An Update on the Effects of Glyceollins on Human Health: Possible Anticancer Effects and Underlying Mechanisms


Abstract
:1. Introduction
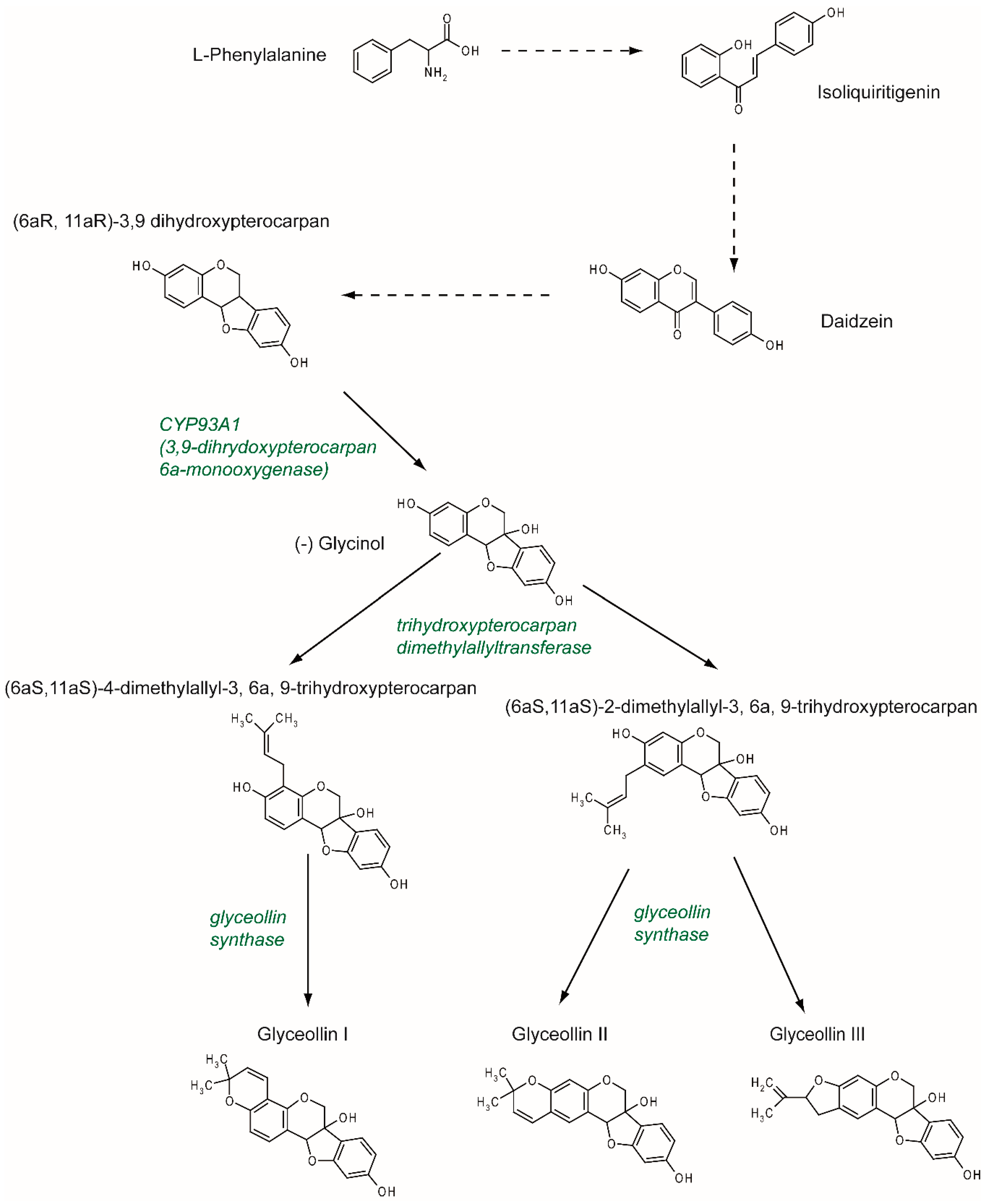
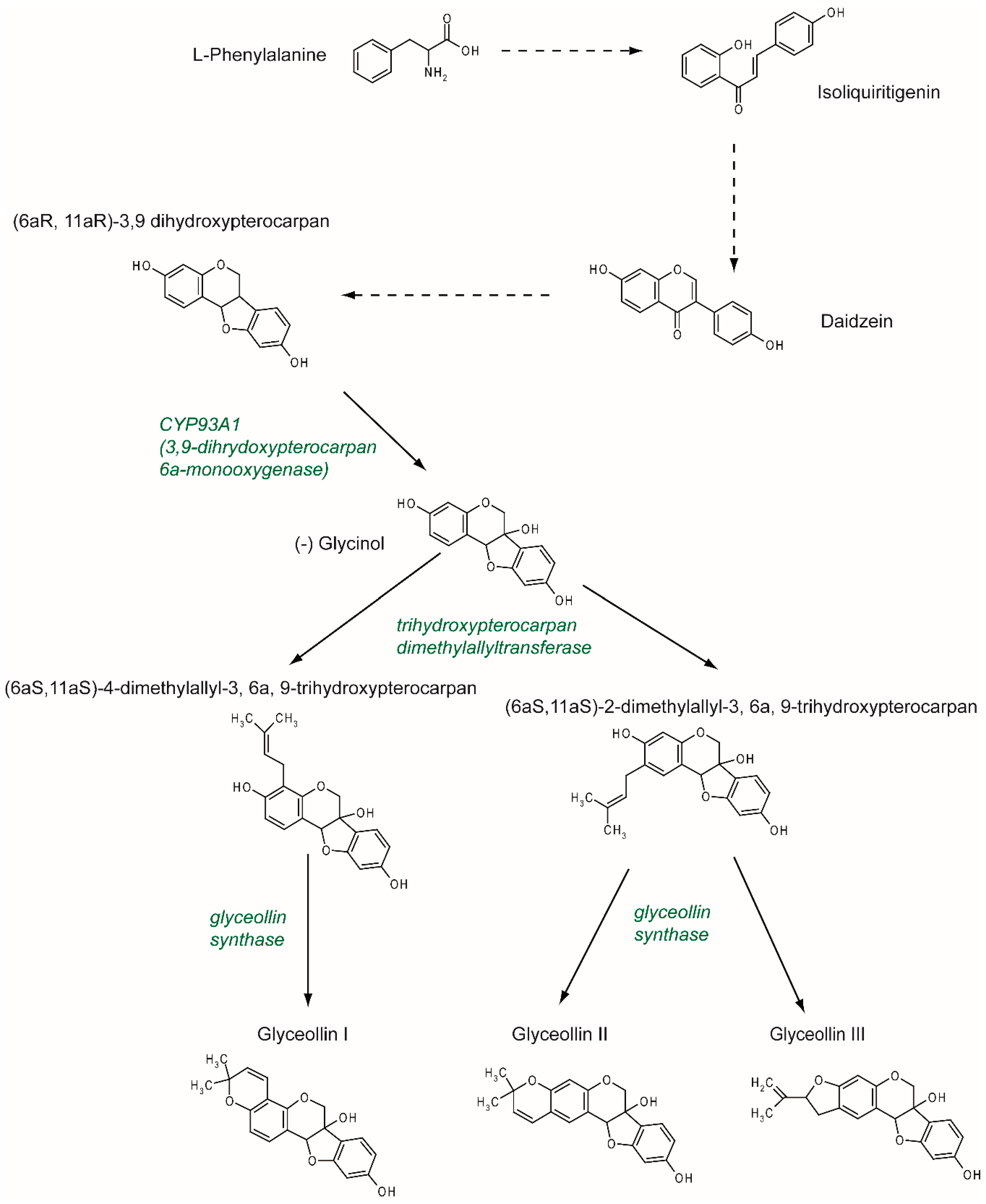
2. Synthesis and Structure
3. Metabolism and Pharmacokinetics
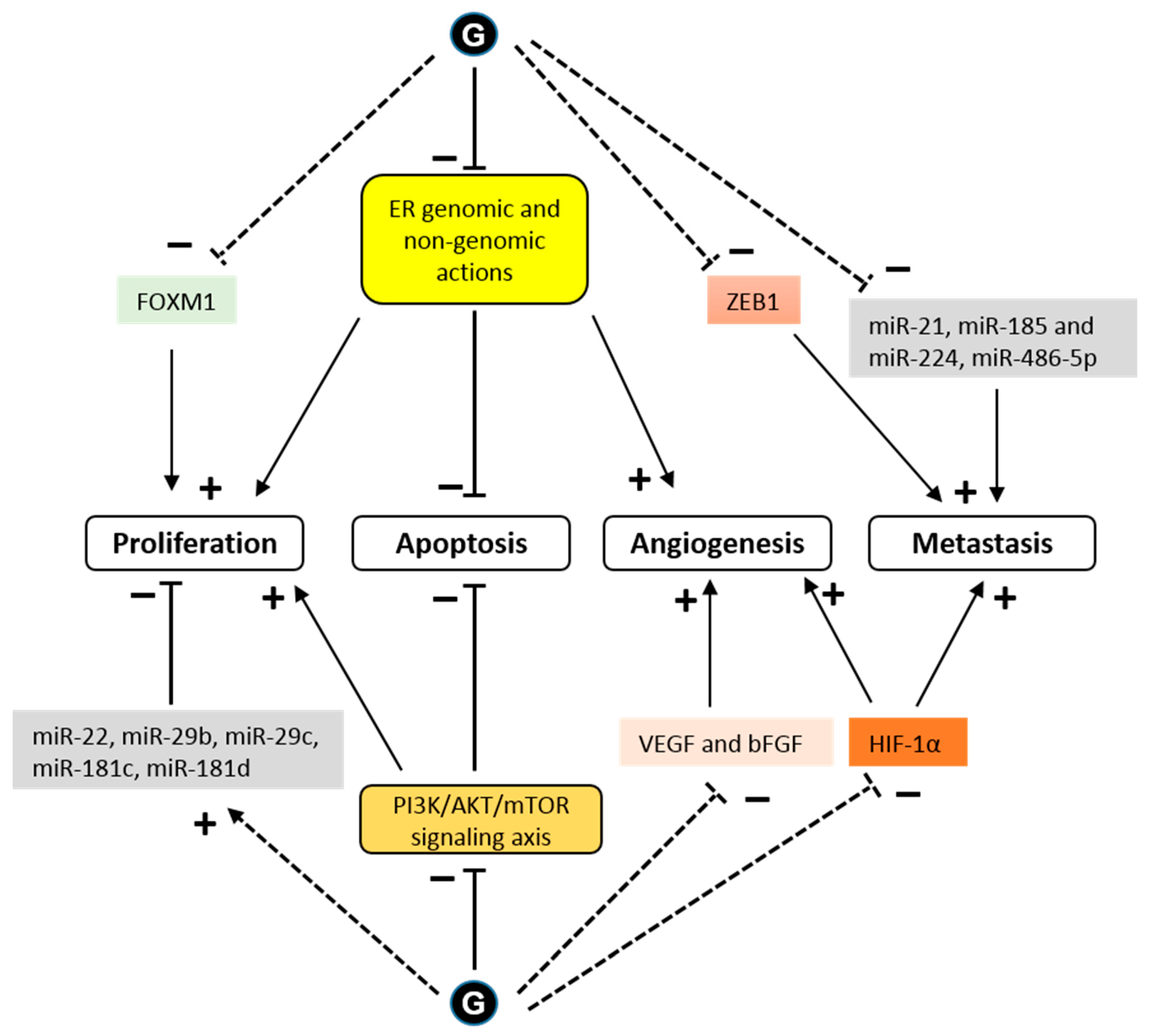
4. Anticancer Effects
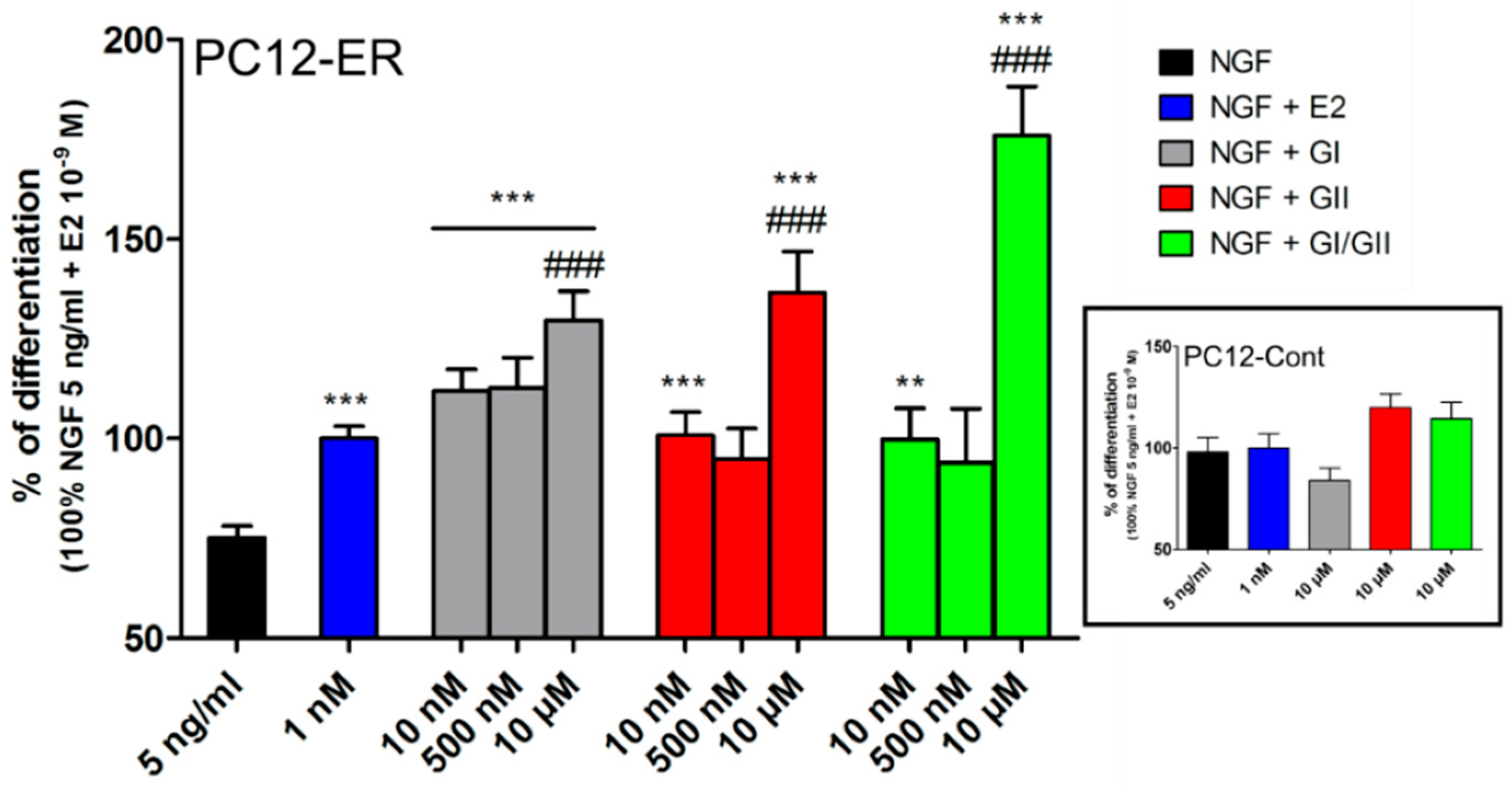
4.1. Estrogen-Dependent Effects
4.2. Estrogen-Independent Effect
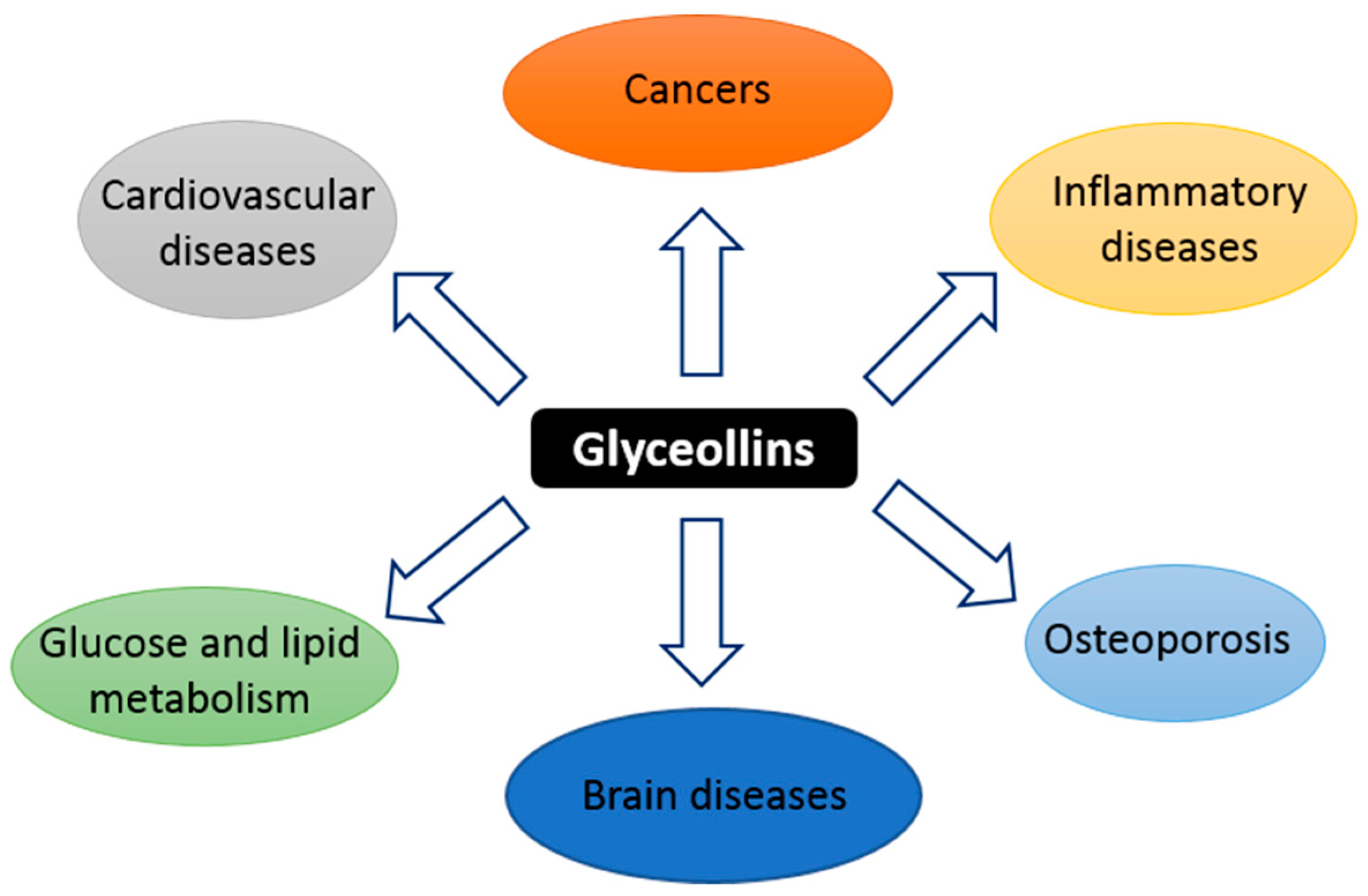
5. Other Effects in Noncancer Diseases
5.1. Osteoporosis
5.2. Glucose and Lipid Metabolism
5.3. Antioxidant Effect
5.4. Effect on the Central Nervous System
5.5. Anti-Inflammatory Activity and Cardiovascular Disease
5.6. Anti-Melanogenesis Activity
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, food and pharma. Current knowledge and directions for future research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Knaze, V.; Rothwell, J.A.; Zamora-Ros, R.; Moskal, A.; Kyrø, C.; Jakszyn, P.; Skeie, G.; Weiderpass, E.; Santucci de Magistris, M.; Agnoli, C.; et al. A new food-composition database for 437 polyphenols in 19,899 raw and prepared foods used to estimate polyphenol intakes in adults from 10 European countries. Am. J. Clin. Nutr. 2018, 108, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Maraldi, T.; Milenkovic, D.; Vauzour, D. Dietary Polyphenols and Their Effects on Cell Biochemistry and Pathophysiology 2014. Oxidative Med. Cell. Longev. 2015, 2015, 1–2. [Google Scholar] [CrossRef]
- Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Hennig, B.; Petriello, M.C.; Gamble, M.V.; Surh, Y.-J.; Kresty, L.A.; Frank, N.; Rangkadilok, N.; Ruchirawat, M.; Suk, W.A. The role of nutrition in influencing mechanisms involved in environmentally mediated diseases. Rev. Environ. Health 2018, 33, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Wongwarawipat, T.; Papageorgiou, N.; Bertsias, D.; Siasos, G.; Tousoulis, D. Olive Oil-related Anti-inflammatory Effects on Atherosclerosis: Potential Clinical Implications. Endocr. Metab. Immune Disord. Drug Targets 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Zujko, M.E.; Waśkiewicz, A.; Witkowska, A.M.; Szcześniewska, D.; Zdrojewski, T.; Kozakiewicz, K.; Drygas, W. Dietary Total Antioxidant Capacity and Dietary Polyphenol Intake and Prevalence of Metabolic Syndrome in Polish Adults: A Nationwide Study. Oxidative Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Goetz, M.E.; Judd, S.E.; Safford, M.M.; Hartman, T.J.; McClellan, W.M.; Vaccarino, V. Dietary flavonoid intake and incident coronary heart disease: the REasons for Geographic and Racial Differences in Stroke (REGARDS) study. Am. J. Clin. Nutr. 2016, 104, 1236–1244. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 384S–392S. [Google Scholar] [CrossRef]
- Anunciato, T.P.; da Rocha Filho, P.A. Carotenoids and polyphenols in nutricosmetics, nutraceuticals, and cosmeceuticals. J. Cosmet. Dermatol. 2012, 11, 51–54. [Google Scholar] [CrossRef]
- Pezzuto, J.M.; Kondratyuk, T.P.; Ogas, T. Resveratrol derivatives: a patent review (2009–2012). Expert Opin. Ther. Pat. 2013, 23, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Dragan, S.; Andrica, F.; Serban, M.-C.; Timar, R. Polyphenols-rich natural products for treatment of diabetes. Curr. Med. Chem. 2015, 22, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Ahiahonu, P.W.K. Metabolism and detoxification of phytoalexins and analogs by phytopathogenic fungi. Phytochemistry 2005, 66, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Dakora, F.D.; Phillips, D.A. Diverse functions of isoflavonoids in legumes transcend anti-microbial definitions of phytoalexins. Physiol. Mol. Plant Pathol. 1996, 49, 1–20. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Huffaker, A.; Sims, J.W.; Christensen, S.A.; Lu, X.; Okada, K.; Peters, R.J. Biosynthesis, elicitation and roles of monocot terpenoid phytoalexins. Plant J. 2014, 79, 659–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, Y.; Dang, P.; Zhao, S.; Lai, D.; Zhou, L. Rice Secondary Metabolites: Structures, Roles, Biosynthesis, and Metabolic Regulation. Molecules 2018, 23, 3098. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Kim, H.J.; Jeong, Y.-S.; Kim, W.-K.; Kim, J.-S. Differential abilities of Korean soybean varieties to biosynthesize glyceollins by biotic and abiotic elicitors. Food Sci. Biotechnol. 2017, 26, 255–261. [Google Scholar] [CrossRef]
