Fatty Acid Lingual Application Activates Gustatory and Reward Brain Circuits in the Mouse

 , ,
, , 
Abstract
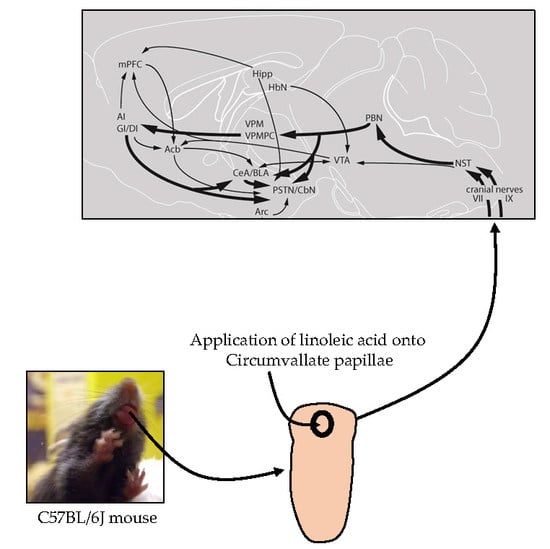
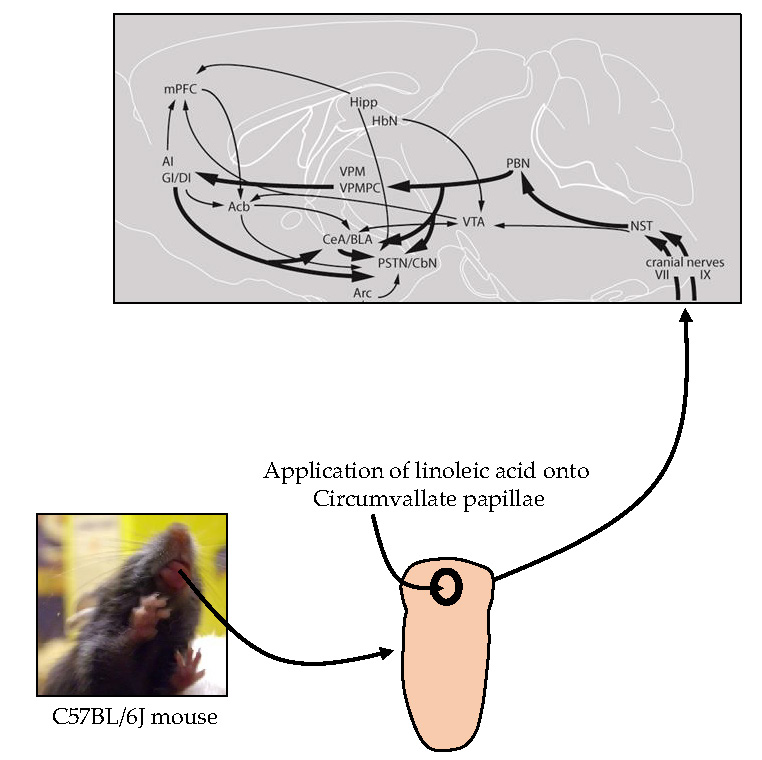
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Set-Up
2.2. Immunohistochemistry
Fos Immunostaining
2.3. Quantification of c-Fos Immunopositive Neurons
2.4. Statistical Analysis
3. Results
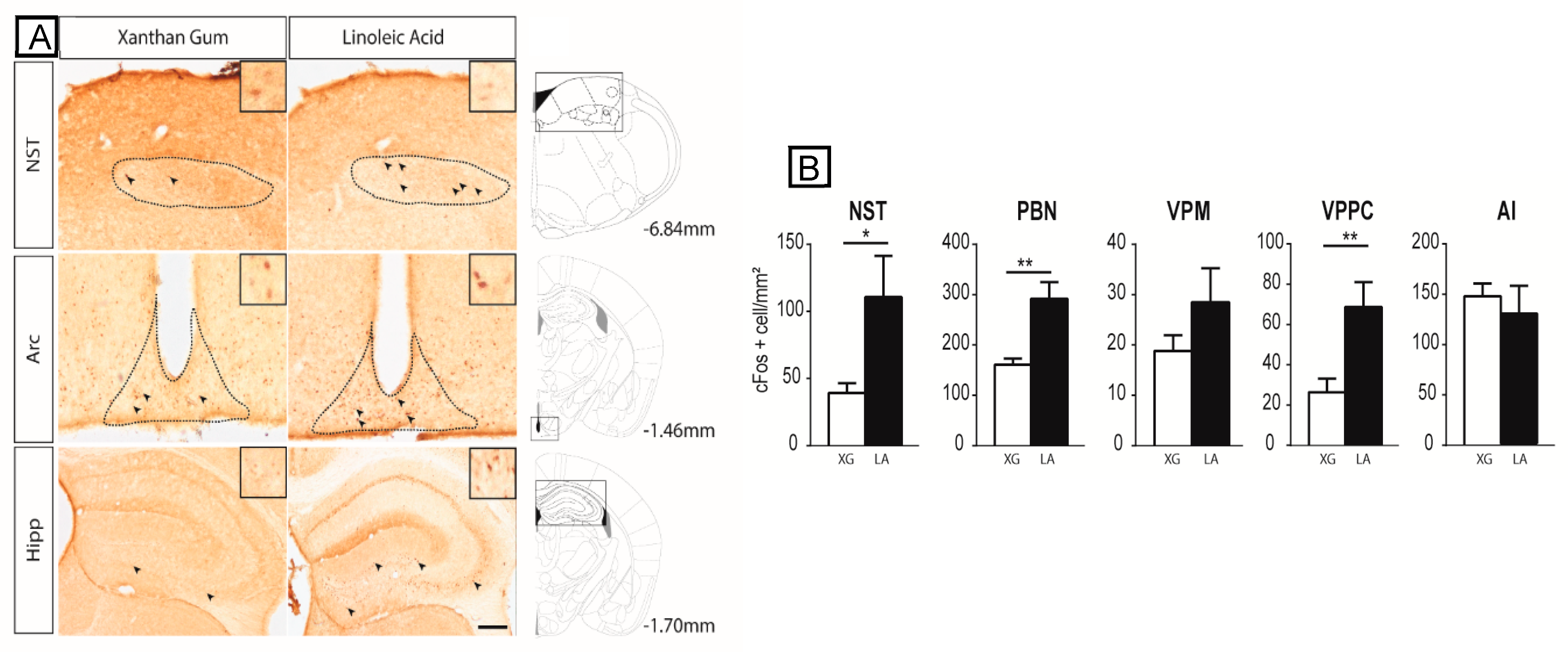
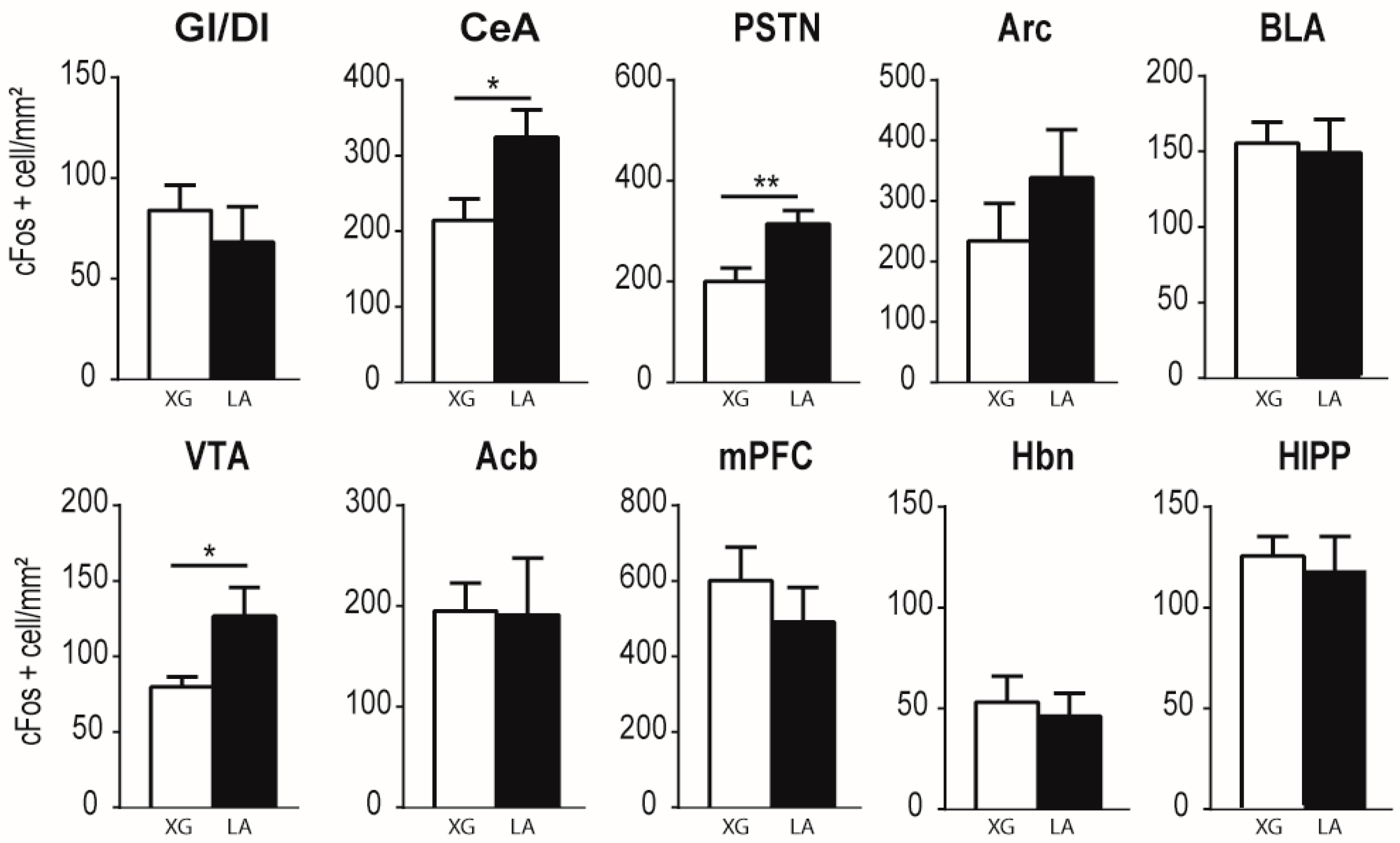
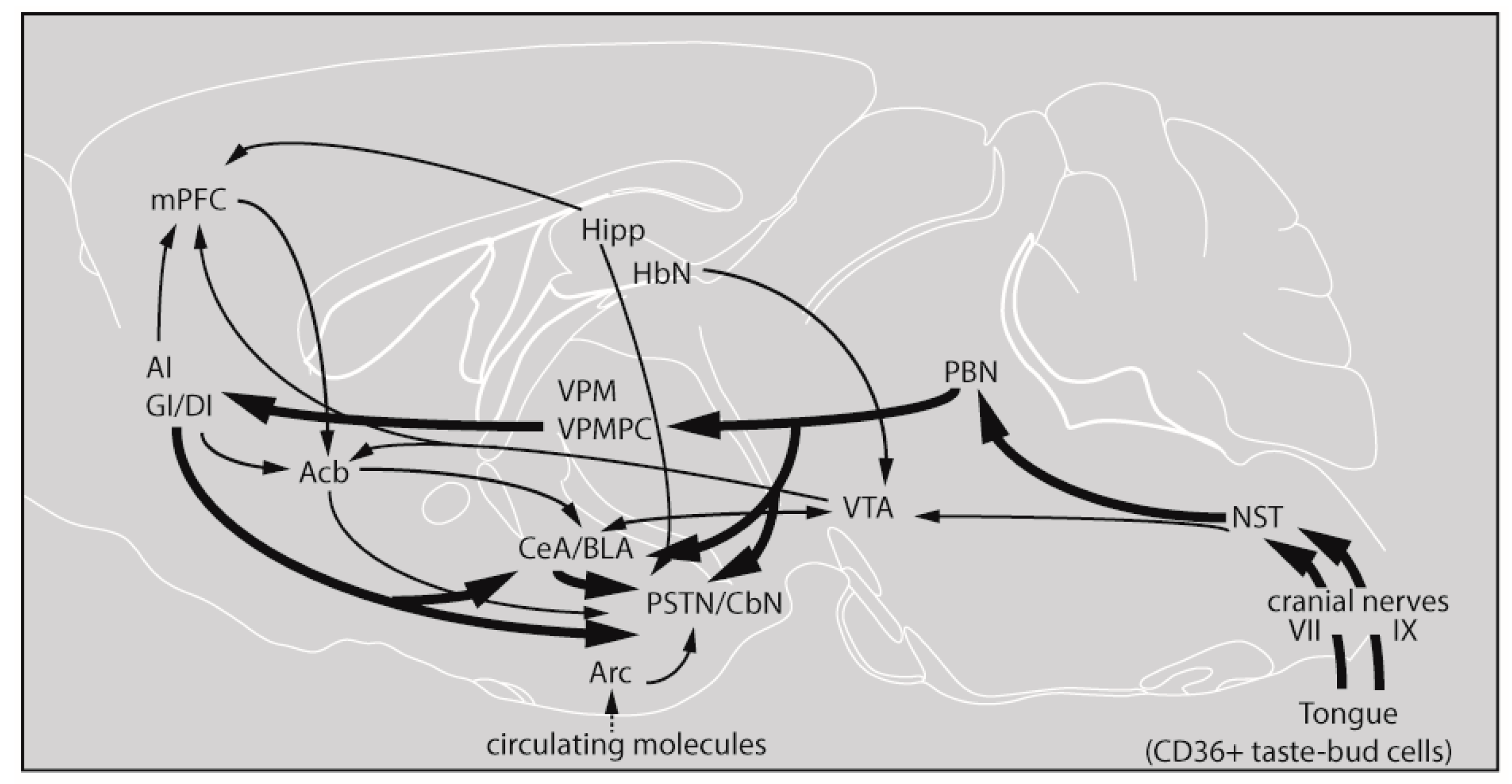
3.1. Lingual LA Stimulation Triggers Neuronal Activation of the Canonical Central Cerebral Gustatory Reward Pathway
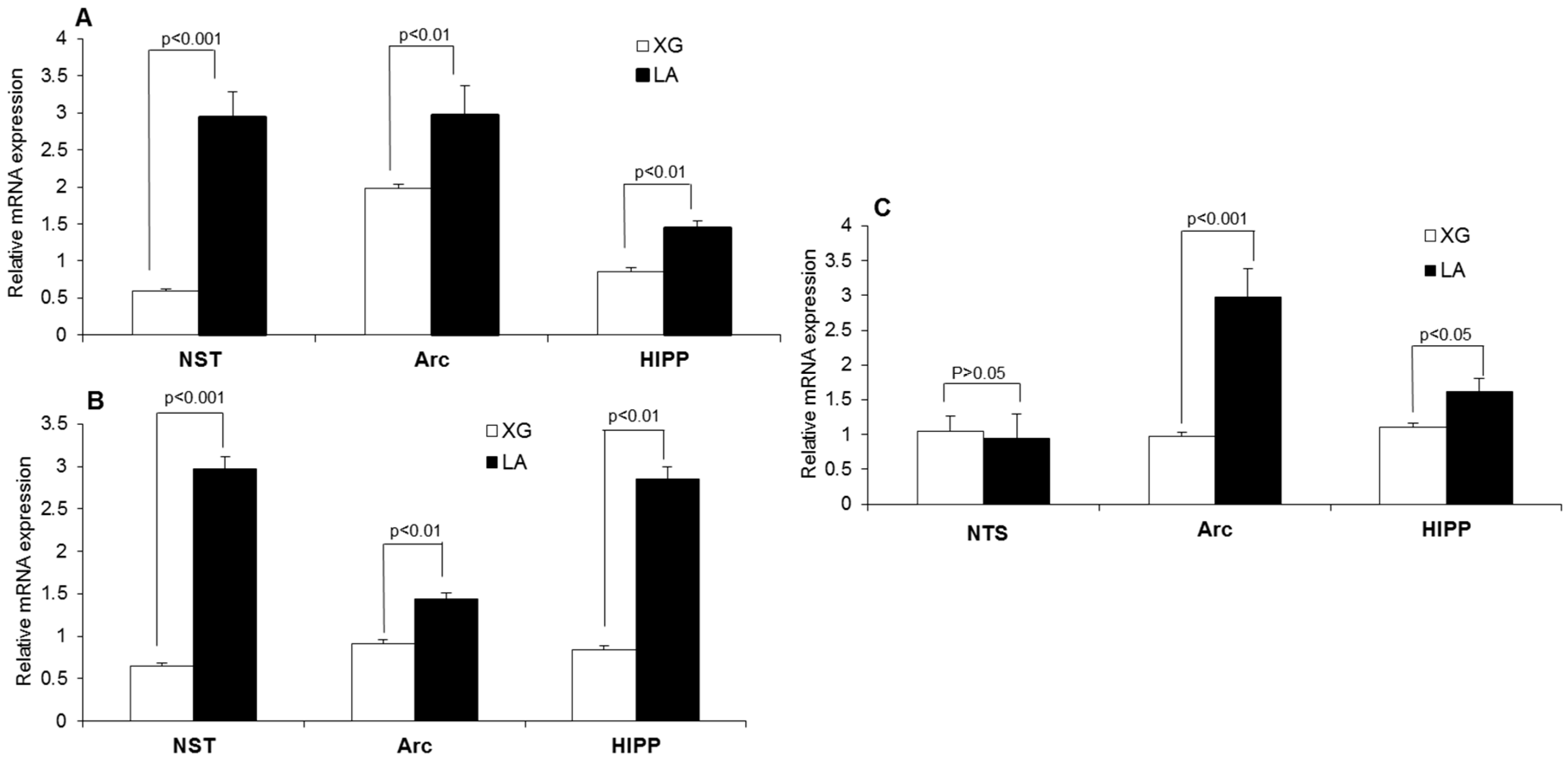
3.2. Lingual LA Stimulation Modulates the Expression of mRNA Encoding BDNF, Zif-268 and Glut-1
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hess, M.A. Resetting the American table—Creating a new alliance of taste and health. J. Am. Diet. Assoc. 1991, 91, 228–230. [Google Scholar] [PubMed]
- Chandrashekar, J.; Hoon, M.A.; Ryba, N.J.; Zuker, C.S. The receptors and cells for mammalian taste. Nature 2006, 444, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Roper, S.D. Signal transduction and information processing in mammalian taste buds. Pflugers Arch. 2007, 454, 759–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, P.; Passilly-Degrace, P.; Khan, N.A. Taste of fat: A sixth taste modality? Physiol. Rev. 2016, 96, 151–176. [Google Scholar] [CrossRef] [PubMed]
