Blood and Brain Biochemistry and Behaviour in NTBC and Dietary Treated Tyrosinemia Type 1 Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. NTBC
2.4. Experimental Diets
2.5. Biochemical Analyses
2.6. Behavioral Analyses
2.7. Statistics
3. Results
3.1. General Health and Dietary Intake
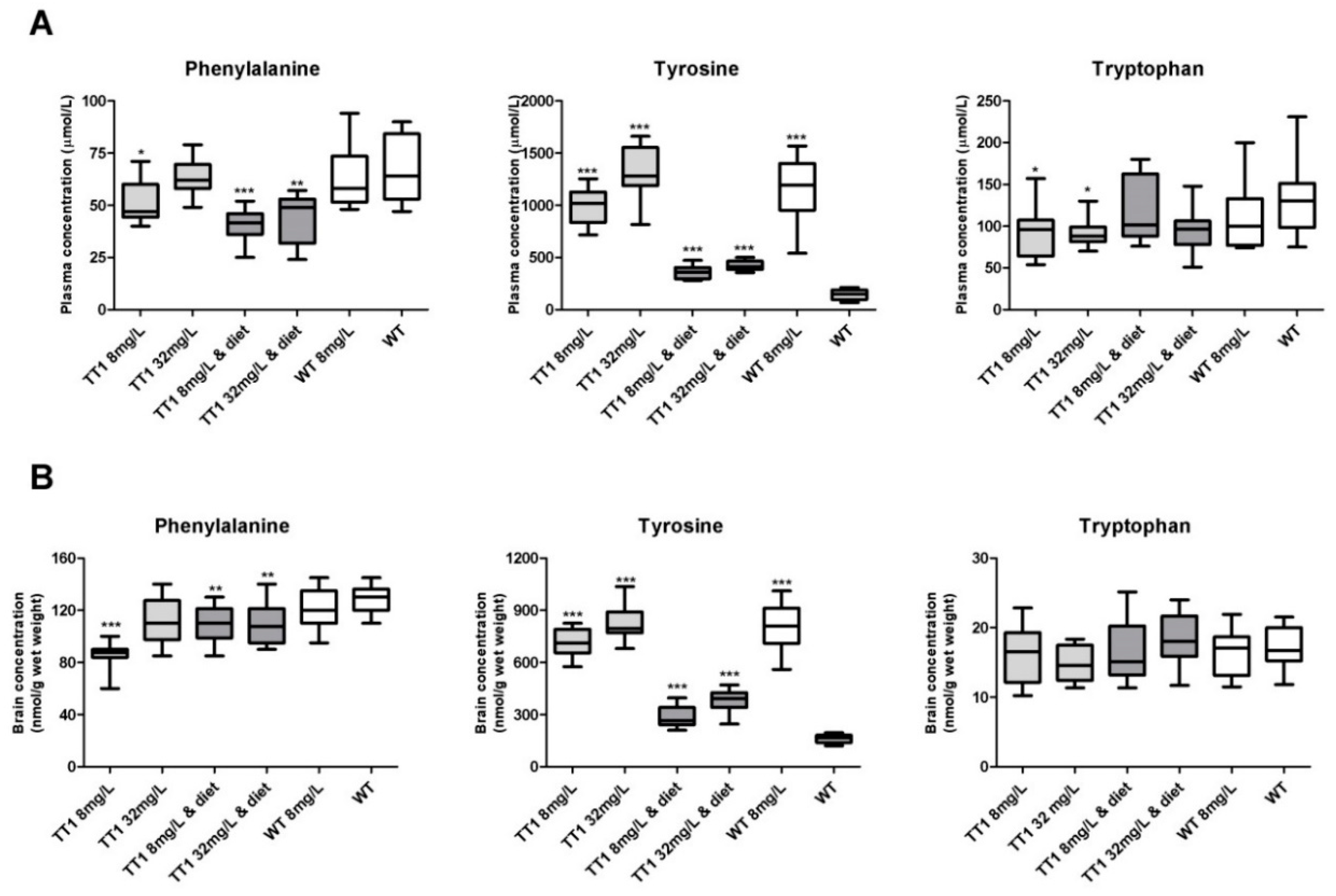
3.2. Plasma Biochemistry
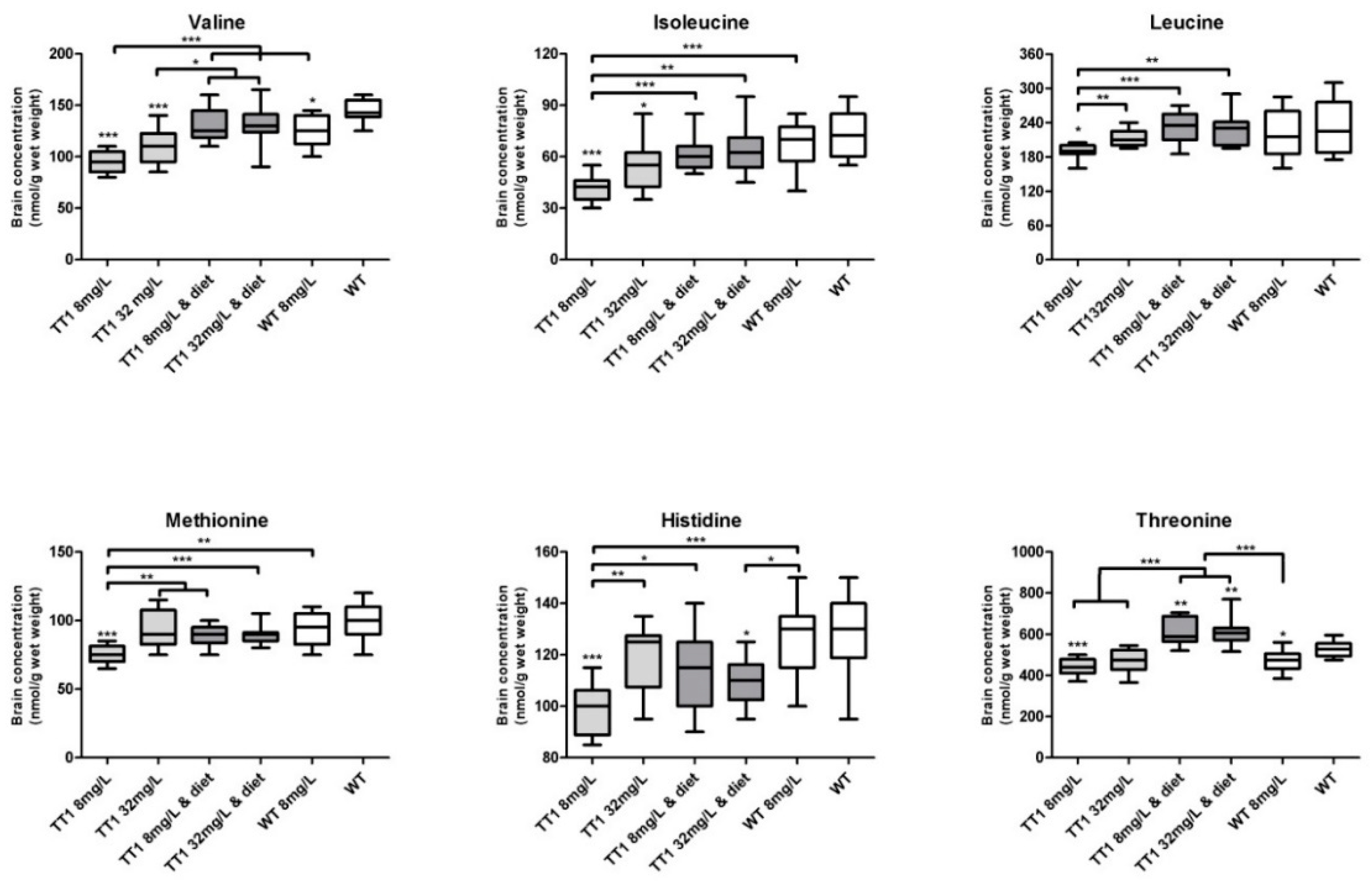
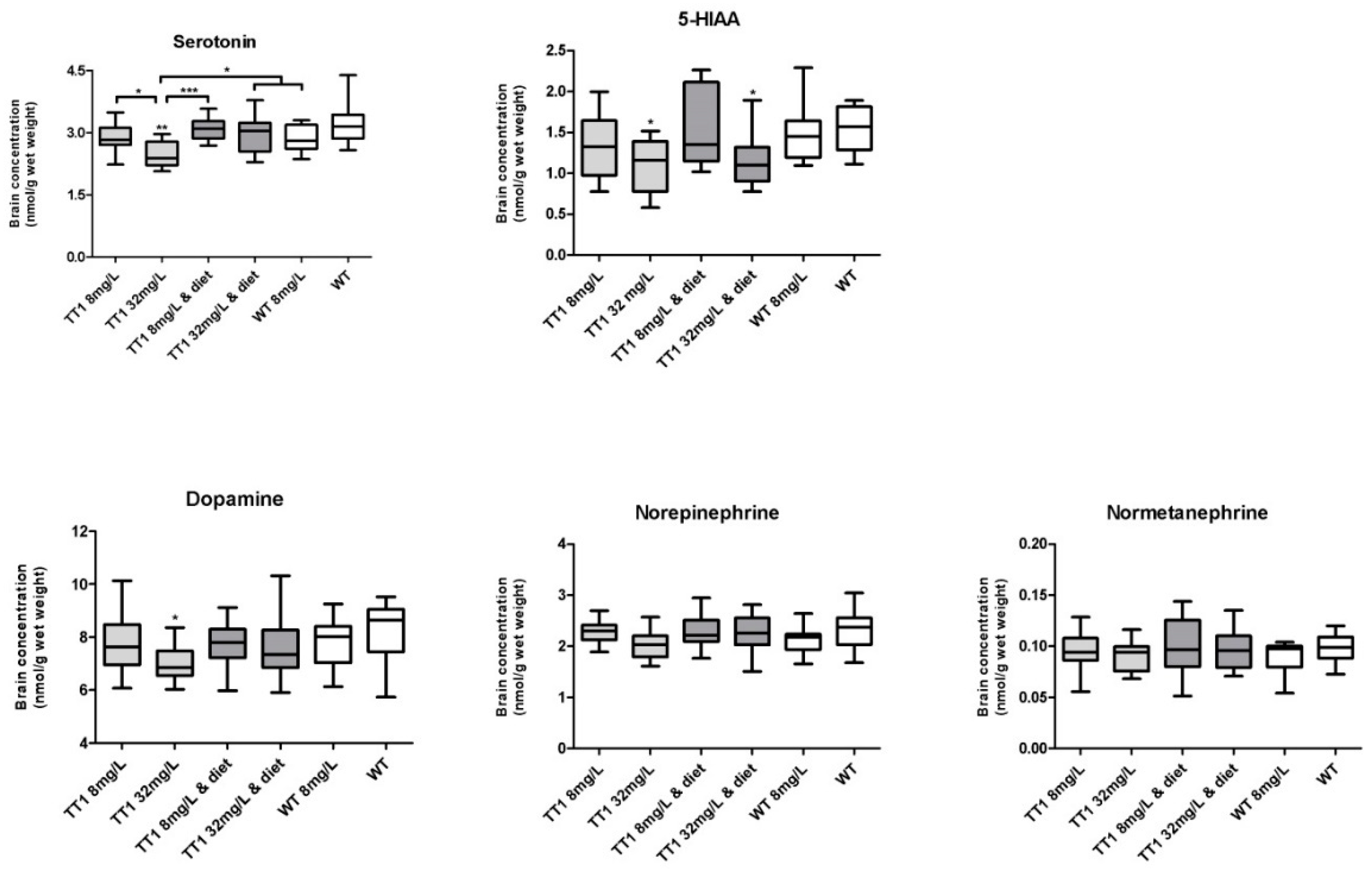
3.3. Brain Biochemistry
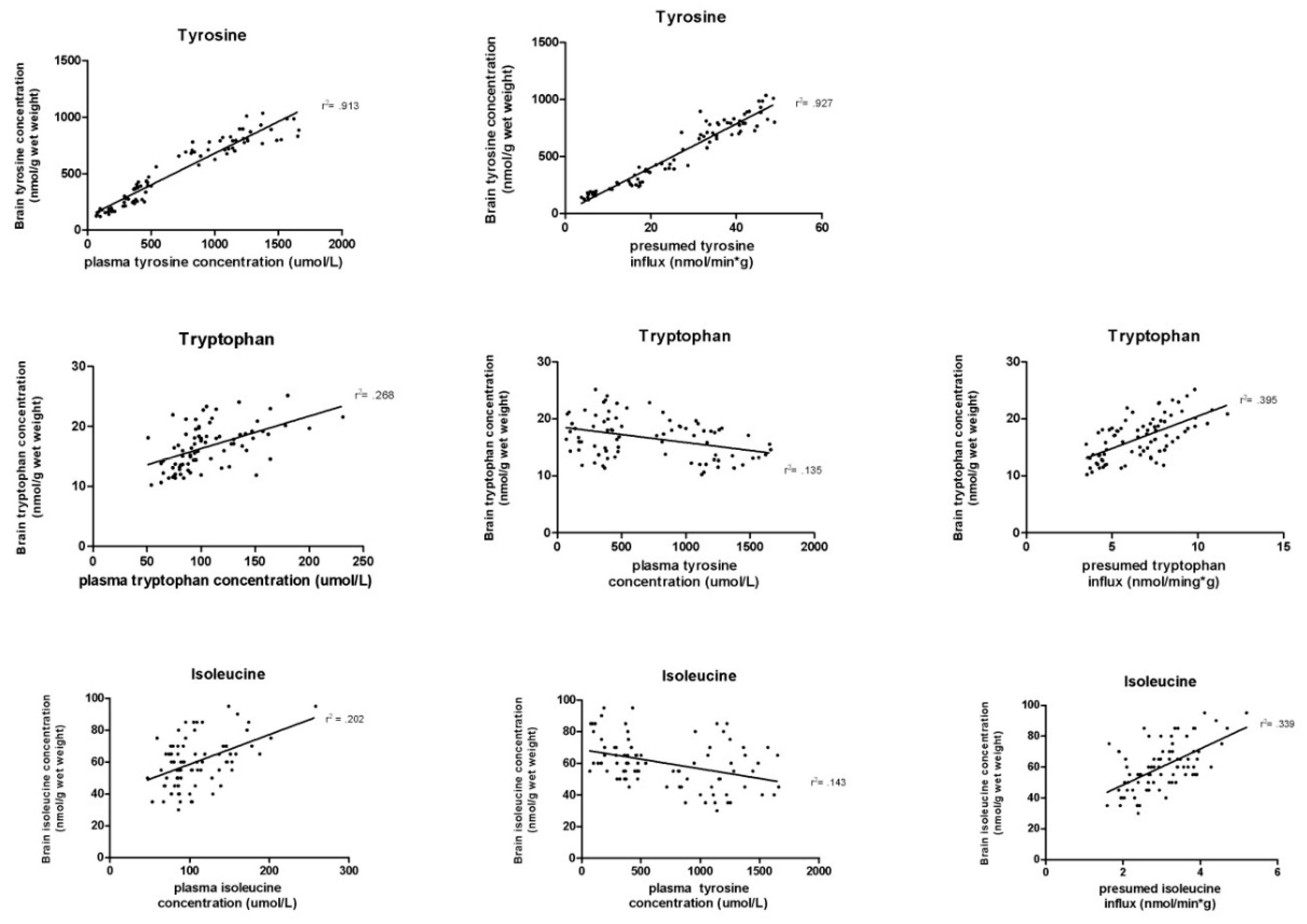
3.4. Association between Different Biochemical Parameters
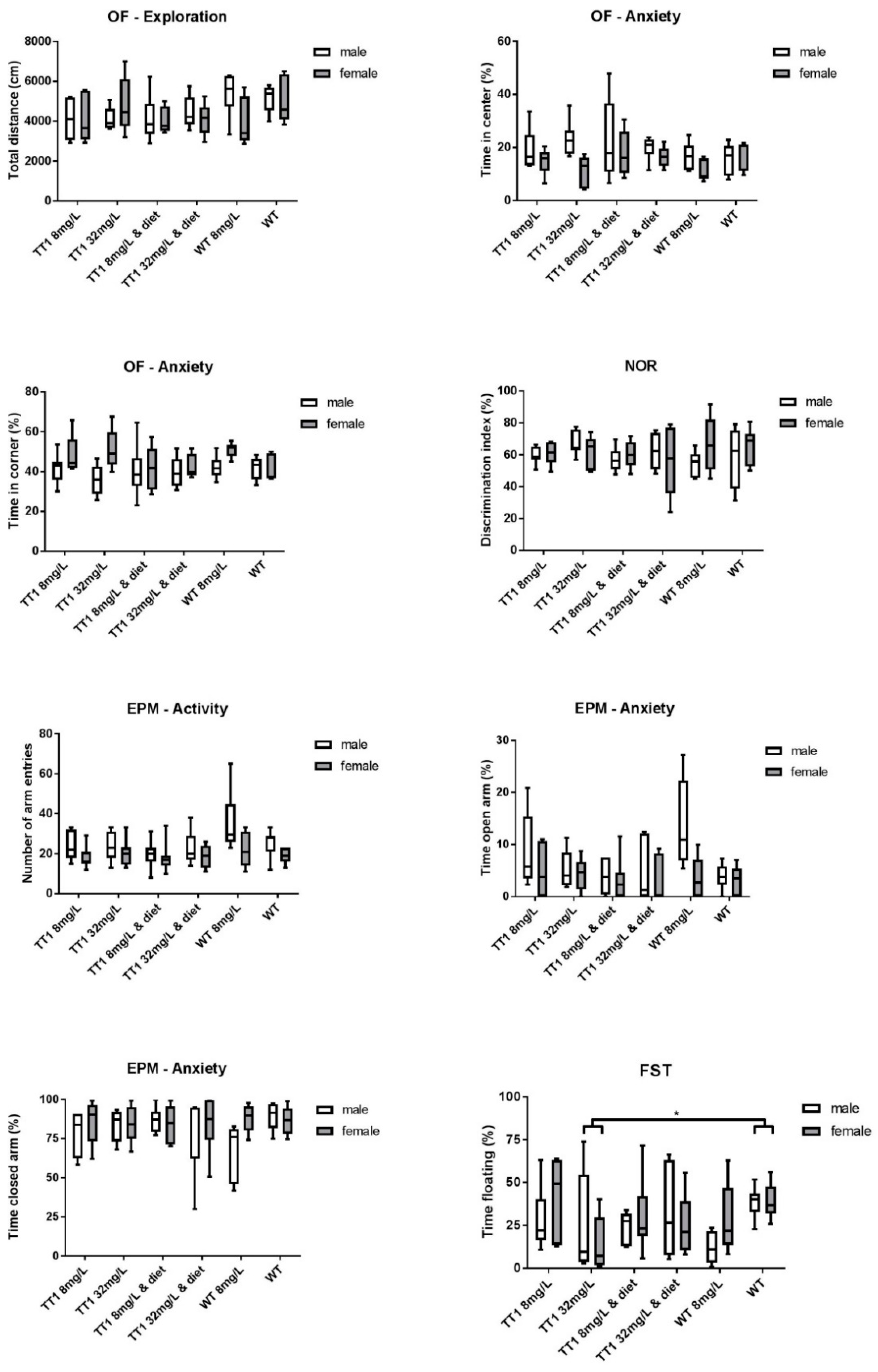
3.5. Behavioral Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NTBC | 2-(2-Nitro-4-trifluoromethylbenoyl)-1,3-cyclohexanedione |
| HPPD | 4-hydroxyphenylpyruvate dioxygenase |
| 5-HIAA | 5-hydroxyindoleacetic acid |
| EPM | elevated plus maze |
| FST | forced swim test |
| FAH | fumarylacetoacetate hydrolase |
| LNAA | large neutral amino acids |
| MSUD | maple syrup urine disease |
| NOR | novel object recognition |
| OF | open field |
| PKU | phenylketonuria |
| TT1 | tyrosinemia type 1 |
| SA | succinylacetone |
| WT | wild-type |
Appendix A

References