- Block, A.K.; Vaughan, M.M.; Schmelz, E.A.; Christensen, S.A. Biosynthesis and function of terpenoid defense compounds in maize (Zea mays). Planta 2018. [Google Scholar] [CrossRef]
- Ejike, C.E.C.C.; Gong, M.; Udenigwe, C.C. Phytoalexins from the Poaceae: Biosynthesis, function and prospects in food preservation. Food Res. Int. 2013, 52, 167–177. [Google Scholar] [CrossRef]
- Hidalgo, W.; Chandran, J.N.; Menezes, R.C.; Otálvaro, F.; Schneider, B. Phenylphenalenones protect banana plants from infection by Mycosphaerella fijiensis and are deactivated by metabolic conversion. Plant Cell Environ. 2016, 39, 492–513. [Google Scholar] [CrossRef]
- Timperio, A.M.; D’Alessandro, A.; Fagioni, M.; Magro, P.; Zolla, L. Production of the phytoalexins trans-resveratrol and delta-viniferin in two economy-relevant grape cultivars upon infection with Botrytis cinerea in field conditions. Plant Physiol. Biochem. PPB 2012, 50, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Kim, J.Y.; Chun, J.; Park, S.; Kim, H.J.; Kim, J.-S.; Jeong, J.-I.; Kim, J.H. Induction of glyceollins by fungal infection in varieties of Korean soybean. J. Microbiol. Biotechnol. 2010, 20, 1226–1229. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lim, J.-S.; Kim, W.-K.; Kim, J.-S. Soyabean glyceollins: biological effects and relevance to human health. Proc. Nutr. Soc. 2012, 71, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Bamji, S.F.; Corbitt, C. Glyceollins: Soybean phytoalexins that exhibit a wide range of health-promoting effects. J. Funct. Foods 2017, 34, 98–105. [Google Scholar] [CrossRef]
- Lecomte, S.; Chalmel, F.; Ferriere, F.; Percevault, F.; Plu, N.; Saligaut, C.; Surel, C.; Lelong, M.; Efstathiou, T.; Pakdel, F. Glyceollins trigger anti-proliferative effects through estradiol-dependent and independent pathways in breast cancer cells. Cell Commun. Signal. CCS 2017, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Salvo, V.A.; Boue, S.M.; Fonseca, J.P.; Elliott, S.; Corbitt, C.; Collins-Burow, B.M.; Curiel, T.J.; Srivastav, S.K.; Shih, B.Y.; Carter-Wientjes, C.; et al. Antiestrogenic Glyceollins Suppress Human Breast and Ovarian Carcinoma Tumorigenesis. Clin. Cancer Res. 2006, 12, 7159–7164. [Google Scholar] [CrossRef] [Green Version]
- Burow, M.E.; Boue, S.M.; Collins-Burow, B.M.; Melnik, L.I.; Duong, B.N.; Carter-Wientjes, C.H.; Li, S.; Wiese, T.E.; Cleveland, T.E.; McLachlan, J.A. Phytochemical Glyceollins, Isolated from Soy, Mediate Antihormonal Effects through Estrogen Receptor α and β. J. Clin. Endocrinol. Metab. 2001, 86, 1750–1758. [Google Scholar] [PubMed]
- Zimmermann, M.C.; Tilghman, S.L.; Boue, S.M.; Salvo, V.A.; Elliott, S.; Williams, K.Y.; Skripnikova, E.V.; Ashe, H.; Payton-Stewart, F.; Vanhoy-Rhodes, L.; et al. Glyceollin I, a Novel Antiestrogenic Phytoalexin Isolated from Activated Soy. J. Pharmacol. Exp. Ther. 2010, 332, 35–45. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.; Jung, M.H.; Lee, Y.M. Glyceollins, a novel class of soy phytoalexins, inhibit angiogenesis by blocking the VEGF and bFGF signaling pathways. Mol. Nutr. Food Res. 2013, 57, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Jee, J.-G.; Bae, J.-S.; Liu, K.-H.; Lee, Y.M. A Group of Novel HIF-1α Inhibitors, Glyceollins, Blocks HIF-1α Synthesis and Decreases Its Stability via Inhibition of the PI3K/AKT/mTOR Pathway and Hsp90 Binding: GLYCEOLLINS INHIBIT HIF-1α. J. Cell. Physiol. 2015, 230, 853–862. [Google Scholar] [CrossRef]
- Carriere, P.P.; Llopis, S.D.; Naiki, A.C.; Nguyen, G.; Phan, T.; Nguyen, M.M.; Preyan, L.C.; Yearby, L.; Pratt, J.; Burks, H.; et al. Glyceollin I Reverses Epithelial to Mesenchymal Transition in Letrozole Resistant Breast Cancer through ZEB1. Int. J. Environ. Res. Public. Health 2016, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Burden, R.S.; Bailey, J.A. Structure of the phytoalexin from soybean. Phytochemistry 1975, 14, 1389–1390. [Google Scholar] [CrossRef]
- Keen, N.T.; Zaki, A.I.; Sims, J.J. Biosynthesis of hydroxyphaseollin and related isoflavanoids in disease-resistant soybean hypocotyls. Phytochemistry 1972, 11, 1031–1039. [Google Scholar] [CrossRef]
- Schopfer, C.R.; Kochs, G.; Lottspeich, F.; Ebel, J. Molecular characterization and functional expression of dihydroxypterocarpan 6a-hydroxylase, an enzyme specific for pterocarpanoid phytoalexin biosynthesis in soybean (Glycine max L.). FEBS Lett. 1998, 432, 182–186. [Google Scholar] [CrossRef]