- Ozdener, M.H.; Subramaniam, S.; Sundaresan, S.; Sery, O.; Hashimoto, T.; Asakawa, Y.; Besnard, P.; Abumrad, N.A.; Khan, N.A. CD36-and GPR120-mediated Ca2+ signaling in human taste bud cells mediates differential responses to fatty acids and is altered in obese mice. Gastroenterology 2014, 146, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, T.A.; Khan, N.A. Cell signaling mechanisms of oro-gustatory detection of dietary fat: Advances and challenges. Prog. Lipid Res. 2014, 53, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, D.; Laugerette, F.; Darcel, N.; El-Yassimi, A.; Passilly-Degrace, P.; Hichami, A.; Khan, N.A.; Montmayeur, J.-P.; Besnard, P. The gustatory pathway is involved in CD36-mediated orosensory perception of long-chain fatty acids in the mouse. FASEB J. 2008, 22, 1458–1468. [Google Scholar] [CrossRef] [PubMed]
- Kadohisa, M.; Verhagen, J.; Rolls, E. The primate amygdala: Neuronal representations of the viscosity, fat texture, temperature, grittiness and taste of foods. Neuroscience 2005, 132, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.V.; Rolls, E.T.; Kadohisa, M. Neurons in the primate orbitofrontal cortex respond to fat texture independently of viscosity. J. Neurophysiol. 2003, 90, 1514–1525. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E. Taste, olfactory and food texture reward processing in the brain and obesity. Int. J. Obes. 2011, 35, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.V.; Kadohisa, M.; Rolls, E.T. Primate insular/opercular taste cortex: Neuronal representations of the viscosity, fat texture, grittiness, temperature, and taste of foods. J. Neurophysiol. 2004, 92, 1685–1699. [Google Scholar] [CrossRef] [PubMed]
