Early-Life Iron Deficiency Reduces Brain Iron Content and Alters Brain Tissue Composition Despite Iron Repletion: A Neuroimaging Assessment



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Housing
2.2. Dietary Treatments
2.3. Magnetic Resonance Imaging
2.3.1. Structural MRI Acquisition & Analysis
2.3.2. Quantitative Susceptibility Mapping
2.3.3. Tract-Based Spatial Statistics
2.4. Statistical Analysis
2.4.1. Voxel-Based Morphometry Statistics
2.4.2. Tract-Based Spatial Statistics
3. Results
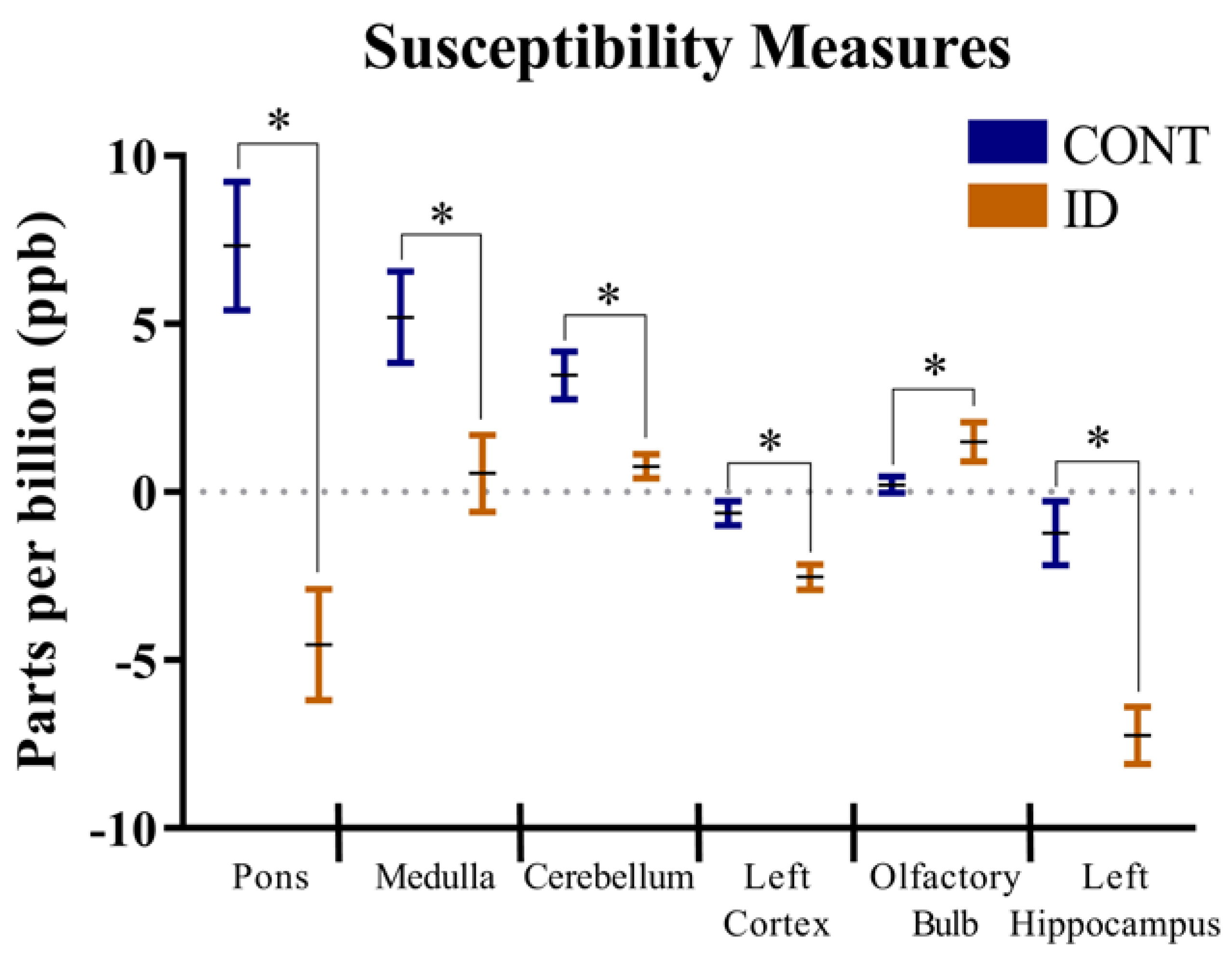
3.1. Quantitative Susceptibility Mapping
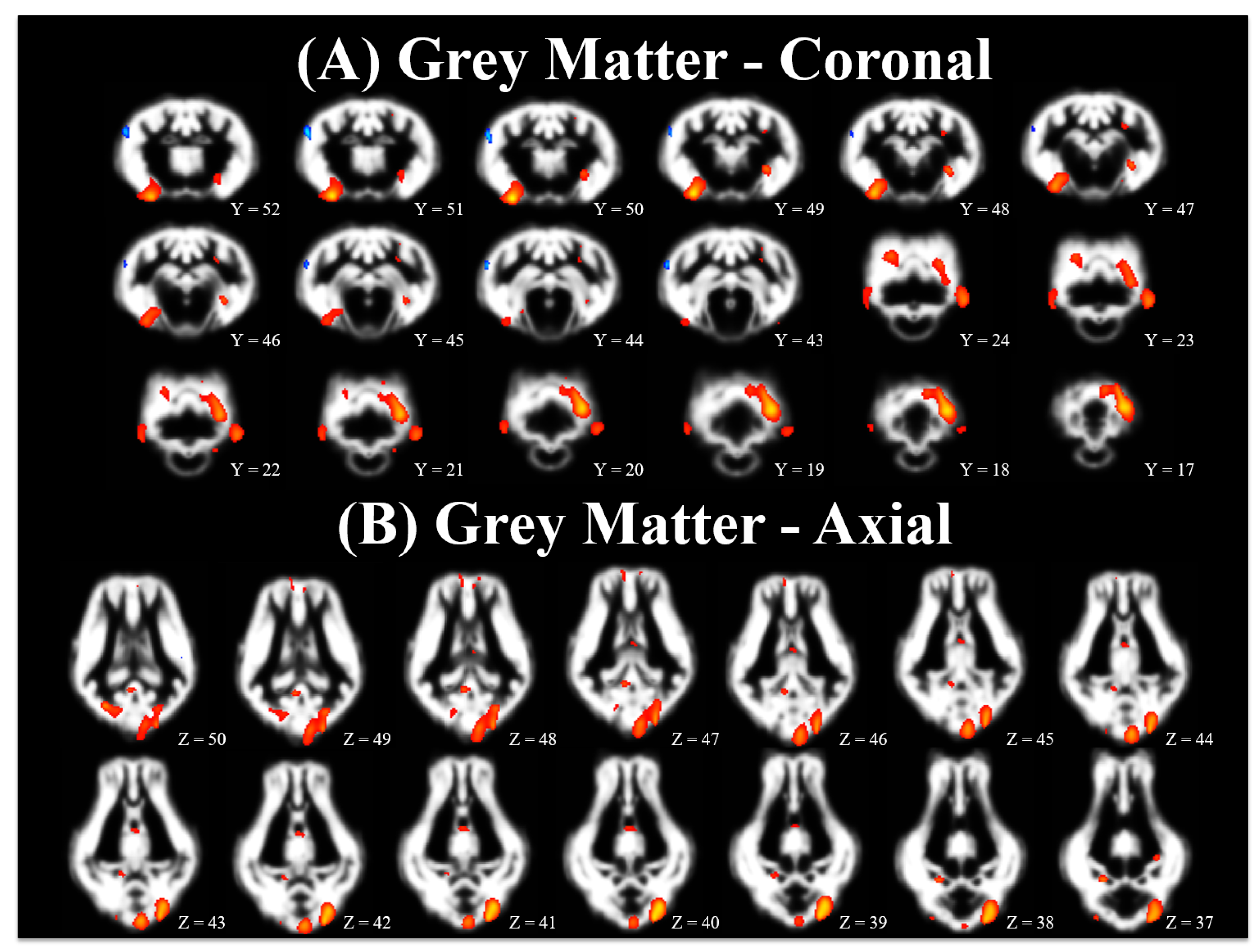
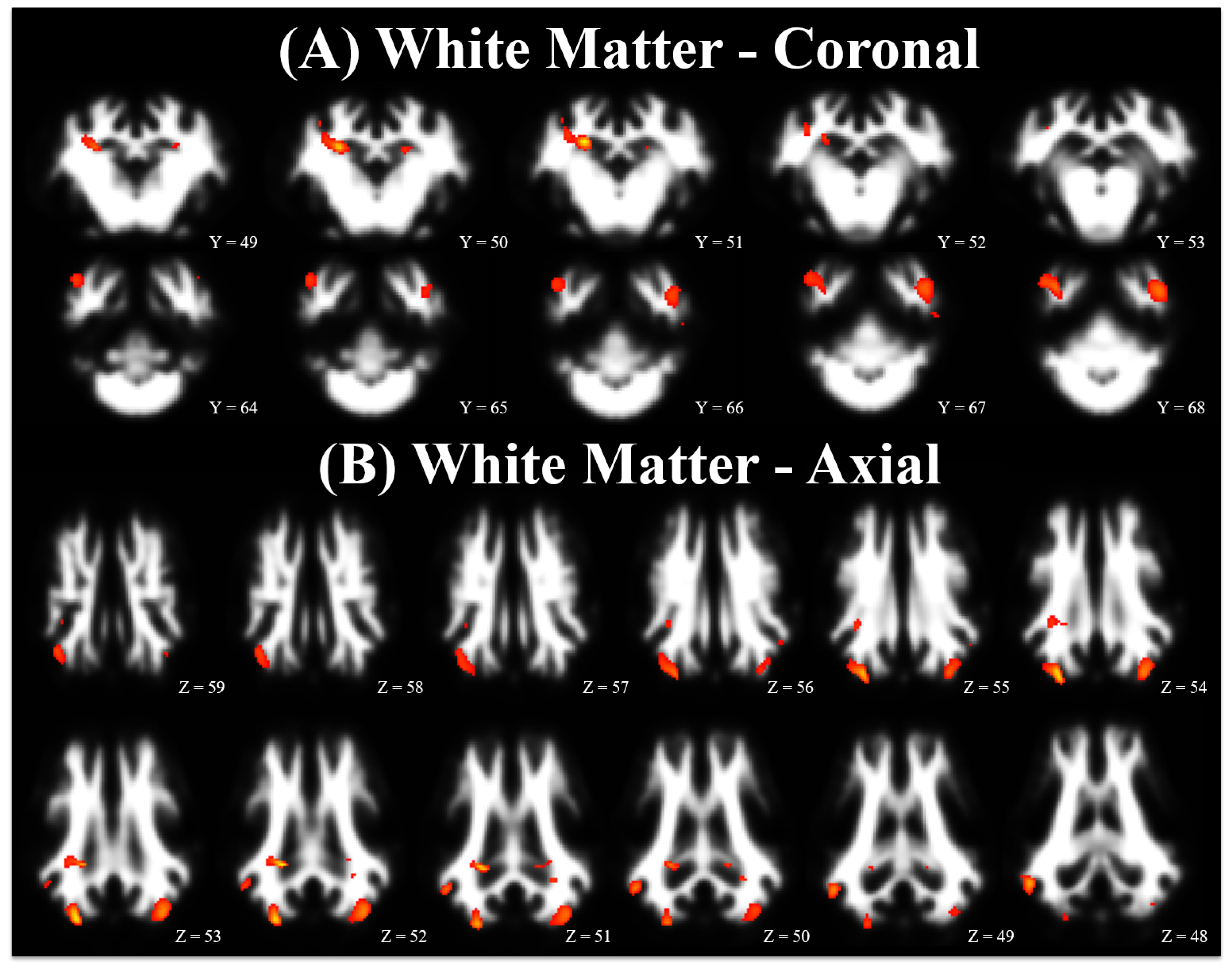
3.2. Voxel-Based Morphometry
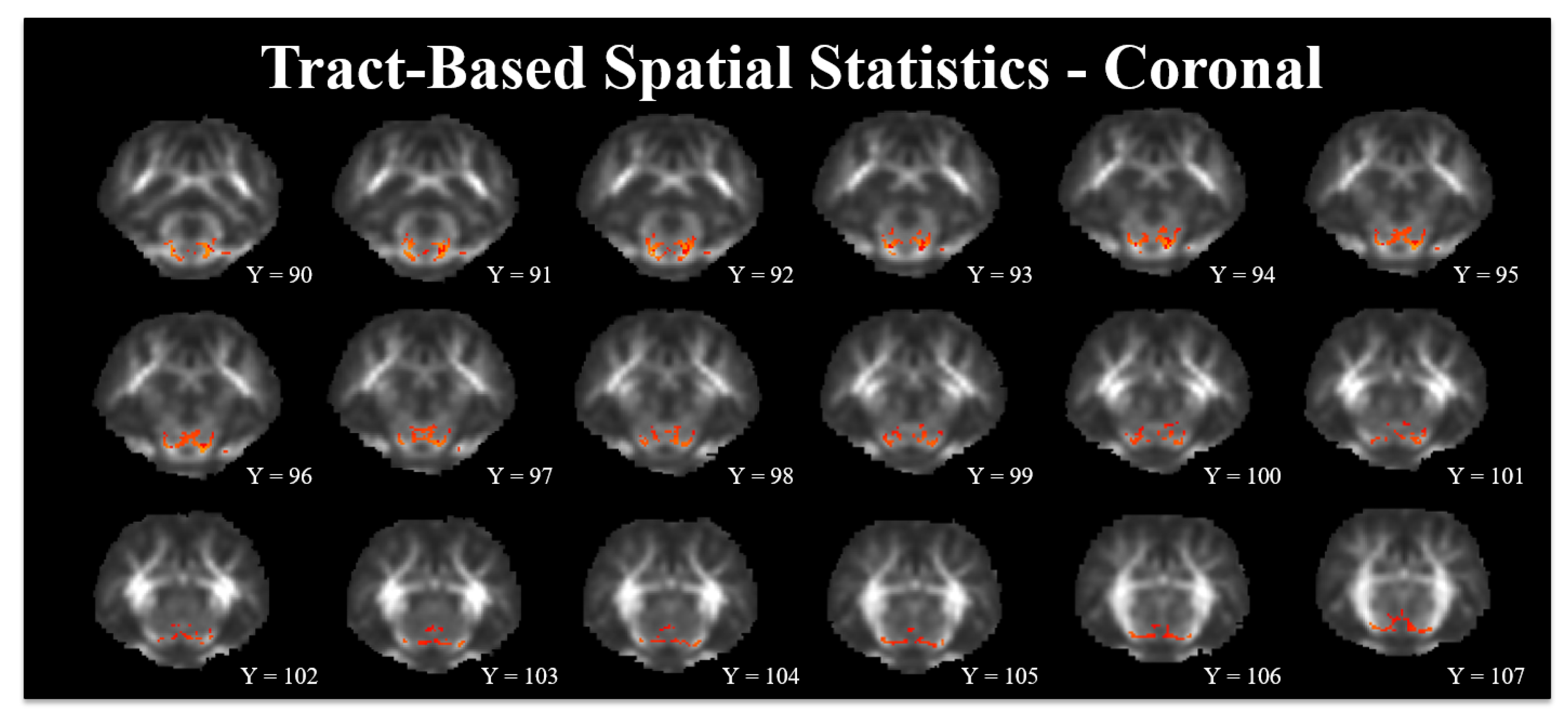
3.3. Tract-Based Spatial Statistics
4. Discussion
4.1. Quantitative Susceptibility Measures
4.2. Voxel-Based Morphometry
4.3. Tract-Based Spatial Statistics
4.4. Limitations
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Micronutrient Deficiencies—Iron Deficiency Anemia; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Iron Deficiency Anaemia: Assessment, Prevention, and Control: A Guide for Programme Managers; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; de Benoist, B. Worldwide prevalence of anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993–2005. Public Health Nutr. 2009, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Georgieff, M.K. Iron deficiency and brain development. Semin. Pediatr. Neurol. 2006, 13, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Iron assessment to protect the developing brain. Am. J. Clin. Nutr. 2017, 106, 1588S–1593S. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Nutrition and the developing brain: Nutrient priorities and measurement. Am. J. Clin. Nutr. 2007, 85, 614–620. [Google Scholar]
- Radlowski, E.C.; Johnson, R.W. Perinatal iron deficiency and neurocognitive development. Front. Hum. Neurosci. 2013, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shafir, T.; Angulo-Barroso, R.; Jing, Y.; Angelilli, M.L.; Jacobson, S.W.; Lozoff, B. Iron deficiency and infant motor development. Early Hum. Dev. 2008, 84, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Congdon, E.L.; Westerlund, A.; Algarin, C.R.; Peirano, P.D.; Gregas, M.; Lozoff, B.; Nelson, C.A. Iron deficiency in infancy is associated with altered neural correlates of recognition memory at 10 years. J. Pediatr. 