- Yoneyama, K.; Akashi, T.; Aoki, T. Molecular Characterization of Soybean Pterocarpan 2-Dimethylallyltransferase in Glyceollin Biosynthesis: Local Gene and Whole-Genome Duplications of Prenyltransferase Genes Led to the Structural Diversity of Soybean Prenylated Isoflavonoids. Plant Cell Physiol. 2016, 57, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Akashi, T.; Sasaki, K.; Aoki, T.; Ayabe, S.; Yazaki, K. Molecular cloning and characterization of a cDNA for pterocarpan 4-dimethylallyltransferase catalyzing the key prenylation step in the biosynthesis of glyceollin, a soybean phytoalexin. Plant Physiol. 2009, 149, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, A.; McDowell, T.; Chen, L.; Renaud, J.; Dhaubhadel, S. Isoflavonoid-specific prenyltransferase gene family in soybean: GmPT01, a pterocarpan 2-dimethylallyltransferase involved in glyceollin biosynthesis. Plant J. Cell Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Welle, R.; Grisebach, H. Induction of phytoalexin synthesis in soybean: enzymatic cyclization of prenylated pterocarpans to glyceollin isomers. Arch. Biochem. Biophys. 1988, 263, 191–198. [Google Scholar] [CrossRef]
- Farrell, K.; Jahan, M.A.; Kovinich, N. Distinct Mechanisms of Biotic and Chemical Elicitors Enable Additive Elicitation of the Anticancer Phytoalexin Glyceollin I. Molecules 2017, 22, 1261. [Google Scholar] [CrossRef]
- Wood, C.E.; Clarkson, T.B.; Appt, S.E.; Franke, A.A.; Boue, S.M.; Burow, M.E.; McCoy, T.; Cline, J.M. Effects of Soybean Glyceollins and Estradiol in Postmenopausal Female Monkeys. Nutr. Cancer 2006, 56, 74–81. [Google Scholar] [CrossRef]
- Boué, S.M.; Isakova, I.A.; Burow, M.E.; Cao, H.; Bhatnagar, D.; Sarver, J.G.; Shinde, K.V.; Erhardt, P.W.; Heiman, M.L. Glyceollins, Soy Isoflavone Phytoalexins, Improve Oral Glucose Disposal by Stimulating Glucose Uptake. J. Agric. Food Chem. 2012, 60, 6376–6382. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kim, J.H.; Kim, J.S.; Kim, H.J. Glyceollin-containing fermented soybeans improve glucose homeostasis in diabetic mice. Nutrition 2012, 28, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Quadri, S.S.; Stratford, R.E.; Boué, S.M.; Cole, R.B. Screening and Identification of Glyceollins and Their Metabolites by Electrospray Ionization Tandem Mass Spectrometry with Precursor Ion Scanning. Anal. Chem. 2013, 85, 1727–1733. [Google Scholar] [CrossRef] [PubMed]
- Quadri, S.S.; Stratford, R.E.; Boué, S.M.; Cole, R.B. Identification of Glyceollin Metabolites Derived from Conjugation with Glutathione and Glucuronic Acid in Male ZDSD Rats by Online Liquid Chromatography–Electrospray Ionization Tandem Mass Spectrometry. J. Agric. Food Chem. 2014, 62, 2692–2700. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, M. Absorption and metabolism of flavonoids in the caco-2 cell culture model and a perused rat intestinal model. Drug Metab. Dispos. Biol. Fate Chem. 2002, 30, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Radic. Res. 2004, 38, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Andreadi, C.; Britton, R.G.; Horner-Glister, E.; Karmokar, A.; Sale, S.; Brown, V.A.; Brenner, D.E.; Singh, R.; Steward, W.P.; et al. Sulfate metabolites provide an intracellular pool for resveratrol generation and induce autophagy with senescence. Sci. Transl. Med. 2013, 5, 205ra133. [Google Scholar] [CrossRef] [PubMed]
- Chimezie, C.; Ewing, A.C.; Quadri, S.S.; Cole, R.B.; Boué, S.M.; Omari, C.F.; Bratton, M.; Glotser, E.; Skripnikova, E.; Townley, I.; et al. Glyceollin Transport, Metabolism, and Effects on P-Glycoprotein Function in Caco-2 Cells. J. Med. Food 2014, 17, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Chimezie, C.; Ewing, A.; Schexnayder, C.; Bratton, M.; Glotser, E.; Skripnikova, E.; Sá, P.; Boué, S.; Stratford, R.E. Glyceollin Effects on MRP2 and BCRP in Caco-2 Cells, and Implications for Metabolic and Transport Interactions. J. Pharm. Sci. 2016, 105, 972–981. [Google Scholar] [CrossRef] [Green Version]
- Boué, S.; Fortgang, I.; Levy, R.J.; Bhatnagar, D.; Burow, M.; Fahey, G.; Heiman, M.L. A novel gastrointestinal microbiome modulator from soy pods reduces absorption of dietary fat in mice. Obesity 2016, 24, 87–95. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012: Globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xiang, L.; Li, T.; Bai, Z. Cancer Hallmarks, Biomarkers and Breast Cancer Molecular Subtypes. J. Cancer 2016, 7, 1281–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
