Spirulina Liquid Extract Protects against Fibrosis Related to Non-Alcoholic Steatohepatitis and Increases Ursodeoxycholic Acid

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spirulina Liquid Extract
2.2. Mice and Diets
2.3. Blood Analyses and Tissue Collection
2.4. Liver Steatosis and Fibrosis Quantification
2.5. Western Blot
2.6. Superoxide Anion (O2−) and Nitric Oxide (NO) Measurements by Electronic Paramagnetic Resonance (EPR)
2.7. Liver Staining and Confocal Microscopy Imaging
2.8. Glucose Tolerance Test
2.9. Real-Time qRT-PCR
2.10. Statistics
3. Results
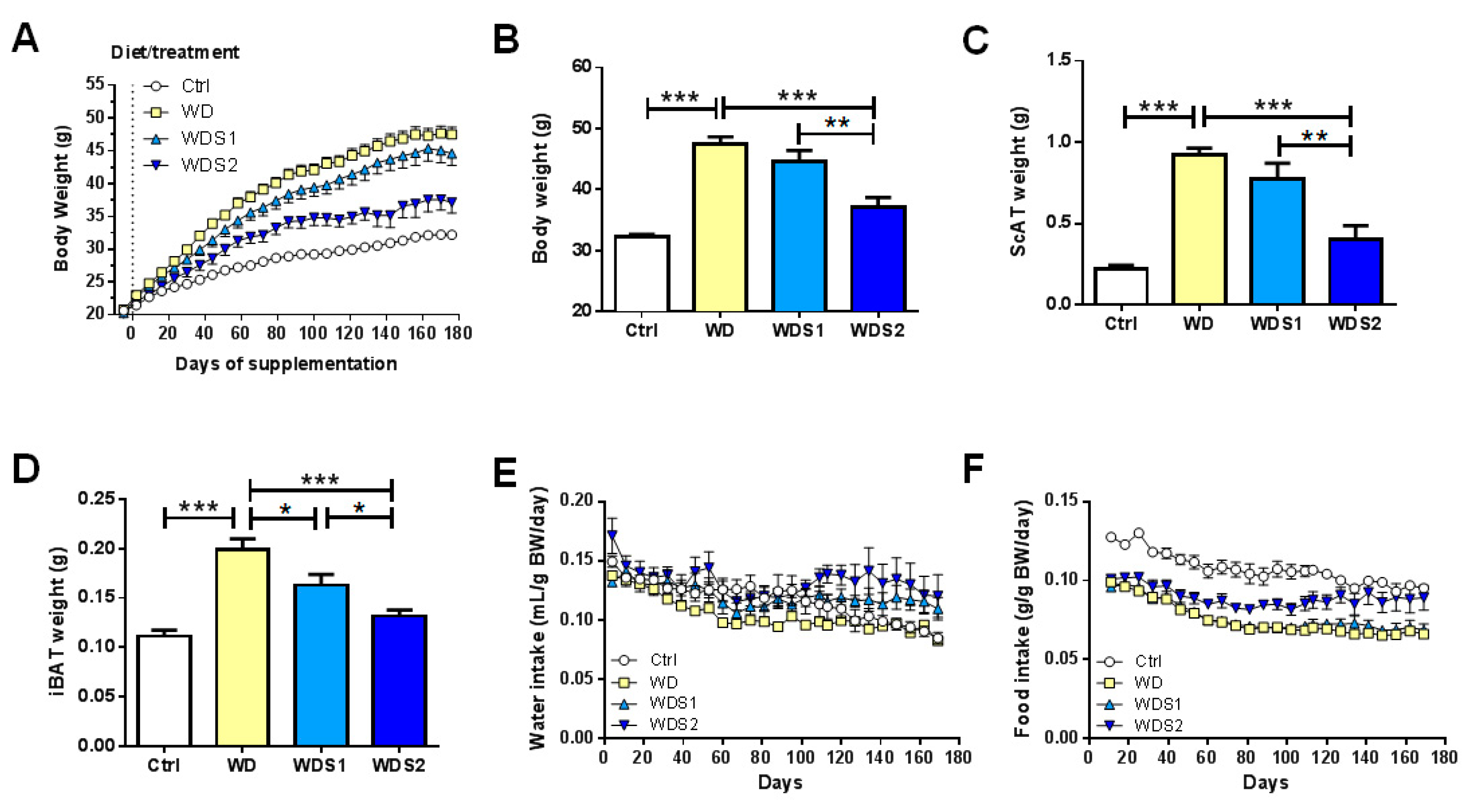
3.1. Spirulina Alleviates Diet Induced Body Weight Gain
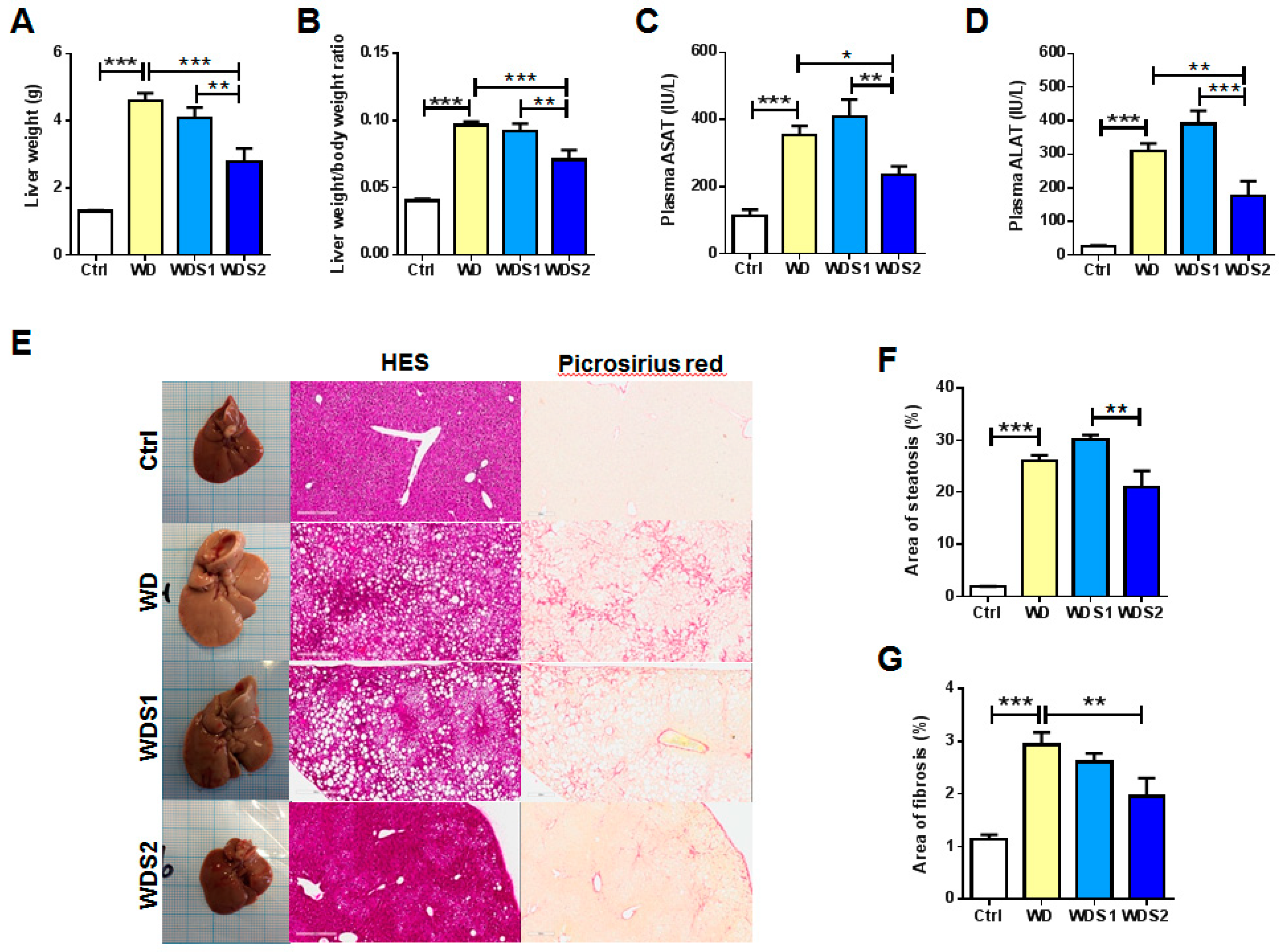
3.2. NASH and Liver Fibrosis is Prevented with Spirulina Supplementation
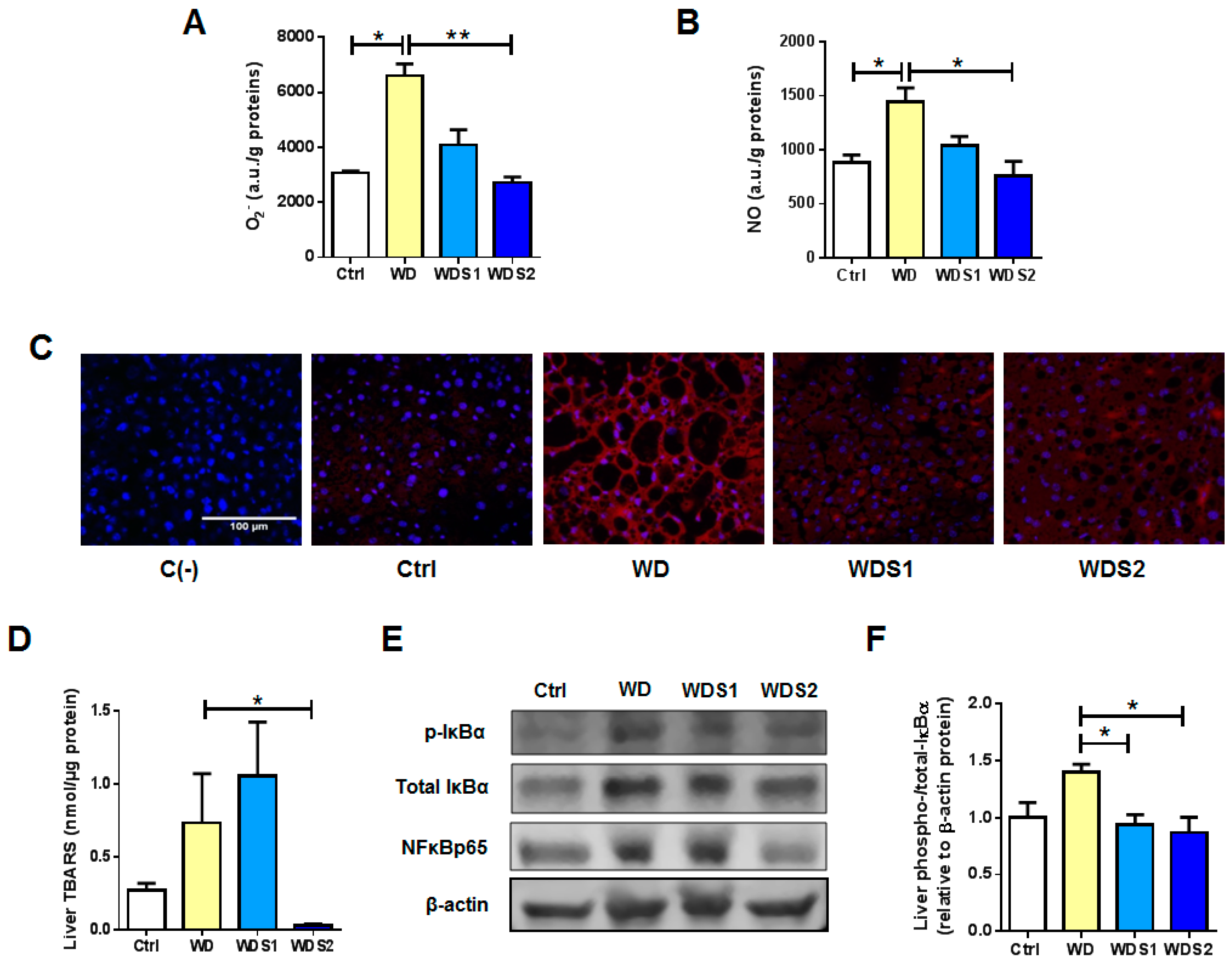
3.3. Liver from Spirulina Supplemented Mice Are Protected against Reactive Oxygen and nitrogen Species Accumulation Related to WD
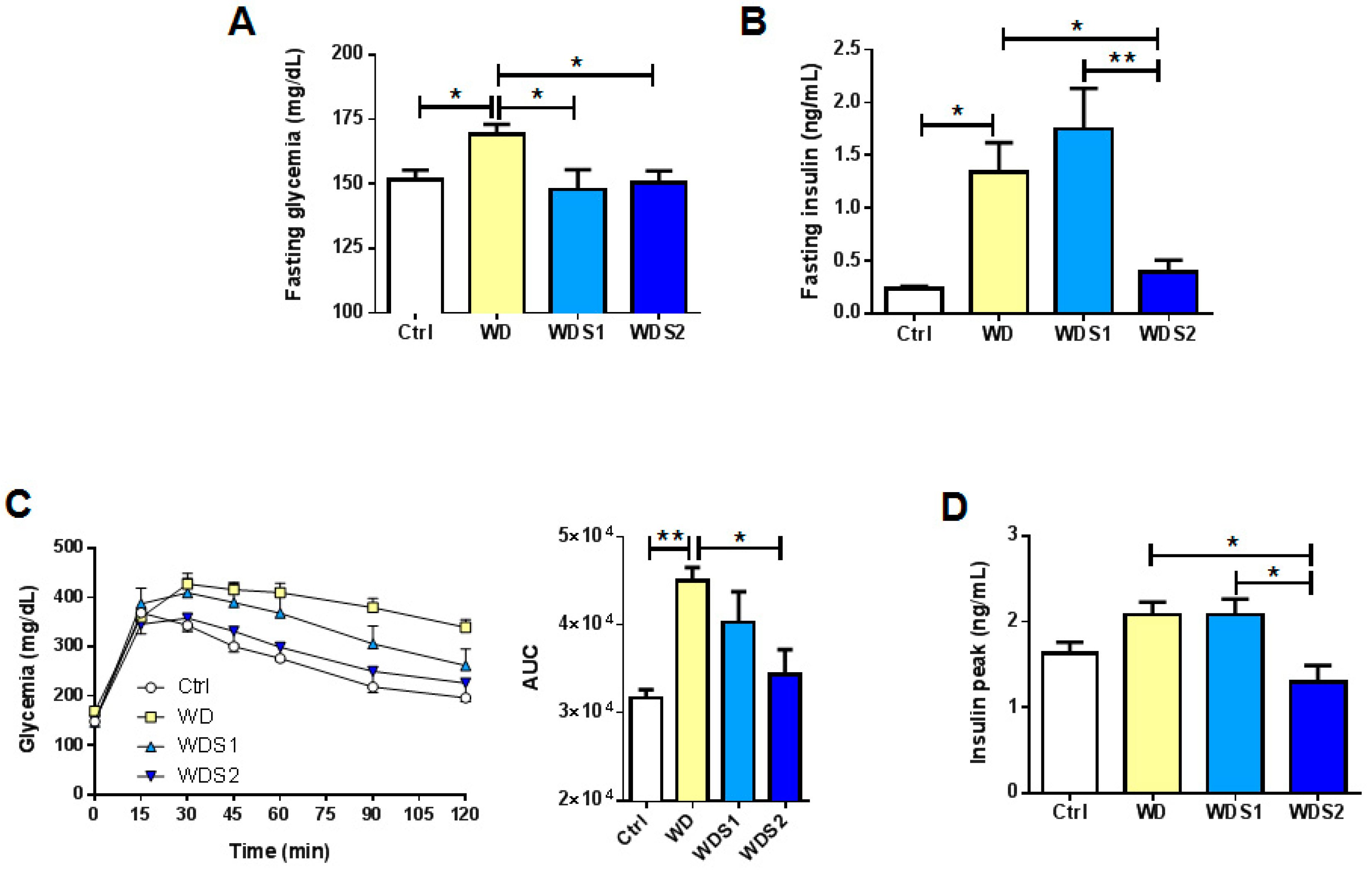
3.4. WD Induced Insulin Resistance Is Reduced with SLE Supplementation
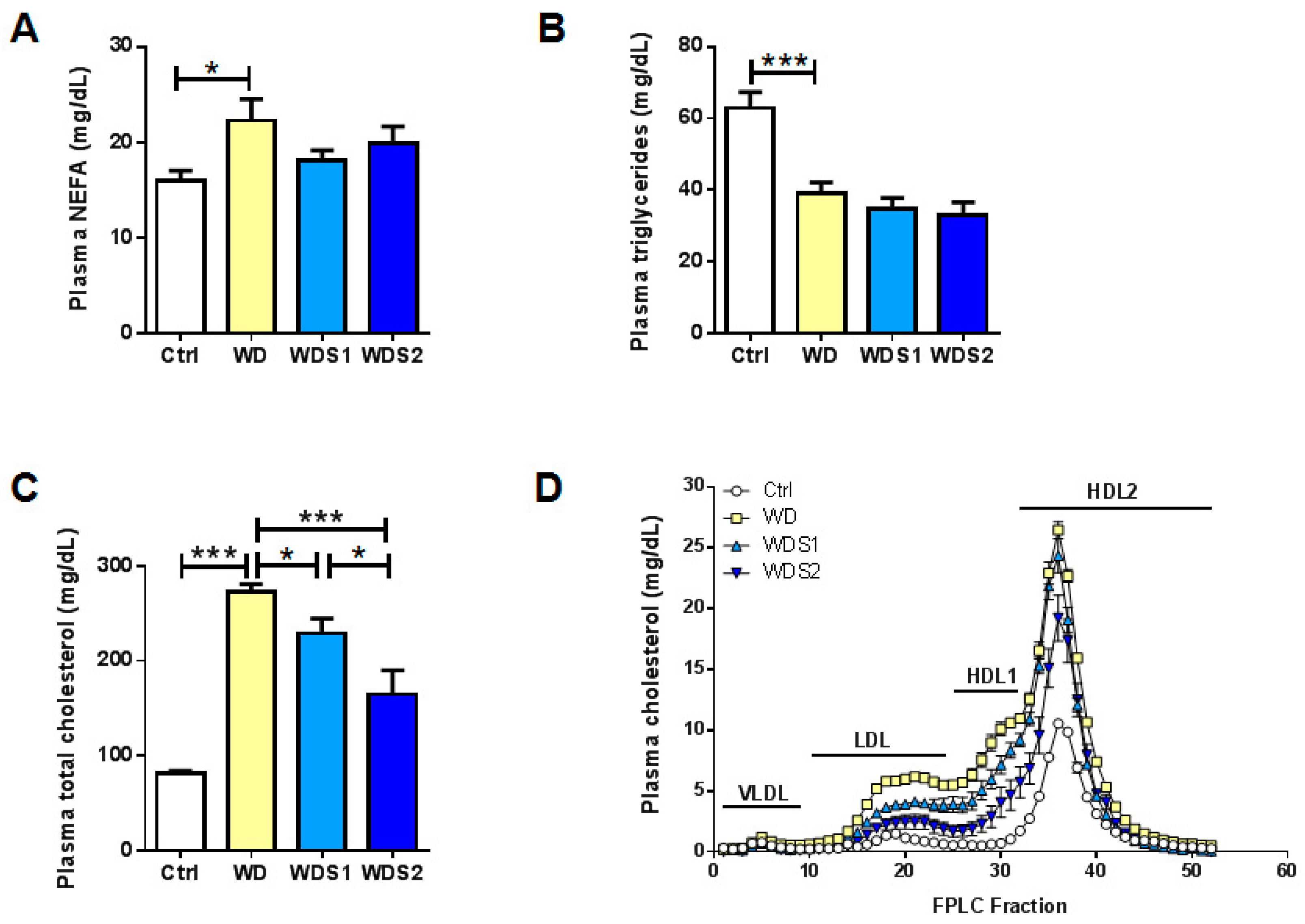
3.5. SLE Maintains a Normal Plasma Lipid Profile as a Marker of Liver Functionality
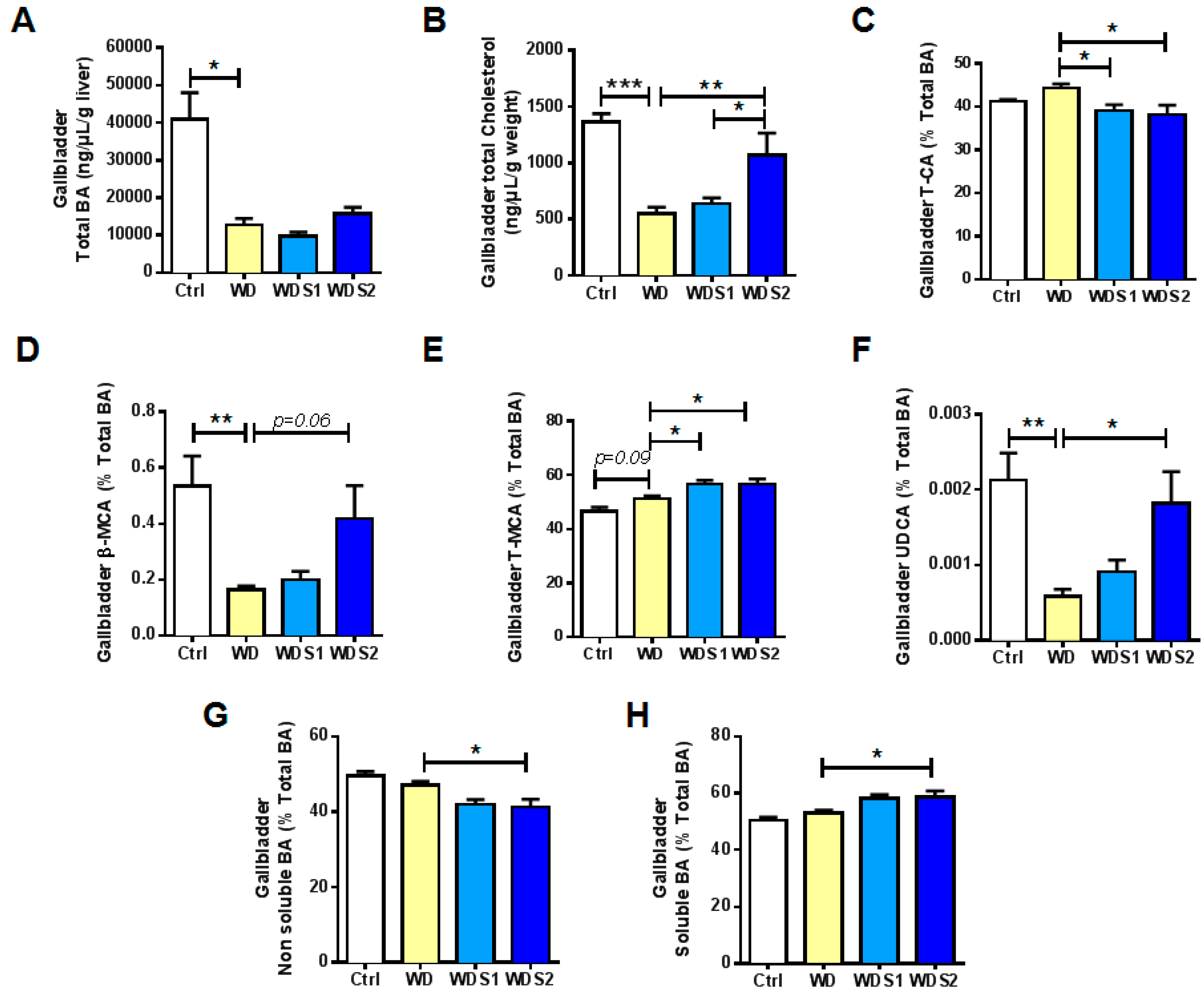
3.6. SLE Promotes a Less Hydrophobic Bile Acid Profile and Increased Ursodeoxycholic Acid Concentration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dulai, P.S.; Singh, S.; Patel, J.; Soni, M.; Prokop, L.J.; Younossi, Z.; Sebastiani, G.; Ekstedt, M.; Hagstrom, H.; Nasr, P.; et al. Increased risk of mortality by fibrosis stage in nonalcoholic fatty liver disease: Systematic review and meta-analysis. Hepatology 2017, 65, 1557–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Learned, R.M.; Rossi, S.J.; DePaoli, A.M.; Tian, H.; Ling, L. Engineered FGF19 eliminates bile acid toxicity and lipotoxicity leading to resolution of steatohepatitis and fibrosis in mice. Hepatol. Commun. 2017, 1, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62 (Suppl. 1), S47–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konerman, M.A.; Jones, J.C.; Harrison, S.A. Pharmacotherapy for NASH: Current and emerging. J. Hepatol. 