- De Laet, C.; Dionisi-Vici, C.; Leonard, J.V.; McKiernan, P.; Mitchell, G.; Monti, L.; de Baulny, H.O.; Pintos-Morell, G.; Spiekerkotter, U. Recommendations for the Management of Tyrosinaemia Type 1. Orphanet J. Rare Dis. 2013, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Chinsky, J.M.; Singh, R.; Ficicioglu, C.; van Karnebeek, C.D.M.; Grompe, M.; Mitchell, G.; Waisbren, S.E.; Gucsavas-Calikoglu, M.; Wasserstein, M.P.; Coakley, K.; et al. Diagnosis and Treatment of Tyrosinemia Type I: A US and Canadian Consensus Group Review and Recommendations. Genet. Med. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Van Spronsen, F.J.; van Rijn, M.; Meyer, U.; Das, A.M. Dietary Considerations in Tyrosinemia Type I. Adv. Exp. Med. Biol. 2017, 959, 197–204. [Google Scholar] [PubMed]
- Van Ginkel, W.G.; Jahja, R.; Huijbregts, S.C.J.; van Spronsen, F.J. Neurological and Neuropsychological Problems in Tyrosinemia Type I Patients. Adv. Exp. Med. Biol. 2017, 959, 111–122. [Google Scholar]
- Pardridge, W.M. Blood-Brain Barrier Carrier-Mediated Transport and Brain Metabolism of Amino Acids. Neurochem. Res. 1998, 23, 635–644. [Google Scholar] [CrossRef]