2012, 160, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, A.F.; Koss, M.; Burden, M.J.; Jonides, J.; Nelson, C.A.; Kaciroti, N.; Jimenez, E.; Lozoff, B. Iron deficiency in infancy and neurocognitive functioning at 19 years: Evidence of long-term deficits in executive function and recognition memory. Nutr. Neurosci. 2010, 13, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Smith, J.B.; Kaciroti, N.; Clark, K.M.; Guevara, S.; Jimenez, E. Functional significance of early-life iron deficiency: Outcomes at 25 years. J. Pediatr. 2013, 163, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Algarín, C.; Peirano, P.; Garrido, M.; Pizarro, F.; Lozoff, B. Iron deficiency anemia in infancy: Long-lasting effects on auditory and visual system functioning. Pediatr. Res. 2003, 53, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Otero, G.A.; Fernández, T.; Pliego-Rivero, F.B.; Mendieta, G.G. Iron therapy substantially restores qEEG maturational lag among iron-deficient anemic infants. Nutr. Neurosci. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bastian, T.W.; Santarriaga, S.; Nguyen, T.A.; Prohaska, J.R.; Georgieff, M.K.; Anderson, G.W. Fetal and neonatal iron deficiency but not copper deficiency increases vascular complexity in the developing rat brain. Nutr. Neurosci. 2015, 18, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Todorich, B.; Pasquini, J.M.; Garcia, C.I.; Paez, P.M.; Connor, J.R. Oligodendrocytes and myelination: The role of iron. Glia 2009, 57, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Unger, E.; Connor, J.; Felt, B.; Georgieff, M. Early iron deficiency has brain and behavior effects consistent with dopaminergic function. J. Nutr. 2011, 141, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Felt, B.T.; Beard, J.L.; Schallert, T.; Shao, J.; Aldridge, J.W.; Connor, J.R.; Georgieff, M.K.; Lozoff, B. Persistent neurochemical and behavioral abnormalities in adulthood