2018, 68, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Golabi, P.; Bush, H.; Younossi, Z.M. Treatment strategies for nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Clin. Liver Dis. 2017, 21, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Riss, J.; Décordé, K.; Sutra, T.; Delage, M.; Baccou, J.C.; Jouy, N.; Brune, J.P.; Oréal, H.; Cristol, J.P.; Rouanet, J.M. Phycobiliprotein C-phycocyanin from Spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters. J. Agric. Food Chem. 2007, 55, 7962–7967. [Google Scholar] [CrossRef]
- Romay, C.; Armesto, J.; Remirez, D.; González, R.; Ledon, N.; García, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef]
- Romay, C.; Delgado, R.; Remirez, D.; González, R.; Rojas, A. Effects of phycocyanin extract on tumor necrosis factor-alpha and nitrite levels in serum of mice treated with endotoxin. Arzneimittelforschung 2001, 51, 733–736. [Google Scholar]
- Satapati, S.; Kucejova, B.; Duarte, J.A.G.; Fletcher, J.A.; Reynolds, L.; Sunny, N.E.; He, T.; Nair, L.A.; Livingston, K.; Fu, X.; et al. Mitochondrial metabolism mediates oxidative stress and inflammation in fatty liver. J. Clin. Investig. 2015, 125, 4447–4462. [Google Scholar] [CrossRef] [Green Version]
- Spahis, S.; Delvin, E.; Borys, J.M.; Levy, E. Oxidative stress as a critical factor in nonalcoholic fatty liver disease pathogenesis. Antioxid. Redox Signal. 2017, 26, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L. Targeting bile acids and lipotoxicity for NASH treatment. Hepatol. Commun. 2017, 1, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Chow, T.J. Hypolipidemic, antioxidant, and antiinflammatory activities of microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.S.; Yang, Y.; Park, Y.; Lee, J. Health benefits of blue-green algae: Prevention of cardiovascular disease and nonalcoholic fatty liver disease. J. Med. Food 2013, 16, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Pak, W.; Takayama, F.; Mine, M.; Nakamoto, K.; Kodo, Y.; Mankura, M.; Egashira, T.; Kawasaki, H.; Mori, A. Anti-oxidative and anti-inflammatory effects of spirulina on rat model of non-alcoholic steatohepatitis. J. Clin. Biochem. Nutr. 2012, 51, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chetiveaux, M.; Nazih, H.; Ferchaud-Roucher, V.; Lambert, G.; Zaïr, Y.; Masson, M.; Ouguerram, K.; Bouhours, D.; Krempf, M. The differential apoA-I enrichment of prebeta1 and alphaHDL is detectable by gel filtration separation. J. Lipid Res. 2002, 43, 1986–1993. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, V.; Ramin-Mangata, S.; Billon-Crossouard, S.; Aguesse, A.; Durand, M.; Chemello, K.; Nativel, B.; Flet, L.; Chétiveaux, M.; Jacobi, D.; et al. Kinetics of plasma apolipoprotein E isoforms by LC-MS/MS: A pilot study. J. Lipid Res. 2018, 59, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Trenteseaux, C.; Gaston, A.T.; Aguesse, A.; Poupeau, G.; de Coppet, P.; Andriantsitohaina, R.; Laschet, J.; Amarger, V.; Krempf, M.; Nobecourt-Dupuy, E.; et al. Perinatal hypercholesterolemia exacerbates atherosclerosis lesions in offspring by altering metabolism of trimethylamine-N-oxide and bile acids. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2053–2063. [Google Scholar] [CrossRef]
- Yagi, K. A simple fluorometric assay for lipoperoxide in blood plasma. Biochem. Med. 1976, 15, 212–216. [Google Scholar] [CrossRef]
- Boursier, J.; de Ledinghen, V.; Sturm, N.; Amrani, L.; Bacq, Y.; Sandrini, J.; Le Bail, B.; Chaigneau, J.; Zarski, J.P.; Gallois, Y.; et al. Precise evaluation of liver histology by computerized morphometry shows that steatosis influences liver stiffness measured by transient elastography in chronic hepatitis C. J. Gastroenterol. 2014, 49, 527–537. [Google Scholar] [CrossRef]
- Chalopin, M.; Tesse, A.; Martínez, M.C.; Rognan, D.; Arnal, J.F.; Andriantsitohaina, R. Estrogen receptor alpha as a key target of red wine polyphenols action on the endothelium. PLoS ONE 2010, 5, e8554. [Google Scholar] [CrossRef] [PubMed]
- Miczke, A.; Szulińska, M.; Hansdorfer-Korzon, R.; Kręgielska-Narożna, M.; Suliburska, J.; Walkowiak, J.; Bogdański, P. Effects of spirulina consumption on body weight, blood pressure, and endothelial function in overweight hypertensive Caucasians: A double-blind, placebo-controlled, randomized trial. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 150–156. [Google Scholar] [PubMed]