- Thimm, E.; Herebian, D.; Assmann, B.; Klee, D.; Mayatepek, E.; Spiekerkoetter, U. Increase of CSF Tyrosine and Impaired Serotonin Turnover in Tyrosinemia Type I. Mol. Genet. Metab. 2011, 102, 122–125. [Google Scholar] [CrossRef]
- Van Ginkel, W.G.; van Vliet, D.; Burgerhof, J.G.M.; de Blaauw, P.; Rubio Gozalbo, M.E.; Heiner-Fokkema, M.R.; van Spronsen, F.J. Presumptive Brain Influx of Large Neutral Amino Acids and the Effect of Phenylalanine Supplementation in Patients with Tyrosinemia Type 1. PLoS ONE 2017, 12, e0185342. [Google Scholar] [CrossRef]
- Vogel, K.R.; Arning, E.; Wasek, B.L.; McPherson, S.; Bottiglieri, T.; Gibson, K.M. Brain-Blood Amino Acid Correlates Following Protein Restriction in Murine Maple Syrup Urine Disease. Orphanet J. Rare Dis. 2014, 9, 73. [Google Scholar] [CrossRef]
- Van Vliet, D.; Bruinenberg, V.M.; Mazzola, P.N.; van Faassen, M.H.; de Blaauw, P.; Kema, I.P.; Heiner-Fokkema, M.R.; van Anholt, R.D.; van der Zee, E.A.; van Spronsen, F.J. Large Neutral Amino Acid Supplementation Exerts its Effect through Three Synergistic Mechanisms: Proof of Principle in Phenylketonuria Mice. PLoS ONE 2015, 10, e0143833. [Google Scholar] [CrossRef]
- Van Ginkel, W.G.; Jahja, R.; Huijbregts, S.C.; Daly, A.; MacDonald, A.; De Laet, C.; Cassiman, D.; Eyskens, F.; Korver-Keularts, I.M.; Goyens, P.J.; et al. Neurocognitive Outcome in Tyrosinemia Type 1 Patients Compared to Healthy Controls. Orphanet J. Rare Dis. 2016, 11, 87. [Google Scholar] [CrossRef]