despite early iron supplementation for perinatal iron deficiency anemia in rats. Behav. Brain Res. 2006, 171, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Bastian, T.W.; Von Hohenberg, W.C.; Mickelson, D.J.; Lanier, L.M.; Georgieff, M.K. Iron deficiency impairs developing hippocampal neuron gene expression, energy metabolism, and dendrite complexity. Dev. Neurosci. 2016, 38, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Greminger, A.R.; Lee, D.L.; Shrager, P.; Mayer-Proschel, M. Gestational iron deficiency differentially alters the structure and function of white and gray matter brain regions of developing rats. J. Nutr. 2014, 144, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Monk, C.; Georgieff, M.K.; Xu, D.; Hao, X.; Bansal, R.; Gustafsson, H.; Spicer, J.; Peterson, B.S. Maternal prenatal iron status and tissue organization in the neonatal brain. Pediatr. Res. 2016, 79, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Allen, R.P.; Earley, C.J.; Liu, H.; Cruz, T.E.; Edden, R.A.E.; Barker, P.B.; van Zijl, P.C.M. Brain iron deficiency in idiopathic restless legs syndrome measured by quantitative magnetic susceptibility at 7 tesla. Sleep Med. 2016, 22, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Chan, G.C.-F.; Chu, J.; Chan, Q.; Ha, S.-Y.; Moseley, M.E.; Khong, P.-L. MR quantitative susceptibility imaging for the evaluation of iron loading in the brains of patients with β-thalassemia. Am. J. Neuroradiol. 2014, 35, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.L.H.; Li, W.; Wei, H.; Wu, B.; Xiao, X.; Liu, C.; Worley, G.; Egger, H.L. Magnetic susceptibility of brain iron is associated with childhood spatial IQ. Neuroimage 2016, 132, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Mudd, A.; Fil, J.; Knight, L.; Dilger, R. Dietary iron repletion following early-life dietary iron deficiency does not correct regional volumetric or diffusion tensor changes in the developing pig brain. Front. Neurol. 2017. [Google Scholar] [CrossRef]
- Gan, L.; Yang, B.; Mei, H. The effect of iron dextran on the transcriptome of pig hippocampus. Genes Genom. 2017, 39, 1–14. [Google Scholar] [CrossRef]