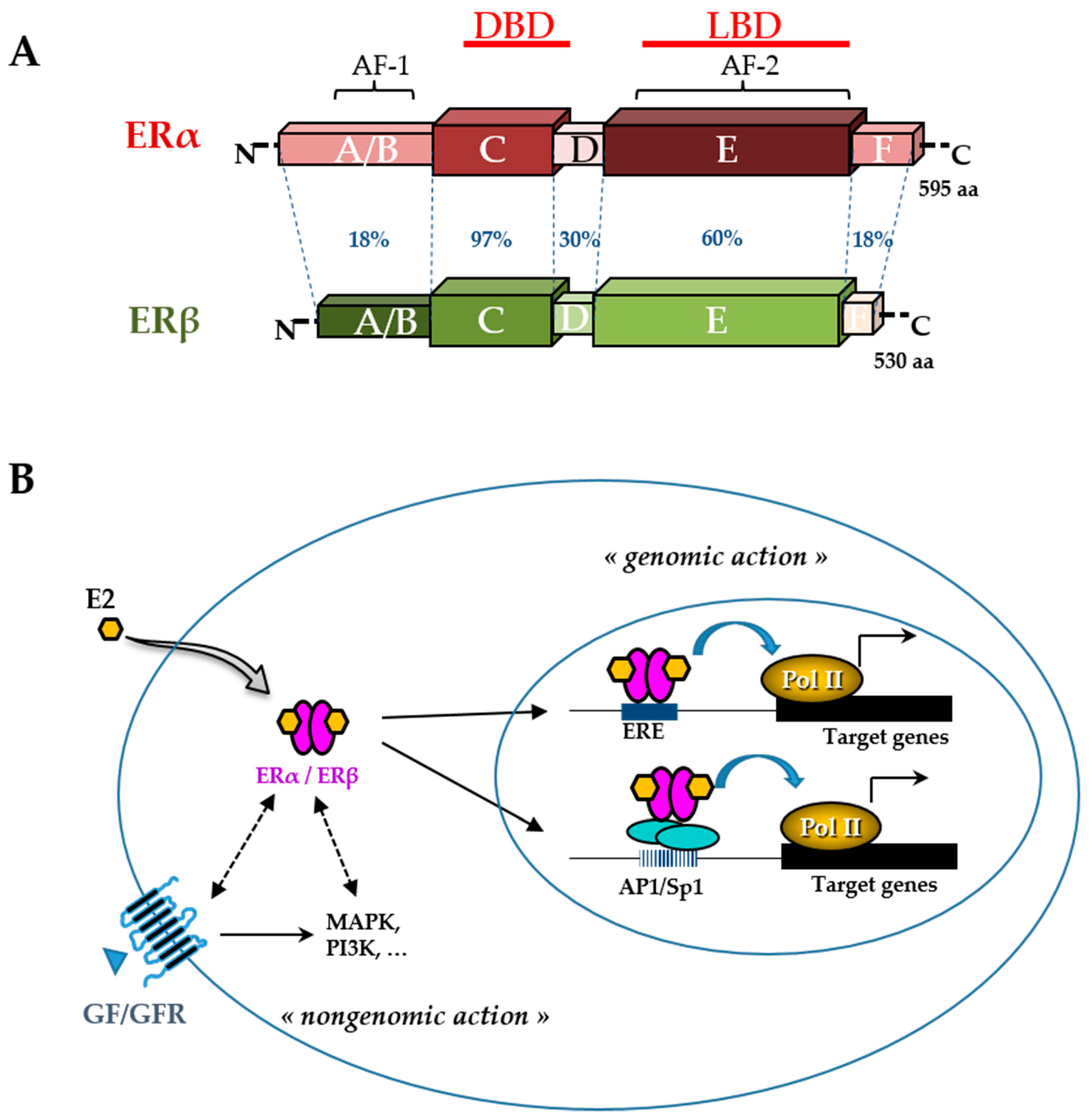
- Pakdel, F. Molecular Pathways of Estrogen Receptor Action. Int. J. Mol. Sci. 2018, 19, 2591. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. Estrogen receptor interaction with co-activators and co-repressors. Steroids 2000, 65, 227–251. [Google Scholar] [CrossRef]
- Safe, S.; Kim, K. Nuclear Receptor-Mediated Transactivation through Interaction with Sp Proteins. In Progress in Nucleic Acid Research and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2004; Volume 77, pp. 1–36. ISBN 978-0-12-540077-0. [Google Scholar]
- Mehta, R.G.; Hawthorne, M.; Mehta, R.R.; Torres, K.E.O.; Peng, X.; McCormick, D.L.; Kopelovich, L. Differential Roles of ERα and ERβ in Normal and Neoplastic Development in the Mouse Mammary Gland. PLoS ONE 2014, 9, e113175. [Google Scholar] [CrossRef] [PubMed]
- Payton-Stewart, F.; Khupse, R.S.; Boué, S.M.; Elliott, S.; Zimmermann, M.C.; Skripnikova, E.V.; Ashe, H.; Tilghman, S.L.; Beckman, B.S.; Cleveland, T.E.; et al. Glyceollin I enantiomers distinctly regulate ER-mediated gene expression. Steroids 2010, 75, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Boué, S.M.; Tilghman, S.L.; Elliott, S.; Zimmerman, M.C.; Williams, K.Y.; Payton-Stewart, F.; Miraflor, A.P.; Howell, M.H.; Shih, B.Y.; Carter-Wientjes, C.H.; et al. Identification of the Potent Phytoestrogen Glycinol in Elicited Soybean (Glycine max). Endocrinology 2009, 150, 2446–2453. [Google Scholar] [CrossRef]
- Bratton, M.R.; Martin, E.C.; Elliott, S.; Rhodes, L.V.; Collins-Burow, B.M.; McLachlan, J.A.; Wiese, T.E.; Boue, S.M.; Burow, M.E. Glyceollin, a novel regulator of mTOR/p70S6 in estrogen receptor positive breast cancer. J. Steroid Biochem. Mol. Biol. 2015, 150, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Fingar, D.C.; Blenis, J. Target of rapamycin (TOR): an integrator of nutrient and growth factor signals and coordinator of cell growth and cell cycle progression. Oncogene 2004, 23, 3151–3171. [Google Scholar] [CrossRef] [Green Version]
- Brown, K. Breast cancer chemoprevention: risk-benefit effects of the antioestrogen tamoxifen. Expert Opin. Drug Saf. 2002, 1, 253–267. [Google Scholar] [CrossRef]
- Payton-Stewart, F.; Schoene, N.W.; Kim, Y.S.; Burow, M.E.; Cleveland, T.E.; Boue, S.M.; Wang, T.T.Y. Molecular effects of soy phytoalexin glyceollins in human prostate cancer cells LNCaP. Mol. Carcinog. 2009, 48, 862–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, L.V.; Tilghman, S.L.; Boue, S.M.; Wang, S.; Khalili, H.; Muir, S.E.; Bratton, M.R.; Zhang, Q.; Wang, G.; Burow, M.E.; et al. Glyceollins as novel targeted therapeutic for the treatment of triple-negative breast cancer. Oncol. Lett. 2012, 3, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Guarino, M.; Rubino, B.; Ballabio, G. The role of epithelial-mesenchymal transition in cancer pathology. Pathology 2007, 39, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Schmalhofer, O.; Brabletz, S.; Brabletz, T. E-cadherin, beta-catenin, and ZEB1 in malignant progression of cancer. Cancer Metastasis Rev. 2009, 28, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G.; Ratcliffe, P.J. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Isaacs, J.S.; Jung, Y.-J.; Mimnaugh, E.G.; Martinez, A.; Cuttitta, F.; Neckers, L.M. Hsp90 regulates a von Hippel Lindau-independent hypoxia-inducible factor-1 alpha-degradative pathway. J. Biol. Chem. 2002, 277, 29936–29944. [Google Scholar] [CrossRef]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef]
- Kim, H.J.; Jung, C.L.; Jeong, Y.S.; Kim, J.-S. Soybean-derived glyceollins induce apoptosis through ROS generation. Food Funct. 2014, 5, 688–695. [Google Scholar] [CrossRef]
- Kim, B.R.; Seo, J.-Y.; Sung, M.-K.; Park, J.H.Y.; Suh, H.-J.; Liu, K.H.; Kim, J.-S. Suppression of 7,12-dimethylbenz(a)anthracene-induced mammary tumorigenesis by glyceollins. Mol. Nutr. Food Res. 2015, 59, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.K.; Hampson, G. The pathogenesis, diagnosis, investigation and management of osteoporosis. J. Clin. Pathol. 2011, 64, 1042–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghebati-Maleki, L.; Dolati, S.; Zandi, R.; Fotouhi, A.; Ahmadi, M.; Aghebati, A.; Nouri, M.; Shakouri, S.K.; Yousefi, M. Prospect of mesenchymal stem cells in therapy of osteoporosis: A review. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bateman, M.E.; Strong, A.L.; Hunter, R.S.; Bratton, M.R.; Komati, R.; Sridhar, J.; Riley, K.E.; Wang, G.; Hayes, D.J.; Boue, S.M.; et al. Osteoinductive effects of glyceollins on adult mesenchymal stromal/stem cells from adipose tissue and bone marrow. Phytomed. Int. J. Phytother. Phytopharm. 2017, 27, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Aragaki, A.K.; Rossouw, J.E.; Anderson, G.L.; Prentice, R.L.; LaCroix, A.Z.; Chlebowski, R.T.; Howard, B.V.; Thomson, C.A.; Margolis, K.L.; et al. Menopausal Hormone Therapy and Long-term All-Cause and Cause-Specific Mortality: The Women’s Health Initiative Randomized Trials. JAMA 2017, 318, 927–938. [Google Scholar] [CrossRef]