- Hillgartner, M.A.; Coker, S.B.; Koenig, A.E.; Moore, M.E.; Barnby, E.; MacGregor, G.G. Tyrosinemia Type I and Not Treatment with NTBC Causes Slower Learning and Altered Behavior in Mice. J. Inherit. Metab. Dis. 2016, 39, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.E.; Koenig, A.E.; Hillgartner, M.A.; Otap, C.C.; Barnby, E.; MacGregor, G.G. Abnormal Social Behavior in Mice with Tyrosinemia Type I is Associated with an Increase of Myelin in the Cerebral Cortex. Metab. Brain Dis. 2017, 32, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Grompe, M.; al-Dhalimy, M.; Finegold, M.; Ou, C.N.; Burlingame, T.; Kennaway, N.G.; Soriano, P. Loss of Fumarylacetoacetate Hydrolase is Responsible for the Neonatal Hepatic Dysfunction Phenotype of Lethal Albino Mice. Genes Dev. 1993, 7, 2298–2307. [Google Scholar] [CrossRef] [PubMed]
- Bruinenberg, V.M.; van der Goot, E.; van Vliet, D.; de Groot, M.J.; Mazzola, P.N.; Heiner-Fokkema, M.R.; van Faassen, M.; van Spronsen, F.J.; van der Zee, E.A. The Behavioral Consequence of Phenylketonuria in Mice Depends on the Genetic Background. Front. Behav. Neurosci. 2016, 10, 233. [Google Scholar] [CrossRef]