- Eldeghaidy, S.; Marciani, L.; Hort, J.; Hollowood, T.; Singh, G.; Bush, D.; Foster, T.; Taylor, A.J.; Busch, J.; Spiller, R.C. Prior consumption of a fat meal in healthy adults modulates the brain’s response to fat. J. Nutr. 2016, 146, 2187–2198. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, I.E.; Rolls, E.T. Representation in the human brain of food texture and oral fat. J. Neurosci. 2004, 24, 3086–3093. [Google Scholar] [CrossRef] [PubMed]
- Eldeghaidy, S.; Marciani, L.; McGlone, F.; Hollowood, T.; Hort, J.; Head, K.; Taylor, A.J.; Busch, J.; Spiller, R.C.; Gowland, P.A. The cortical response to the oral perception of fat emulsions and the effect of taster status. J. Neurophysiol. 2011, 105, 2572–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabenhorst, F.; Rolls, E.T.; Parris, B.A.; d’Souza, A.A. How the brain represents the reward value of fat in the mouth. Cereb. Cortex 2010, 20, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Burger, K.S.; Yokum, S. Relative ability of fat and sugar tastes to activate reward, gustatory, and somatosensory regions. Am. J. Clin. Nutr. 2013, 98, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Archer, N.; Duesing, K.; Hannan, G.; Keast, R. Mechanism of fat taste perception: Association with diet and obesity. Prog. Lipid Res. 2016, 63, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Hichami, A.; Datiche, F.; Ullah, S.; Liénard, F.; Chardigny, J.