- Mazokopakis, E.E.; Papadomanolaki, M.G.; Fousteris, A.A.; Kotsiris, D.A.; Lampadakis, I.M.; Ganotakis, E.S. The hepatoprotective and hypolipidemic effects of Spirulina (Arthrospira platensis) supplementation in a Cretan population with non-alcoholic fatty liver disease: A prospective pilot study. Ann. Gastroenterol. 2014, 27, 387–394. [Google Scholar] [PubMed]
- Yousefi, R.; Mottaghi, A.; Saidpour, A. Spirulina platensis effectively ameliorates anthropometric measurements and obesity-related metabolic disorders in obese or overweight healthy individuals: A randomized controlled trial. Complement. Ther. Med. 2018, 40, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Matip, M.E.; Pieme, C.A.; Azabji-Kenfack, M.; Biapa, P.C.; Germaine, N.; Heike, E.; Moukette, B.M.; Emmanuel, K.; Philippe, S.; Mbofung, C.M.; et al. Effects of Spirulina platensis supplementation on lipid profile in HIV-infected antiretroviral naive patients in Yaounde-Cameroon: A randomized trial study. Lipids Health Dis. 2014, 13, 191. [Google Scholar] [CrossRef]
- Samuels, R.; Mani, U.V.; Iyer, U.M.; Nayak, U.S. Hypocholesterolemic effect of spirulina in patients with hyperlipidemic nephrotic syndrome. J. Med. Food 2002, 5, 91–96. [Google Scholar] [CrossRef]
- Fujimoto, M.; Tsuneyama, K.; Fujimoto, T.; Selmi, C.; Gershwin, M.E.; Shimada, Y. Spirulina improves non-alcoholic steatohepatitis, visceral fat macrophage aggregation, and serum leptin in a mouse model of metabolic syndrome. Dig. Liver Dis. 2012, 44, 767–774. [Google Scholar] [CrossRef]
- Ou, Y.; Lin, L.; Yang, X.; Pan, Q.; Cheng, X. Antidiabetic potential of phycocyanin: Effects on KKAy mice. Pharm. Biol. 2013, 51, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D. Loss of stearoyl-CoA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [Green Version]
- Zeinalian, R.; Farhangi, M.A.; Shariat, A.; Saghafi-Asl, M. The effects of Spirulina Platensis on anthropometric indices, appetite, lipid profile and serum vascular endothelial growth factor (VEGF) in obese individuals: A randomized double blinded placebo controlled trial. BMC Complement. Altern. Med. 2017, 17, 225. [Google Scholar] [CrossRef]