- Kienstra, N.S.; van Reemst, H.E.; van Ginkel, W.G.; Daly, A.; van Dam, E.; MacDonald, A.; Burgerhof, J.G.M.; de Blaauw, P.; McKiernan, P.J.; Heiner-Fokkema, M.R.; et al. Daily Variation of NTBC and its Relation to Succinylacetone in Tyrosinemia Type 1 Patients Comparing a Single Dose to Two Doses a Day. J. Inherit. Metab. Dis. 2018, 41, 181–186. [Google Scholar] [CrossRef]
- Smith, Q.R. Transport of Glutamate and Other Amino Acids at the Blood-Brain Barrier. J. Nutr. 2000, 130, 1016–1022. [Google Scholar] [CrossRef]
- Strauss, K.A.; Brumbaugh, J.; Duffy, A.; Wardley, B.; Robinson, D.; Hendrickson, C.; Tortorelli, S.; Moser, A.B.; Puffenberger, E.G.; Rider, N.L.; et al. Safety, Efficacy and Physiological Actions of a Lysine-Free, Arginine-Rich Formula to Treat Glutaryl-CoA Dehydrogenase Deficiency: Focus on Cerebral Amino Acid Influx. Mol. Genet. Metab. 2011, 104, 93–106. [Google Scholar] [CrossRef]
- Harding, C.O.; Winn, S.R.; Gibson, K.M.; Arning, E.; Bottiglieri, T.; Grompe, M. Pharmacologic Inhibition of L-Tyrosine Degradation Ameliorates Cerebral Dopamine Deficiency in Murine Phenylketonuria (PKU). J. Inherit. Metab. Dis. 2014, 37, 735–743. [Google Scholar] [CrossRef]