-M.; Cattarelli, M.; Khan, N.A. Olfactory discrimination ability and brain expression of c-Fos, Gir and Glut1 mRNA are altered in n-3 fatty acid-depleted rats. Behav. Brain Res. 2007, 184, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K. The Mouse Brain Atlas in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Gal-Ben-Ari, S.; Kenney, J.W.; Ounalla-Saad, H.; Taha, E.; David, O.; Levitan, D.; Gildish, I.; Panja, D.; Pai, B.; Wibrand, K. Consolidation and translation regulation. Learn. Mem. 2012, 19, 410–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, G.E.; Smith, M.S.; Verbalis, J.G. C-fos and related immediate early gene products as markers of activity in neuroendocrine systems. Front. Neuroendocrinol. 1993, 14, 173–213. [Google Scholar] [CrossRef] [PubMed]
- Morin, J.P.; Quiroz, C.; Mendoza-Viveros, L.; Ramirez-Amaya, V.; Bermudez-Rattoni, F. Familiar taste induces higher dendritic levels of activity-regulated cytoskeleton-associated protein in the insular cortex than a novel one. Learn. Mem. 2011, 18, 610–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laugerette, F.; Passilly-Degrace, P.; Patris, B.; Niot, I.; Febbraio, M.; Montmayeur, J.-P.; Besnard, P. CD36 involvement in orosensory detection of dietary lipids, spontaneous fat preference, and digestive secretions. J. Clin. Investig. 2005, 115, 3177–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, R.A. Brain reward circuitry: Insights from unsensed incentives. Neuron 2002, 36, 229–240. [Google Scholar] [CrossRef]
- Dela Cruz, J.A.; Coke, T.; Bodnar, R.J. Simultaneous detection of c-fos activation from mesolimbic and mesocortical dopamine reward sites following naive sugar and fat ingestion in rats. J. Vis. Exp. 2016, 114, e53897. [Google Scholar] [CrossRef] [PubMed]