- Nagaoka, S.; Shimizu, K.; Kaneko, H.; Shibayama, F.; Morikawa, K.; Kanamaru, Y.; Otsuka, A.; Hirahashi, T.; Kato, T. A novel protein C-phycocyanin plays a crucial role in the hypocholesterolemic action of Spirulina platensis concentrate in rats. J. Nutr. 2005, 135, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Farrell, G.C.; van Rooyen, D.; Gan, L.; Chitturi, S. NASH is an inflammatory disorder: Pathogenic, prognostic and therapeutic implications. Gut Liver 2012, 6, 149–171. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Sanchez, N.; Cruz-Ramon, V.C.; Ramirez-Perez, O.L.; Hwang, J.P.; Barranco-Fragoso, B.; Cordova-Gallardo, J. New aspects of lipotoxicity in nonalcoholic steatohepatitis. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Farrell, G.C.; Haczeyni, F.; Chitturi, S. Pathogenesis of NASH: How metabolic complications of overnutrition favour lipotoxicity and pro-inflammatory fatty liver disease. Adv. Exp. Med. Biol. 2018, 1061, 19–44. [Google Scholar] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.L.; Lim, Y.W.; Radhakrishnan, A.K.; Lim, P.E. Protective effect of aqueous extract from Spirulina platensis against cell death induced by free radicals. BMC Complement. Altern. Med. 2010, 10, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gdara, N.B.; Belgacem, A.; Khemiri, I.; Mannai, S.; Bitri, L. Protective effects of phycocyanin on ischemia/reperfusion liver injuries. Biomed. Pharmacother. 2018, 102, 196–202. [Google Scholar] [CrossRef]
- Lee, E.H.; Park, J.E.; Choi, Y.J.; Huh, K.B.; Kim, W.Y. A randomized study to establish the effects of spirulina in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 295–300. [Google Scholar] [CrossRef]
- Ou, Y.; Lin, L.; Pan, Q.; Yang, X.; Cheng, X. Preventive effect of phycocyanin from Spirulina platensis on alloxan-injured mice. Environ. Toxicol. Pharmacol. 2012, 34, 721–726. [Google Scholar] [CrossRef]
- Ou, Y.; Ren, Z.; Wang, J.; Yang, X. Phycocyanin ameliorates alloxan-induced diabetes mellitus in mice: Involved in insulin signaling pathway and GK expression. Chem. Biol. Interact. 