- Van Vliet, D.; van Dam, E.; van Rijn, M.; Derks, T.G.; Venema-Liefaard, G.; Hitzert, M.M.; Lunsing, R.J.; Heiner-Fokkema, M.R.; van Spronsen, F.J. Infants with Tyrosinemia Type 1: Should Phenylalanine be Supplemented. In JIMD Reports; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Van Vliet, D.; Bruinenberg, V.M.; Mazzola, P.N.; van Faassen, M.H.; de Blaauw, P.; Pascucci, T.; Puglisi-Allegra, S.; Kema, I.P.; Heiner-Fokkema, M.R.; van der Zee, E.A.; et al. Therapeutic Brain Modulation with Targeted Large Neutral Amino Acid Supplements in the Pah-Enu2 Phenylketonuria Mouse Model. Am. J. Clin. Nutr. 2016, 104, 1292–1300. [Google Scholar] [CrossRef]
- Macedo, L.G.; Carvalho-Silva, M.; Ferreira, G.K.; Vieira, J.S.; Olegario, N.; Goncalves, R.C.; Vuolo, F.S.; Ferreira, G.C.; Schuck, P.F.; Dal-Pizzol, F.; et al. Effect of Acute Administration of L-Tyrosine on Oxidative Stress Parameters in Brain of Young Rats. Neurochem. Res. 2013, 38, 2625–2630. [Google Scholar] [CrossRef]
- De Pra, S.D.; Ferreira, G.K.; Carvalho-Silva, M.; Vieira, J.S.; Scaini, G.; Leffa, D.D.; Fagundes, G.E.; Bristot, B.N.; Borges, G.D.; Ferreira, G.C.; et al. L-Tyrosine Induces DNA