- Chometton, S.; Pedron, S.; Peterschmitt, Y.; Van Waes, V.; Fellmann, D.; Risold, PY. A premammillary lateral hypothalamic nuclear complex responds to hedonic but not aversive tastes in the male rat. Brain Struct. Funct. 2016, 221, 2183–2208. [Google Scholar] [CrossRef] [PubMed]
- Barbier, M.; Chometton, S.; Peterschmitt, Y.; Fellmann, D.; Risold, P.Y. Parasubthalamic and calbindin nuclei in the posterior lateral hypothalamus are the major hypothalamic targets for projections from the central and anterior basomedial nuclei of the amygdala. Brain Struct. Funct. 2017, 222, 2961–2991. [Google Scholar] [CrossRef] [PubMed]
- Murer, M.G.; Yan, Q.; Raisman-Vozari, R. Brain-derived neurotrophic factor in the control human brain, and in alzheimer’s disease and parkinson’s disease. Prog. Neurobiol. 2001, 63, 71–124. [Google Scholar] [CrossRef]
- Alonso, M.; Vianna, M.R.; Depino, A.M.; Mello e Souza, T.; Pereira, P.; Szapiro, G.; Viola, H.; Pitossi, F.; Izquierdo, I.; Medina, J.H. Bdnf-triggered events in the rat hippocampus are required for both short- and long-term memory formation. Hippocampus 2002, 12, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Genoud, C.; Knott, G.W.; Sakata, K.; Lu, B.; Welker, E. Altered synapse formation in the adult somatosensory cortex of brain-derived neurotrophic factor heterozygote mice. J. Neurosci. 2004, 24, 2394–2400. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, M.E.; Gafford, G.M.; Jarome, T.J.; Helmstetter, F.J. Time-dependent expression of arc and zif268 after acquisition of fear conditioning. Neural. Plast. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Errington, M.; French, P.; Fine, A.; Bliss, T.; Garel, S.; Charnay, P.; Bozon, B.; Laroche, S.; Davis, S. A requirement for the immediate early gene zif268 in the expression of late ltp and long-term memories. Nat. Neurosci. 2001, 4, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Maher, F.; Davies-Hill, T.M.; Lysko, P.G.; Henneberry, R.C.; Simpson, I.A. Expression of two glucose transporters, glut1 and glut3, in cultured cerebellar neurons: Evidence for neuron-specific expression of glut3. Mol. Cell Neurosci. 1991, 2, 351–360. [Google Scholar] [CrossRef]
- Seidner, G.; Alvarez, M.G.; Yeh, J.-I.; O’Driscoll, K.R.; Klepper, J.; Stump, T.S.; Wang, D.; Spinner, N.B.; Birnbaum, M.J.; Darryl, C. Glut-1 deficiency syndrome caused by haploinsufficiency of the blood-brain barrier hexose carrier. Nat. Genet. 1998, 18, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.F.; Santos, J.R.; Meurer, Y.S.; Macedo, P.T.; Ferreira, L.M.; Pontes, I.M.; Ribeiro, A.M.; Silva, R.H. Differential cortical c-fos and zif-268 expression after object and spatial memory processing in a standard or episodic-like object recognition task. Front. Behav. Neurosci. 2013, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.G.; Pereira, A.C.; Botelho, E.P.; Pereira, S.S.; Fiorani, M.; Gattass, R. Differential expression of zif268 and c-fos in the primary visual cortex and lateral geniculate nucleus of normal cebus monkeys and after monocular lesions. J. Comp. Neurol. 