- Antonides, A.; Schoonderwoerd, A.C.; Scholz, G.; Berg, B.M.; Nordquist, R.E.; van der Staay, F.J. Pre-weaning dietary iron deficiency impairs spatial learning and memory in the cognitive holeboard task in piglets. Front. Behav. Neurosci. 2015, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (USA). Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Mudd, A.T.; Getty, C.; Sutton, B.; Dilger, R. Perinatal choline deficiency delays brain development and alters metabolite concentrations in the young pig. Nutr. Neurosci. 2016, 19, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.S.; Sutton, B.P.; Dilger, R.N.; Johnson, R. An in vivo three-dimensional magnetic resonance imaging-based averaged brain collection of the neonatal piglet (Sus scrofa). PLoS ONE 2014, 9, 107650. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.; Liang, Z.-P. A subspace approach to high-resolution spectroscopic imaging. Magn. Reson. Med. 2014, 71, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Lam, F.; Li, Y.; Clifford, B.; Liang, Z.-P. Simultaneous QSM and metabolic imaging of the brain using SPICE. Magn. Reson. Med. 2018, 79, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, T. Quantitative susceptibility mapping (QSM): Decoding MRI data for a tissue magnetic biomarker. Magn. Reson. Med. 2015, 73, 82–101. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Liu, T.; Spincemaille, P.; Wang, Y. Background field removal by solving the Laplacian boundary value problem. NMR Biomed. 2014, 27, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.J.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004, 23, S208–S219. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Jenkinson, M.; Johansen-Berg, H.; Rueckert, D.; Nichols, T.E.; Mackay, C.E.; Watkins, K.E.; Ciccarelli, O.; Cader, M.Z.; Matthews, P.M.; et al. Tract-based spatial statistics: Voxelwise analysis of multi-subject diffusion data. Neuroimage 2006, 31, 1487–1505. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Long-term brain and behavioral consequences of early iron deficiency. Nutr. Rev. 2011, 69, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Haacke, E.M.; Cheng, N.Y.C.; House, M.J.; Liu, Q.; Neelavalli, J.; Ogg, R.J.; Khan, A.; Ayaz, M.; Kirsch, W.; Obenaus, A. Imaging iron stores in the brain using magnetic resonance imaging. Magn. Reson. Imaging 2005, 23, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, M.W. Transport of iron in the blood-brain-cerebrospinal fluid system. J. Neurochem. 1997, 69, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Rytych, J.L.; Elmore, M.R.P.; Burton, M.D.; Conrad, M.S.; Donovan, S.M.; Dilger, R.N.; Johnson, R.W. Early life iron deficiency impairs spatial cognition in neonatal piglets. J. Nutr. 2012, 142, 2050–2056. [Google Scholar] [CrossRef] [PubMed]
- Jorgenson, L.A.; Wobken, J.D.; Georgieff, M.K. Perinatal iron deficiency alters apical dendritic growth in hippocampal CA1 pyramidal neurons. Dev. Neurosci. 2003, 25, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Schachtschneider, K.M.; Liu, Y.; Rund, L.A.; Madsen, O.; Johnson, R.W.; Groenen, M.A.M.; Schook, L.B. Impact of neonatal iron deficiency on hippocampal DNA methylation and gene transcription in a porcine biomedical model of cognitive development. BMC Genom. 2016, 17, 856. [Google Scholar] [CrossRef] [PubMed]
- Drobyshevsky, A.; Song, S.; Gamkrelidze, G.; Wyrwicz, A.M.; Derrick, M.; Meng, F.; Li, L.; Ji, X.; Trommer, B.; Beardsley, D.J.; et al. Developmental changes in diffusion anisotropy coincide with immature oligodendrocyte progression and maturation of compound action potential. J. Neurosci. 2005, 25, 5988–5997. [Google Scholar] [CrossRef] [PubMed]
- Hermoye, L.; Saint-Martin, C.; Cosnard, G.; Lee, S.-K.; Kim, J.; Nassogne, M.-C.; Menten, R.; Clapuyt, P.; Donohue, P.K.; Hua, K.; et al. Pediatric diffusion tensor imaging: Normal database and observation of the white matter maturation in early childhood. Neuroimage 2006, 29, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-L.; Zhang, L.; Shao, J.; Qin, Y.-F.; Yang, R.-W.; Zhao, Z.-Y. Effect of perinatal iron deficiency on myelination and associated behaviors in rat pups. Behav. Brain Res. 2008, 188, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.S.; Steinkirchner, T.M.; Rao, G.A.; Larkin, E.C. Effect of prenatal iron deficiency on myelination in rat pups. Am. J. Pathol. 1986, 125, 620–624. [Google Scholar] [PubMed]
