasamoelina, M.; Hall, J.S.; van Breugel, M. A universal airborne LiDAR approach for tropical forest carbon mapping. Oecologia 2012, 168, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Lefsky, M.A.; Cohen, W.; Acker, S.; Parker, G.G.; Spies, T.; Harding, D. Lidar remote sensing of the canopy structure and biophysical properties of Douglas-fir western hemlock forests. Remote Sens. Environ. 1999, 70, 339–361. [Google Scholar] [CrossRef]
- Alonzo, M.; McFadden, J.P.; Nowak, D.J.; Roberts, D.A. Mapping urban forest structure and function using hyperspectral imagery and lidar data. Urban For. Urban Green. 2016, 17, 135–147. [Google Scholar] [CrossRef]
- Popescu, S.C. Estimating biomass of individual pine trees using airborne lidar. Biomass Bioenergy 2007, 31, 646–655. [Google Scholar] [CrossRef]
- Drake, J.B.; Dubayah, R.O.; Clark, D.B.; Knox, R.G.; Blair, J.B.; Hofton, M.A.; Chazdon, R.L.; Weishampel, J.F.; Prince, S. Estimation of tropical forest structural characteristics using large-footprint lidar. Remote Sens. Environ. 2002, 79, 305–319. [Google Scholar] [CrossRef]
- Drake, J.B.; Dubayah, R.O.; Knox, R.G.; Clark, D.B.; Blair, J.B. Sensitivity of large-footprint lidar to canopy structure and biomass in a neotropical rainforest. Remote Sens. Environ. 2002, 81, 378–392. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 045023. [Google Scholar] [CrossRef]
- Hudak, A.T.; Lefsky, M.A.; Cohen, W.B.; Berterretche, M. Integration of lidar and Landsat ETM+ data for estimating and mapping forest canopy height. Remote Sens. Environ. 2002, 82, 397–416. [Google Scholar] [CrossRef]
- Fayad, I.; Baghdadi, N.; Bailly, J.; Barbier, N.; Gond, V.; Hérault, B.; El Hajj, M.; Lochard, J.; Perrin, J. Regional scale rain-forest height mapping using regression-kriging of spaceborne and airborne LiDAR data: Application on French Guiana. In Proceedings of the 2015 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Milan, Italy, 26–31 July 2015; pp. 4109–4112.
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Cerioli, A. Testing mutual independence between two discrete-valued spatial processes: A correction to pearson chi-squared. Biometrics 2002, 58, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Deblauwe, V.; Kennel, P.; Couteron, P. Testing pairwise association between spatially autocorrelated variables: A new approach using surrogate lattice data. PLoS ONE 2012, 7, e48766. [Google Scholar] [CrossRef] [PubMed]
- Altoa 2016 Website. Altoa. Maitrisez Votre Espace. Available online: http://www.altoa.org/fr/lidar.html (accessed on 10 January 2016).
- Nobre, A.; Cuartas, L.; Hodnett, M.; Rennó, C.; Rodrigues, G.; Silveira, A.; Waterloo, M.; Saleska, S. Height above the nearest drainage—A hydrologically relevant new terrain model. J. Hydrol. 2011, 404, 13–29. [Google Scholar] [CrossRef]
- Fortin, M.J.; Drapeau, P.; Legendre, P. Spatial autocorrelation and sampling design in plant ecology. Vegetatio 1989, 83, 209–222. [Google Scholar] [CrossRef]
- Tsui, O.W.; Coops, N.C.; Wulder, M.A.; Marshall, P.L. Integrating airborne LiDAR and space-borne radar via multivariate kriging to estimate above-ground biomass. Remote Sens. Environ. 2013, 139, 340–352. [Google Scholar] [CrossRef]
- R Project Software 2015. Available online: http://www.r-project.org/ (accessed on 20 December 2015).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the Geospatial Data Abstraction Library. R Package Version 0.8-10. Available online: https://cran.r-project.org/web/packages/rgdal/index.html (accessed on 20 December 2015).