2016, 247, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Gruen, M.L.; Plummer, M.R.; Zhang, W.; Posey, K.A.; Linton, M.F.; Fazio, S.; Hasty, A.H. Persistence of high density lipoprotein particles in obese mice lacking apolipoprotein A-I. J. Lipid Res. 2005, 46, 2007–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, S.M.; Perez-Tilve, D.; Greer, T.M.; Coburn, B.A.; Grant, E.; Basford, J.E.; Tschöp, M.H.; Hui, D.Y. Defective lipid delivery modulates glucose tolerance and metabolic response to diet in apolipoprotein E-deficient mice. Diabetes 2008, 57, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Kim, M.Y.; Sok, D.E.; Hwang, S.Y.; Kim, J.H.; Kim, H.R.; Lee, J.H.; Kim, Y.B.; Kim, M.R. Spirulina prevents atherosclerosis by reducing hypercholesterolemia in rabbits fed a high-cholesterol diet. J. Nutr. Sci. Vitaminol. 2010, 56, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Parikh, P.; Mani, U.; Iyer, U. Role of spirulina in the control of glycemia and lipidemia in type 2 diabetes mellitus. J. Med. Food 2001, 4, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Torres-Duran, P.V.; Ferreira-Hermosillo, A.; Juarez-Oropeza, M.A. Antihyperlipemic and antihypertensive effects of Spirulina maxima in an open sample of Mexican population: A preliminary report. Lipids Health Dis. 2007, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Beuers, U.; Trauner, M.; Jansen, P.; Poupon, R. New paradigms in the treatment of hepatic cholestasis: From UDCA to FXR, PXR and beyond. J. Hepatol. 2015, 62 (Suppl. 1), S25–S37. [Google Scholar] [CrossRef] [Green Version]
- Trauner, M.; Graziadei, I.W. Review article: Mechanisms of action and therapeutic applications of ursodeoxycholic acid in chronic liver diseases. Aliment. Pharmacol. Ther. 1999, 13, 979–996. [Google Scholar] [CrossRef]
- Arab, J.P.; Karpen, S.J.; Dawson, P.A.; Arrese, M.; Trauner, M. Bile acids and nonalcoholic fatty liver disease: Molecular insights and therapeutic perspectives. Hepatology 2017, 65, 350–362. [Google Scholar] [CrossRef]
- Sanchez-Garcia, A.; Sahebkar, A.; Simental-Mendía, M.; Simental-Mendía, L.E. Effect of ursodeoxycholic acid on glycemic markers: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2018, 135, 144–149. [Google Scholar] [CrossRef]
- Xiang, Z.; Chen, Y.P.; Ma, K.F.; Ye, Y.F.; Zheng, L.; Yang, Y.D.; Li, Y.M.; Jin, X. The role of ursodeoxycholic acid in non-alcoholic steatohepatitis: A systematic review. BMC Gastroenterol. 2013, 13, 140. [Google Scholar] [CrossRef]