- Xu, X.; Wang, Q.; Zhang, M. Age, gender, and hemispheric differences in iron deposition in the human brain: An in vivo MRI study. Neuroimage 2008, 40, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Leyshon, B.J.; Radlowski, E.C.; Mudd, A.T.; Steelman, A.J.; Johnson, R.W. Postnatal iron deficiency impairs brain development in piglets. J. Nutr. 2016, 146, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Nelissen, E.; De Vry, J.; Antonides, A.; Paes, D.; Schepers, M.; van der Staay, F.J.; Prickaerts, J.; Vanmierlo, T. Early-postnatal iron deficiency impacts plasticity in the dorsal and ventral hippocampus in piglets. Int. J. Dev. Neurosci. 2017, 59, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.S.; Dilger, R.N.; Johnson, R.W. Brain growth of the domestic pig (Sus scrofa) from 2 to 24 weeks of age: A longitudinal MRI study. Dev. Neurosci. 2012, 34, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.; Mirmiran, M.; Butler, E.E.; Lin, C.Y.; Barnes, P.D.; Kermoian, R.; Stevenson, D.K. Neonatal microstructural development of the internal capsule on diffusion tensor imaging correlates with severity of gait and motor deficits. Dev. Med. Child Neurol. 2007, 49, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Hartmann-von Monakow, K.; Akert, K.; Künzle, H. Projections of the precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey. Exp. Brain Res. 1978, 33, 395–403. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Title | CONT | ID | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Region of interest | PND 32 | PND 61 | PND 32 | PND 61 | SEM | Diet | Day | Diet × Day |
| Caudate | −6.3 | −3.4 | −2.1 | −2.4 | 1.51 | 0.103 | 0.318 | 0.217 |
| Cerebellum | 4.6 | 1.6 | 0.9 | 0.5 | 0.91 | 0.005 | 0.069 | 0.144 |
| Cerebral Aqueduct | −9.0 | −15.8 | −13.3 | −9.2 | 3.14 | 0.723 | 0.609 | 0.065 |
| Corpus Callosum | 0.6 | 5.3 | −0.1 | 4.5 | 2.04 | 0.676 | 0.029 | 0.994 |
| Fourth Ventricle | −1.1 | −9.8 | −3.6 | −7.5 | 3.59 | 0.985 | 0.057 | 0.428 |
| Hypothalamus | −4.7 | −11.0 | −2.6 | −6.3 | 2.48 | 0.161 | 0.044 | 0.550 |