- Bivand, R.; Lewin-Koh, N. Maptools: Tools for Reading and Handling Spatial Objects. R Package Version 0.8-27. Available online: http://cran. r-project. org/web/packages/maptools/maptools (accessed on 20 December 2015).
- Vincent, G.; Sabatier, D.; Blanc, L.; Chave, J.; Weissenbacher, E.; Pélissier, R.; Fonty, E.; Molino, J.F.; Couteron, P. Accuracy of small footprint airborne LiDAR in its predictions of tropical moist forest stand structure. Remote Sens. Environ. 2012, 125, 23–33. [Google Scholar] [CrossRef]
- Asner, G.P.; Kellner, J.R.; Kennedy-Bowdoin, T.; Knapp, D.E.; Anderson, C.; Martin, R.E. Forest canopy gap distributions in the southern Peruvian Amazon. PLoS ONE 2013, 8, e60875. [Google Scholar] [CrossRef] [PubMed]
- Bruijnzeel, L.A.; Scatena, F.N.; Hamilton, L.S. Tropical Montane Cloud Forests: Science for Conservation and Management; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Lefsky, M.A. A global forest canopy height map from the moderate resolution imaging spectroradiometer and the geoscience laser altimeter system. Geophys. Res. Lett. 2010, 37, 15. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Sabatier, D.; Grimaldi, M.; Prévost, M.F.; Guillaume, J.; Godron, M.; Dosso, M.; Curmi, P. The influence of soil cover organization on the floristic and structural heterogeneity of a Guianan rain forest. Plant Ecol. 1997, 131, 81–108. [Google Scholar] [CrossRef]
- Paget, D. Etude de la Diversite Spatiale des Ecosystemes Forestiers Guyanais: Reflexion Methodologique et Application. Ph.D. Thesis, Ecole Nationale de Génie Rural des Eaux et Forêts (ENGREF), Nancy, France, 1999. [Google Scholar]
- Sabatier, D. Diversité des arbres et du peuplement forestier en Guyane. In Gestion de l’écosysteme Forestier et Aménagement de L’espace Régional; Actes du II éme Congrés Régional de L’Evironnement: Sapanguy, Cayenne, 1990; pp. 41–47. [Google Scholar]
- Lescure, J.P.; Boulet, R. Relationships between soil and vegetation in a tropical rain forest in French Guiana. Biotropica 1985, 17, 155–164. [Google Scholar] [CrossRef]
- Hilbert, C.; Schmullius, C. Influence of surface topography on ICESat/GLAS forest height estimation and waveform shape. Remote Sens. 2012, 4, 2210–2235. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variable | Abbreviation | Range | Unit |
|---|---|---|---|
| Slope | Slope | (0.1∘69.9) | |
| Terrain Ruggedness Index | TRI | (0.01–5.2) | |
| Topographic Exposure | TOPEX | (1–4.9) | |
| Drained Area | DA | (3.2–18.4) | |
| Hydraulic Altitude | HA | (0–78.6) | m |
| Height Above the Nearest Drainage | HAND | (0–1) | m |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goulamoussène, Y.; Bedeau, C.; Descroix, L.; Deblauwe, V.; Linguet, L.; Hérault, B. Weak Environmental Controls of Tropical Forest Canopy Height in the Guiana Shield. Remote Sens. 2016, 8, 747. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8090747
Goulamoussène Y, Bedeau C, Descroix L, Deblauwe V, Linguet L, Hérault B. Weak Environmental Controls of Tropical Forest Canopy Height in the Guiana Shield. Remote Sensing. 2016; 8(9):747. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8090747
Chicago/Turabian StyleGoulamoussène, Youven, Caroline Bedeau, Laurent Descroix, Vincent Deblauwe, Laurent Linguet, and Bruno Hérault. 2016. "Weak Environmental Controls of Tropical Forest Canopy Height in the Guiana Shield" Remote Sensing 8, no. 9: 747. https://0-doi-org.brum.beds.ac.uk/10.3390/rs8090747