- Poupon, R.E.; Ouguerram, K.; Chrétien, Y.; Verneau, C.; Eschwège, E.; Magot, T.; Poupon, R. Cholesterol-lowering effect of ursodeoxycholic acid in patients with primary biliary cirrhosis. Hepatology 1993, 17, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Innerarity, T.L.; Pitas, R.E.; Mahley, R.W. Modulating effects of canine high density lipoproteins on cholesteryl ester synthesis induced by beta-very low density lipoproteins in macrophages. Possible in vitro correlates with atherosclerosis. Arteriosclerosis 1982, 2, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Taminiau, B.; Walgrave, H.; Daube, G.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Spirulina protects against hepatic inflammation in aging: An effect related to the modulation of the gut microbiota? Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Ctrl | WD | WDS1 | WDS2 |
|---|---|---|---|---|
| Uptake | ||||
| Slc2a2 | 1 ± 0.06 | 0.95 ± 0.04 | 0.79 ± 0.12 | 0.63 ± 0.10 $ |
| Cd36 | 1 ± 0.10 | 7.54 ± 0.7 *** | 6..44 ± 0.96 | 4.3 ± 1.11 $ |
| Lipogenesis | ||||
| Acc-1 | 1 ± 0.06 | 1.05 ± 0.05 | 0.89 ± 0.14 | 0.65 ± 0.15 $ |
| Scd-1 | 1 ± 0.10 | 1.99 ± 0.14 *** | 1.68 ± 0.22 | 1.15 ± 0.19 $$ |
| Srebp-1 | 1 ± 0.06 | 1.71 ± 0.11 ** | 1.30 ± 0.19 | 1.29 ± 0.17 p=0.10 |
| Dgat-1 | 1 ± 0.04 | 1.13 ± 0.06 | 0.99 ± 0.09 | 0.89 ± 0.6 $ |
| Oxidation | ||||
| Ppar-α | 1 ± 0.06 | 1.08 ± 0.05 | 0.93 ± 0.08 | 0.76 ± 0.05 $$ |
| Cpt1a | 1 ± 0.03 | 2.09 ± 0.11 *** | 1.89 ± 0.2 | 1.68 ± 0.18 p=0.15 |
| Fibrogenesis | ||||
| Col1a1 | 1 ± 0.15 | 15.56 ± 0.87 *** | 14.32 ± 1.06 | 8.10 ± 2.73 $$ |
| Timp1 | 1 ± 0.16 | 55.78 ± 3.25 *** | 61.93 ± 7.40 | 27.99 ± 9.73 $ |
| Inflammation | ||||
| Tgf-β1 | 1 ± 0.10 | 2.12 ± 0.13 *** | 1.99 ± 0.09 | 1.46 ± 0.19 $$ |
| Tlr9 | 1 ± 0.14 | 4.20 ± 0.37 *** | 4.24 ± 0.37 | 2.10 ± 0.36 $$$ |
| Ctrl | WD | WDS1 | WDS2 | |
|---|---|---|---|---|
| Lipoproteins | ||||
| VLDL-C (mg/dL) | 3.9 ± 0.5 | 5.9 ± 0.3 | 3.3 ± 0.7 $ | 1.8 ± 0.4 $$$ |
| LDL-C (mg/dL) | 11.8 ± 0.3 | 57.2 ± 1.8 *** | 47.0 ± 5.2 | 30.2 ± 5.1 $$$ |
| HDL1-C (mg/dL) | 5.0 ± 0.3 | 50.1 ± 1.3 *** | 41.5 ± 5.2 | 27.4 ± 5.2 $$$ |
| HDL2-C (mg/dL) | 60.5 ± 3.1 | 160.8 ± 5.7 *** | 138.5 ± 6.8 | 106.3 ± 25.2 $$$ |