- De Wilde, A.; Lieberherr, M.; Colin, C.; Pointillart, A. A low dose of daidzein acts as an ERbeta-selective agonist in trabecular osteoblasts of young female piglets. J. Cell. Physiol. 2004, 200, 253–262. [Google Scholar] [CrossRef]
- Park, S.; Ahn, I.S.; Kim, J.H.; Lee, M.R.; Kim, J.S.; Kim, H.J. Glyceollins, one of the phytoalexins derived from soybeans under fungal stress, enhance insulin sensitivity and exert insulinotropic actions. J. Agric. Food Chem. 2010, 58, 1551–1557. [Google Scholar] [CrossRef]
- Huang, H.; Xie, Z.; Boue, S.M.; Bhatnagar, D.; Yokoyama, W.; Yu, L.L.; Wang, T.T.Y. Cholesterol-lowering activity of soy-derived glyceollins in the golden Syrian hamster model. J. Agric. Food Chem. 2013, 61, 5772–5782. [Google Scholar] [CrossRef]
- Wood, C.E.; Boue, S.M.; Collins-Burow, B.M.; Rhodes, L.V.; Register, T.C.; Cline, J.M.; Dewi, F.N.; Burow, M.E. Glyceollin-elicited soy protein consumption induces distinct transcriptional effects as compared to standard soy protein. J. Agric. Food Chem. 2012, 60, 81–86. [Google Scholar] [CrossRef]
- Kim, H.J.; Suh, H.-J.; Kim, J.H.; Park, S.; Joo, Y.C.; Kim, J.-S. Antioxidant activity of glyceollins derived from soybean elicited with Aspergillus sojae. J. Agric. Food Chem. 2010, 58, 11633–11638. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; di Luccio, E.; Kong, A.-N.T.; Kim, J.-S. Nrf2-mediated induction of phase 2 detoxifying enzymes by glyceollins derived from soybean exposed to Aspergillus sojae. Biotechnol. J. 2011, 6, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.L.; Kim, H.J.; Park, J.H.Y.; Kong, A.-N.T.; Lee, C.H.; Kim, J.-S. Synergistic activation of the Nrf2-signaling pathway by glyceollins under oxidative stress induced by glutathione depletion. J. Agric. Food Chem. 2013, 61, 4072–4078. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.Y.; Kim, B.R.; Oh, J.; Kim, J.-S. Soybean-Derived Phytoalexins Improve Cognitive Function through Activation of Nrf2/HO-1 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 268. [Google Scholar] [CrossRef] [PubMed]
- Bamji, S.F.; Page, R.B.; Patel, D.; Sanders, A.; Alvarez, A.R.; Gambrell, C.; Naik, K.; Raghavan, A.M.; Burow, M.E.; Boue, S.M.; et al. Soy glyceollins regulate transcript abundance in the female mouse brain. Funct. Integr. Genom. 2015, 15, 549–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamji, S.F.; Rouchka, E.; Zhang, Y.; Li, X.; Kalbfleisch, T.; Corbitt, C. Next generation sequencing analysis of soy glyceollins and 17-β estradiol: Effects on transcript abundance in the female mouse brain. Mol. Cell. Endocrinol. 2018, 471, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Sung, M.-K.; Kim, J.-S. Anti-inflammatory effects of glyceollins derived from soybean by elicitation with Aspergillus sojae. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. Al 2011, 60, 909–917. [Google Scholar] [CrossRef]
- Yoon, E.-K.; Kim, H.-K.; Cui, S.; Kim, Y.-H.; Lee, S.-H. Soybean glyceollins mitigate inducible nitric oxide synthase and cyclooxygenase-2 expression levels via suppression of the NF-κB signaling pathway in RAW 264.7 cells. Int. J. Mol. Med. 2012, 29, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Ku, S.-K.; Lee, Y.-M.; Bae, J.-S. Anti-septic effects of glyceollins in HMGB1-induced inflammatory responses in vitro and in vivo. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 63, 1–8. [Google Scholar] [CrossRef]
- Kim, H.J.; Cha, B.-Y.; Choi, B.; Lim, J.S.; Woo, J.-T.; Kim, J.-S. Glyceollins inhibit platelet-derived growth factor-mediated human arterial smooth muscle cell proliferation and migration. Br. J. Nutr. 2012, 107, 24–35. [Google Scholar] [CrossRef]