Damage in Brain and Blood of Rats. Neurochem. Res. 2014, 39, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Teodorak, B.P.; Scaini, G.; Carvalho-Silva, M.; Gomes, L.M.; Teixeira, L.J.; Rebelo, J.; De Pra, S.D.; Zeni, N.; Schuck, P.F.; Ferreira, G.C.; et al. Antioxidants Reverse the Changes in Energy Metabolism of Rat Brain After Chronic Administration of L.-Tyrosine. Metab. Brain Dis. 2017, 32, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Brain Metabolism: A Perspective from the Blood-Brain Barrier. Physiol. Rev. 1983, 63, 1481–1535. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.; Williams, D.L. Enhancement of Rat Brain Catecholamine Synthesis by Administration of Small Doses of Tyrosine and Evidence for Substrate Inhibition of Tyrosine Hydroxylase Activity by Large Doses of the Amino Acid. Biochem. J. 1982, 206, 165–168. [Google Scholar] [CrossRef]
- Ribeiro, P.; Pigeon, D.; Kaufman, S. The Hydroxylation of Phenylalanine and Tyrosine by Tyrosine Hydroxylase from Cultured Pheochromocytoma Cells. J. Biol. Chem. 1991, 266, 16207–16211. [Google Scholar]
- Reed, M.C.; Lieb, A.; Nijhout, H.F. The Biological Significance of Substrate Inhibition: A Mechanism with Diverse Functions. Bioessays 2010, 32, 422–429. [Google Scholar] [CrossRef]