2005, 482, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Aydin-Abidin, S.; Trippe, J.; Funke, K.; Eysel, U.T.; Benali, A. High-and low-frequency repetitive transcranial magnetic stimulation differentially activates c-fos and zif268 protein expression in the rat brain. Exp. Brain Res. 2008, 188, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Hauguel-de Mouzon, S.; Leturque, A.; Alsat, E.; Loizeau, M.; Evain-Brion, D.; Girard, J. Developmental expression of glut1 glucose transporter and c-fos genes in human placental cells. Placenta 1994, 15, 35–46. [Google Scholar] [CrossRef]
- Kreisberg, J.I.; Radnik, R.A.; Ayo, S.H.; Garoni, J.; Saikumar, P. High glucose elevates c-fos and c-jun transcripts and proteins in mesangial cell cultures. Kidney Int. 1994, 46, 105–112. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterschmitt, Y.; Abdoul-Azize, S.; Murtaza, B.; Barbier, M.; Khan, A.S.; Millot, J.-L.; Khan, N.A. Fatty Acid Lingual Application Activates Gustatory and Reward Brain Circuits in the Mouse. Nutrients 2018, 10, 1246. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10091246
Peterschmitt Y, Abdoul-Azize S, Murtaza B, Barbier M, Khan AS, Millot J-L, Khan NA. Fatty Acid Lingual Application Activates Gustatory and Reward Brain Circuits in the Mouse. Nutrients. 2018; 10(9):1246. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10091246
Chicago/Turabian StylePeterschmitt, Yvan, Souleymane Abdoul-Azize, Babar Murtaza, Marie Barbier, Amira Sayed Khan, Jean-Louis Millot, and Naim Akhtar Khan. 2018. "Fatty Acid Lingual Application Activates Gustatory and Reward Brain Circuits in the Mouse" Nutrients 10, no. 9: 1246. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10091246