| Internal Capsule | −8.6 | −8.0 | −9.5 | −7.5 | 1.04 | 0.760 | 0.303 | 0.560 |
| Lateral Ventricle | −3.8 | −1.3 | −4.3 | −2.4 | 1.48 | 0.542 | 0.110 | 0.834 |
| Left Cortex | −0.6 | −0.5 | −2.2 | −2.8 | 0.60 | 0.004 | 0.614 | 0.470 |
| Left Hippocampus | −0.5 | −1.9 | −7.6 | −6.9 | 1.53 | <0.001 | 0.811 | 0.447 |
| Medulla | 5.1 | 5.3 | −0.2 | 1.5 | 2.09 | 0.018 | 0.666 | 0.729 |
| Midbrain | −6.5 | −6.0 | −8.8 | −7.8 | 2.15 | 0.351 | 0.684 | 0.867 |
| Olfactory Bulb | 0.4 | −0.2 | 1.6 | 1.5 | 0.77 | 0.043 | 0.601 | 0.715 |
| Pons | 4.7 | 10.9 | −6.5 | −2.2 | 2.83 | <0.001 | 0.064 | 0.702 |
| Putamen-Globus Pallidus | −6.3 | −3.9 | −5.1 | −4.5 | 1.22 | 0.786 | 0.175 | 0.392 |
| Right Cortex | −1.0 | 0.0 | −2.7 | −0.5 | 0.72 | 0.083 | 0.033 | 0.383 |
| Right Hippocampus | −3.2 | −0.6 | −7.0 | −4.2 | 2.29 | 0.141 | 0.127 | 0.979 |
| Thalamus | −7.7 | −11.0 | −7.0 | −7.6 | 1.88 | 0.322 | 0.166 | 0.319 |
| Third Ventricle | −4.1 | −4.8 | −1.8 | 0.9 | 2.49 | 0.105 | 0.663 | 0.433 |
| Cluster Level | Peak Level | Local Maxima Coordinates 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Tissue | Comparison | Anatomic Region 2 | Number of Voxels | p-Value | p-Value | Pseudo-t | X | Y | Z |
| Grey | CONT > ID | Right Cortex | 2045 | <0.001 | <0.001 | 7.31 | 14.0 | 0.0 | −12.6 |
| Right Cortex | <0.001 | 5.32 | 18.9 | −13.3 | −8.4 | ||||
| Cerebellum | 2772 | <0.001 | <0.001 | 6.92 | −10.5 | −25.2 | 0.0 | ||
| Cerebellum | <0.001 | 5.98 | −2.8 | −27.3 | 2.8 | ||||
| Left Cortex | <0.001 | 5.16 | −10.5 | −16.8 | 5.6 | ||||
| Right Cortex | 443 | 0.004 | <0.001 | 5.98 | 11.9 | −15.4 | 7.7 | ||
| Cerebellum | 997 | <0.001 | <0.001 | 5.55 | −18.9 | −18.9 | −7.7 | ||
| Cerebellum | 251 | 0.023 | <0.001 | 5.54 | 9.1 | −9.8 | −2.1 | ||
| Right Hippocampus | <0.001 | 5.12 | 2.1 | −9.1 | 6.3 | ||||
| Left Cortex | 242 | 0.025 | <0.001 | 5.27 | −14.0 | −1.4 | −3.5 | ||
| Left Cortex | 204 | 0.037 | <0.001 | 5.01 | −13.3 | −7.0 | 13.3 | ||
| Cerebellum | 47 | 0.291 | <0.001 | 4.83 | 11.9 | −25.9 | −2.8 | ||
| Thalamus | 160 | 0.061 | <0.001 | 4.36 | 0.0 | 10.5 | 0.7 | ||
| Right Cortex | 79 | 0.175 | <0.001 | 4.35 | 3.5 | 36.4 | 4.9 | ||
| Right Cortex | 72 | 0.194 | <0.001 | 4.15 | 13.3 | −9.8 | 11.2 | ||