| Apolipoproteins | ||||
| Apo A-I (mg/dL) | 87.8 ± 8.6 | 946.7 ± 133.1 *** | 915.8 ± 192.3 | 419.1 ± 103.3 $ |
| Apo B100 (mg/dL) | 8.7 ± 1.0 | 19.9 ± 2.2 *** | 17.9 ± 1.4 | 13.3 ± 2.4 $ |
| Apo E (mg/dL) | 9.8 ± 0.4 | 60.0 ± 4.5 *** | 53.3 ± 7.3 | 29.2 ± 6.6 $$ |
| Apo C-II (mg/dL) | 0.7 ± 0.04 | 4.8 ± 0.3 *** | 3.8 ± 0.3 $ | 3.0 ± 0.3 $$$ |
| Apo C-III (mg/dL) | 8.9 ± 1.0 | 23.0 ± 3.1 *** | 15.6 ± 2.6 | 13.1 ± 2.1 $ |
| Gallbladder UDCA (% Total BA) | ||
|---|---|---|
| Variables | r | p Value |
| Body weight | −0.69 | <0.0001 |
| Fasting glycemia | −0.38 | 0.02 |
| Fasting insulinemia | −0.49 | 0.003 |
| AUC GTT | −0.45 | 0.006 |
| ScAT weight | −0.65 | <0.0001 |
| Plasma total cholesterol | −0.72 | <0.0001 |
| Liver weight/Body weight ratio | −0.69 | <0.0001 |
| Fibrosis | −0.67 | <0.0001 |
| Steatosis | −0.64 | <0.0001 |
| Plasma ASAT | −0.59 | 0.0003 |
| Plasma ALAT | −0.68 | <0.0001 |
| Liver O2− | −0.56 | 0.02 |
| Liver NO | −0.60 | 0.005 |
| Liver Scd1 mRNA | −0.63 | <0.0001 |
| Liver Tgfβ1 mRNA | −0.64 | <0.0001 |
| Liver Col1a1 mRNA | −0.70 | <0.0001 |
| Liver Timp1 mRNA | −0.65 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coué, M.; Tesse, A.; Falewée, J.; Aguesse, A.; Croyal, M.; Fizanne, L.; Chaigneau, J.; Boursier, J.; Ouguerram, K. Spirulina Liquid Extract Protects against Fibrosis Related to Non-Alcoholic Steatohepatitis and Increases Ursodeoxycholic Acid. Nutrients 2019, 11, 194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010194
Coué M, Tesse A, Falewée J, Aguesse A, Croyal M, Fizanne L, Chaigneau J, Boursier J, Ouguerram K. Spirulina Liquid Extract Protects against Fibrosis Related to Non-Alcoholic Steatohepatitis and Increases Ursodeoxycholic Acid. Nutrients. 2019; 11(1):194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010194
Chicago/Turabian StyleCoué, Marine, Angela Tesse, Juliette Falewée, Audrey Aguesse, Mikaël Croyal, Lionel Fizanne, Julien Chaigneau, Jérôme Boursier, and Khadija Ouguerram. 2019. "Spirulina Liquid Extract Protects against Fibrosis Related to Non-Alcoholic Steatohepatitis and Increases Ursodeoxycholic Acid" Nutrients 11, no. 1: 194. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010194