- Song, M.-J.; Baek, I.; Jeon, S.B.; Seo, M.; Kim, Y.-H.; Cui, S.; Jeong, Y.-S.; Lee, I.-J.; Shin, D.H.; Hwang, Y.H.; et al. Effects of glyceollin I on vascular contraction in rat aorta. Naunyn-Schmiedebergs Arch. Pharmacol. 2010, 381, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Kim, H.-K.; Lee, K.J.; Jeon, H.W.; Cui, S.; Lee, Y.M.; Moon, B.J.; Kim, Y.-H.; Lee, Y.-S. Inhibitory effect of glyceollin isolated from soybean against melanogenesis in B16 melanoma cells. BMB Rep. 2010, 43, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.-H.; Lee, Y.-M. Glyceollins, a novel class of soybean phytoalexins, inhibit SCF-induced melanogenesis through attenuation of SCF/c-kit downstream signaling pathways. Exp. Mol. Med. 2013, 45, e17. [Google Scholar] [CrossRef] [PubMed]
- Matschke, V.; Theiss, C.; Matschke, J. Oxidative stress: the lowest common denominator of multiple diseases. Neural Regen. Res. 2019, 14, 238–241. [Google Scholar] [CrossRef]
- Cacciapuoti, F. Oxidative Stress as “Mother” of Many Human Diseases at Strong Clinical Impact. J. Cardiovasc. Med. Cardiol. 2016, 3, 001–006. [Google Scholar] [CrossRef]
- Tang, Y.; Tsao, R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory, and potential health beneficial effects: A review. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Alasalvar, C.; Bolling, B.W. Review of nut phytochemicals, fat-soluble bioactives, antioxidant components and health effects. Br. J. Nutr. 2015, 113 (Suppl. 2), S68–S78. [Google Scholar] [CrossRef] [Green Version]
- Baranowska, M.; Bartoszek, A. Antioxidant and antimicrobial properties of bioactive phytochemicals from cranberry. Postepy Hig. Med. Dosw. (Online) 2016, 70, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. A Critical Review on Polyphenols and Health Benefits of Black Soybeans. Nutrients 2017, 9, 455. [Google Scholar] [CrossRef]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: a key player for human disease. Cell Death Dis. 2013, 4, e888. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Kudo, Y.; Aoyagi, K.; Ishikawa, R.; Igarashi, M.; Takahashi, M. Calmodulin-dependent regulation of neurotransmitter release differs in subsets of neuronal cells. Brain Res. 2013, 1535, 1–13. [Google Scholar] [CrossRef] [PubMed]
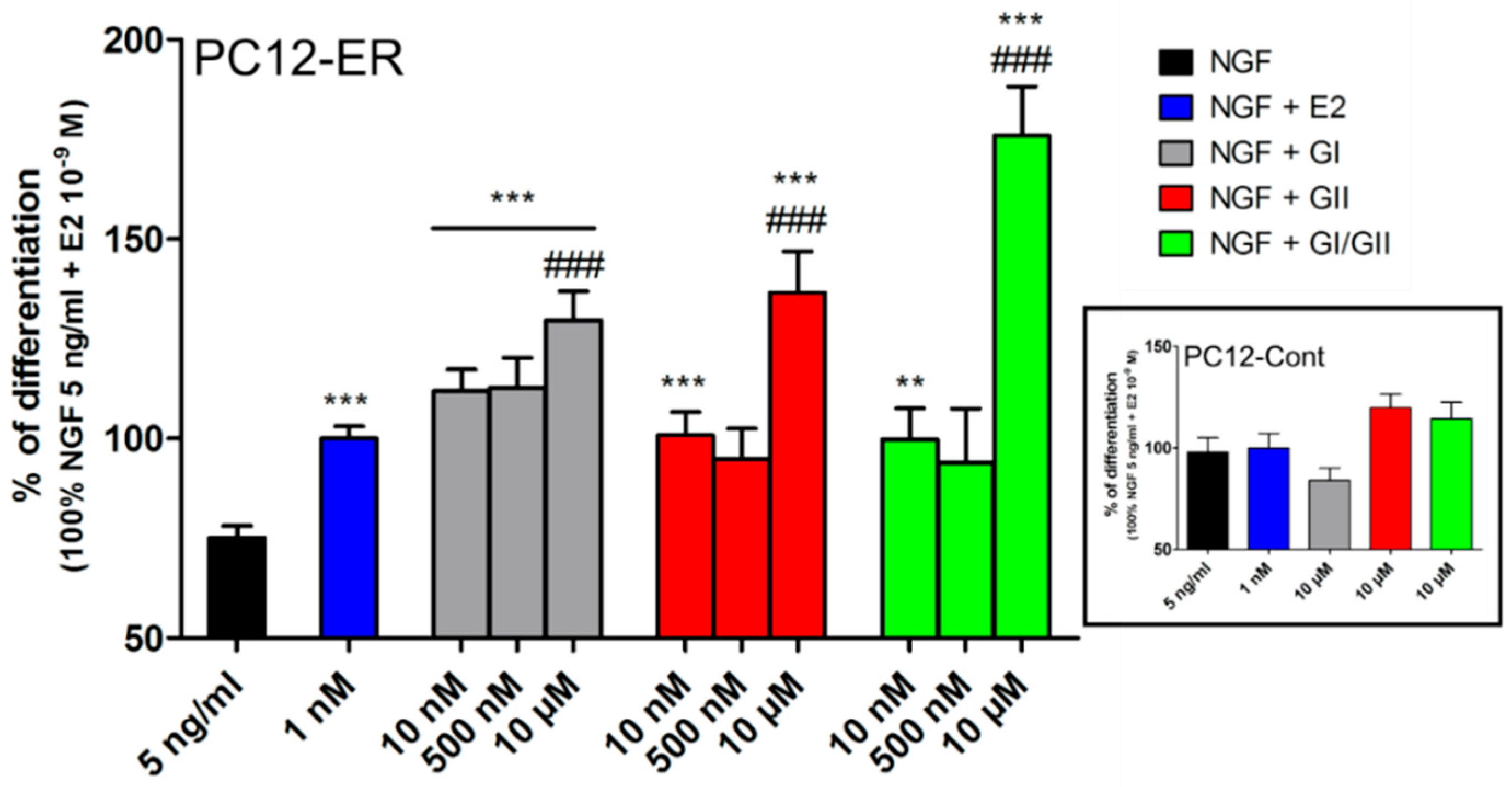
- Ferriere, F.; Habauzit, D.; Pakdel, F.; Saligaut, C.; Flouriot, G. Unliganded Estrogen Receptor Alpha Promotes PC12 Survival during Serum Starvation. PLoS ONE 2013, 8, e69081. [Google Scholar] [CrossRef] [PubMed]
- Habauzit, D.; Ferrière, F.; Botherel, N.; Flouriot, G.; Pakdel, F.; Saligaut, C. Differentiation of PC12 cells expressing estrogen receptor alpha: A new bioassay for endocrine-disrupting chemicals evaluation. Chemosphere 2014, 112, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecomte, S.; Lelong, M.; Bourgine, G.; Efstathiou, T.; Saligaut, C.; Pakdel, F. Assessment of the potential activity of major dietary compounds as selective estrogen receptor modulators in two distinct cell models for proliferation and differentiation. Toxicol. Appl. Pharmacol. 2017, 325, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero-Miliani, L.; Nielsen, O.H.; Andersen, P.S.; Girardin, S.E. Chronic inflammation: importance of NOD2 and NALP3 in interleukin-1β generation. Clin. Exp. Immunol. 2007, 147, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Sailo, B.L.; Banik, K.; Harsha, C.; Prasad, S.; Gupta, S.C.; Bharti, A.C.; Aggarwal, B.B. Chronic diseases, inflammation, and spices: how are they linked? J. Transl. Med. 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Strohacker, K.; McFarlin, B.K. Influence of obesity, physical inactivity, and weight cycling on chronic inflammation. Front. Biosci. Elite Ed. 2010, 2, 98–104. [Google Scholar]
- Pellegrini, L.; Foglio, E.; Pontemezzo, E.; Germani, A.; Russo, M.A.; Limana, F. HMGB1 and repair: focus on the heart. Pharmacol. Ther. 2018. [Google Scholar] [CrossRef]
- Shimokawa, H.; Sunamura, S.; Satoh, K. RhoA/Rho-Kinase in the Cardiovascular System. Circ. Res. 2016, 118, 352–366. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Huang, Y. Targeting the platelet-derived growth factor signalling in cardiovascular disease. Clin. Exp. Pharmacol. Physiol. 2015, 42, 1221–1224. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Downregulation of melanogenesis: drug discovery and therapeutic options. Drug Discov. Today 2017, 22, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Le Marchand, L. Cancer preventive effects of flavonoids—A review. Biomed. Pharmacother. Biomed. Pharmacother. 2002, 56, 296–301. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Van de Schans, M.G.M.; Vincken, J.-P.; de Waard, P.; Hamers, A.R.M.; Bovee, T.F.H.; Gruppen, H. Glyceollins and dehydroglyceollins isolated from soybean act as SERMs and ER subtype-selective phytoestrogens. J. Steroid Biochem. Mol. Biol. 2016, 156, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Vadhanam, M.V. Bioavailability of phytochemicals and its enhancement by drug delivery systems. Cancer Lett. 2013, 334, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walle, T. Bioavailability of resveratrol: Resveratrol bioavailability. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Model | Treatment | Sample | Time | Method of Measure | Major Results | References |
|---|---|---|---|---|---|---|
| Female monkey (Macaca fascicularis) | Diet containing glyceollin mixture 1, 134 mg/day representing 50% of total isoflavonoids in the diet | Plasma | 4 h and 24 h post administration (postad) | Liquid chromatographic-photodiode array mass spectrometric analysis | Plasma concentration of glyceollins: - 4 h: 134.2 ± 34.6 nmol/L, representing only 11.6 % of the plasma isoflavonoids level - 24 h: Undetectable ˂ 1 nmol/L | Wood et al. [40] |
| Male ZDSD/Pco rat | Glyceollin mixture, gavage, 30 and 90 mg/kg | Plasma | 20, 60, 120 and 240 minutes (min) postad | HPLC-electrospray ionization-MS/MS | Plama concentration of glyceollins: - 20 min: Starts to be detectable - the next 220 min: Remains stable For dose 30 mg/kg: Ranges from 81.2 to 118.4 ng/mL For dose 60 mg/kg: Ranges from 118.2 to 159.0 ng/mL - 60 min: Peak concentration | Boué et al. 2012 [41] |
| Glyceollin mixture, gavage, 90 mg/kg/days for 2 weeks | Plasma, feces, and urine | Plasma: 3 h postad Feces: Once daily for 2 weeks Urine: 24 h collection postad a single dose | Precursor and product ion scanning using liquid chromatography coupled online with Electrospray ionization tandem mass spectrometry | - Rapidly absorption, glyceollins undergo phase I and phase II metabolism in the small intestine and the liver - Metabolites of glyceollins were identified in the plasma, the urine, and the feces Phase I conjugation: Epoxidation, hydroxylation… Phase II conjugation: Sulfate and glucuronide conjugations… | Quadri et al. 2013 [43], Quadri et al. 2014 [44] |
| Diseases or Functions | Cell Line/Animal Model | Treatment | Major Effects | References |
|---|---|---|---|---|
| Osteoporosis | Adipose-derived stromal cells and bone marrow stromal cell line | GI 1 and GII 2 | - GI and GII: Increase calcium deposition - GII: Stronger than E2 and GI, increase the expression of genes involved in osteogenic differentiation in a similar manner to E2 | Bateman et al. 2017 [76] |
| Glucose and lipid metabolism | Enteroendocrine NCI-H716 cell line | Glyceollin mixture 3 | - Enhance GLP-1 4 secretion to increase insulinotropic actions | Park et al. 2010 [79] |
| 3T3-L1 adipocyte cell line | Glyceollin mixture | - Increase both insulin-stimulated and basal glucose uptake - Increase glucose transporter GLUT4 5 level - Decrease triacylglycerol accumulation | Boué et al. 2012 [41], Park et al. 2010 [79] | |
| Insulima Min6 cell line | Glyceollin mixture | - Decrease apoptosis - Enhance insulinotropic actions | Park et al. 2010 [79] | |
| Diabetic male C57BL6J mouse | Fermented soybeans containing glyceollins | - Decrease blood glucose level - Increase hepatic glycogen accumulation - Decrease triglyceride storage | Park et al. 2012 [42] | |
| Prediabetic male ZDSD/Pco rat | Glyceollin mixture | - Decrease blood glucose level | Boué et al. 2012 [41] | |
| Male golden Syrian hamster | Diet containing glyceollins | - Reduce plasma VLDL 6, hepatic cholesterol esters, total lipid content - Alter expression of the genes related to cholesterol in liver | Huang et al. 2013 [80] | |
| Female monkey (Macaca fascicularis) | Diet containing glyceollins | - Decrease serum total cholesterol, specifically LDL 7 and VLDL, increase serum triglycerides - Upregulate genes expression of PPAR 8-γ, adiponectin, leptin, liptin 1, lipoprotein lipase and triglyceride | Wood et al. 2012 [81] | |
| Oxidative stress | Tests in vitro | Glyceollin mixture | - Have ferric-reducing antioxidant power, radical scavenging activities | Kim et al. 2010 [82] |
| Hepa1c1c7/ BPRc1 cell line | Glyceollin mixture | - Inhibit H2O2-induced ROS 9 production - Induce Nrf2-mediated phase 2 detoxifying enzymes - Activate Nrf2-signaling pathway under oxidative stress | Kim et al. 2010 [82], Kim et al. 2011 [83] Jung et al. 2013 [84] | |