| Left Cortex | 24 | 0.455 | 0.001 | 3.97 | −5.6 | −20.3 | 11.9 | ||
| Left Cortex | 42 | 0.318 | 0.001 | 3.97 | −2.1 | 33.6 | 5.6 | ||
| Caudate | 21 | 0.486 | 0.001 | 3.81 | 2.1 | 18.2 | 2.8 | ||
| Grey | CONT <ID | Right Cortex | 419 | 0.005 | <0.001 | 5.51 | 22.4 | 0.7 | 10.5 |
| Left Cortex | 57 | 0.246 | <0.001 | 4.45 | −19.6 | 4.2 | 7.0 | ||
| Right Cortex | 35 | 0.363 | <0.001 | 4.34 | 18.2 | 16.1 | 12.6 | ||
| White | CONT > ID | Right Hippocampus | 234 | 0.022 | <0.001 | 6.37 | 8.4 | −2.1 | 8.4 |
| Right Cortex | 629 | 0.001 | <0.001 | 6.19 | 10.5 | −16.8 | 9.1 | ||
| Right Cortex | <0.001 | 4.61 | 15.4 | −9.8 | 13.3 | ||||
| Right Cortex | 378 | 0.005 | <0.001 | 5.85 | 20.3 | −7.0 | 4.2 | ||
| Left Cortex | 653 | 0.001 | <0.001 | 5.26 | −14.0 | −15.4 | 9.1 | ||
| Cerebellum | <0.001 | 4.26 | −18.9 | −16.1 | 0.7 | ||||
| Internal Capsule | 51 | 0.252 | <0.001 | 4.48 | −11.2 | −5.6 | 7.7 | ||
| Left Hippocampus | 61 | 0.212 | <0.001 | 4.42 | −7.0 | −1.4 | 7.0 | ||
| Left Cortex | 26 | 0.415 | <0.001 | 4.07 | −18.9 | −7.7 | 11.2 | ||
| Internal Capsule | 40 | 0.310 | 0.001 | 4.00 | −8.4 | 20.3 | 3.5 | ||
| White | CONT < ID | None | - | - | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mudd, A.T.; Fil, J.E.; Knight, L.C.; Lam, F.; Liang, Z.-P.; Dilger, R.N. Early-Life Iron Deficiency Reduces Brain Iron Content and Alters Brain Tissue Composition Despite Iron Repletion: A Neuroimaging Assessment. Nutrients 2018, 10, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10020135
Mudd AT, Fil JE, Knight LC, Lam F, Liang Z-P, Dilger RN. Early-Life Iron Deficiency Reduces Brain Iron Content and Alters Brain Tissue Composition Despite Iron Repletion: A Neuroimaging Assessment. Nutrients. 2018; 10(2):135. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10020135
Chicago/Turabian StyleMudd, Austin T., Joanne E. Fil, Laura C. Knight, Fan Lam, Zhi-Pei Liang, and Ryan N. Dilger. 2018. "Early-Life Iron Deficiency Reduces Brain Iron Content and Alters Brain Tissue Composition Despite Iron Repletion: A Neuroimaging Assessment" Nutrients 10, no. 2: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10020135