| Female C57BL/6J mouse | Glyceollin mixture | - Increase NAD(P)H:quinone oxidoreductase 1activity in kidney, liver, and large intestine | Kim et al. 2015 [72] | |
| Rat | Glyceollin mixture | - Inhibit lipid peroxidation in liver, kidney, and brain tissue extracts | Kim et al. 2010 [82] | |
| Central nervous system | Breast cancer MCF-7 cell line | Glyceollins mixture | - Increase nerve growth factor receptor gene expression | Zimmermann et al. 2010 [28] |
| Glutamate-sensitive murine hippocampal HT22 cell line | Glyceollin mixture | - Attenuate glutamate-induced neurotoxicity - Suppress glutamate-induced intracellular ROS - Act through Nrf2 signaling pathway, activate hem oxygenase-1 enzyme | Seo et al. 2018 [85] | |
| Primary cortical neurons from wild-type and Nrf2 knockout male C57BL/6J mouse | Glyceollin mixture | - Suppress glutamate-induce cytotoxicity in primary cortical neurons of wild-type mice, but not cells from Nrf2 knockout. | Seo et al. 2018 [85] | |
| Wild-type and Nrf2 knockout male C57BL/6J mouse | Glyceollin mixture | -Improve cognitive deficits caused by scopolamine in wild-type mice, but no effect in Nrf2 knockout mice - Inhibit acetylcholine esterase activity in neurons from the cortex, but not in cells from the hippocampus | Seo et al. 2018 [85] | |
| Ovariectomized adult female CFW mouse | Glyceollin mixture | - Upregulate genes involved in neurogenesis, synaptic plasticity, tissue development and transcripts involved in signal transduction pathways, regulation of nerve impulse, cytoskeletal remodeling, and hormone signaling - Downregulate genes involved in neurodegeneration apoptosis and transcripts involved in developmental neurogenesis, synaptogenesis, and cell adhesion - Act through ER 10-dependent or ER-independent mechanisms depending on the target genes | Bamji et al. 2015 [86], Bamji et al. 2018 [87] | |
| Inflammation | Murine macrophage RAW264.7 cell line | Glyceollin mixture | - Inhibit nitric oxide production and inflammatory mediators release through the NF-κB pathway | Kim et al. 2011 [88], Yoon et al. 2012 [89] |
| Human umbilical vein endothelial cell line | Glyceollin mixture | - Reduce HMGB1 11 protein level induced by LPS 12 - Decrease HMGB1-induced vascular barrier disruption - Decrease HMGB1-mediated pro-inflammatory and inflammatory response | Lee et al. 2014 [90] | |
| ICR mouse | Glyceollin mixture | - Reduce mouse ear swelling caused by 12-O-tetradecanoylphorbol-13-acetate | Kim et al. 2011 [88] | |
| Male C57BL/6 mouse that underwent cecal ligation and puncture | Glyceollin mixture | - Reduce serum HMGB1 level, decrease HMGB1-induced vascular barrier disruption - Reduce HMGB1-induced mortality | Lee et al. 2014 [90] | |
| Cardiovascular diseases | Human aortic smooth muscle cell line | Glyceollin mixture | - Inhibit the cell proliferation and migration induced by PDGF-BB 13 | Kim et al. 2012 [91] |
| Aortic rings from male Sprague-Dawley rat | GI | - Reduce vascular contraction partly through the inhibition of the RhoA/Rho-kinase signaling pathway | Song et al. 2010 [92] | |
| Melanogenesis | B16/B16F10 cell lines | Glyceollin mixture | - Inhibit melanin synthesis by decreasing melanogenic enzymes and other factors | Lee et al. 2010 [93], Shin et al. 2013 [94] |
| Standard AB strain zebrafish (Danio rerio) embryos | Glyceollin mixture | - Decrease the pigmentation of the embryos, melanin synthesis and tyrosinase activity - Decrease Sox10, a pigment cell-specific gene | Shin et al. 2013 [94] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, T.H.; Lecomte, S.; Efstathiou, T.; Ferriere, F.; Pakdel, F. An Update on the Effects of Glyceollins on Human Health: Possible Anticancer Effects and Underlying Mechanisms. Nutrients 2019, 11, 79. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010079
Pham TH, Lecomte S, Efstathiou T, Ferriere F, Pakdel F. An Update on the Effects of Glyceollins on Human Health: Possible Anticancer Effects and Underlying Mechanisms. Nutrients. 2019; 11(1):79. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010079
Chicago/Turabian StylePham, Thu Ha, Sylvain Lecomte, Theo Efstathiou, Francois Ferriere, and Farzad Pakdel. 2019. "An Update on the Effects of Glyceollins on Human Health: Possible Anticancer Effects and Underlying Mechanisms" Nutrients 11, no. 1: 79. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11010079