- Tekin, I.; Roskoski, R.; Carkaci-Salli, N.; Vrana, K.E. Complex Molecular Regulation of Tyrosine Hydroxylase. J. Neural Transm. 2014, 121, 1451–1481. [Google Scholar] [CrossRef]
- Naoi, M.; Maruyama, W.; Takahashi, T.; Ota, M.; Parvez, H. Inhibition of Tryptophan Hydroxylase by Dopamine and the Precursor Amino Acids. Biochem. Pharmacol. 1994, 48, 207–211. [Google Scholar]
- Al-Dhalimy, M.; Overturf, K.; Finegold, M.; Grompe, M. Long-Term Therapy with NTBC and Tyrosine-Restricted Diet in a Murine Model of Hereditary Tyrosinemia Type I. Mol. Genet. Metab. 2002, 75, 38–45. [Google Scholar] [CrossRef]
- Masurel-Paulet, A.; Poggi-Bach, J.; Rolland, M.O.; Bernard, O.; Guffon, N.; Dobbelaere, D.; Sarles, J.; de Baulny, H.O.; Touati, G. NTBC Treatment in Tyrosinaemia Type I: Long-Term Outcome in French Patients. J. Inherit. Metab. Dis. 2008, 31, 81–87. [Google Scholar] [CrossRef]
- De Laet, C.; Munoz, V.T.; Jaeken, J.; Francois, B.; Carton, D.; Sokal, E.M.; Dan, B.; Goyens, P.J. Neuropsychological Outcome of NTBC-Treated Patients with Tyrosinaemia Type 1. Dev. Med. Child Neurol. 2011, 53, 962–964. [Google Scholar] [CrossRef] [PubMed]
- Thimm, E.; Richter-Werkle, R.; Kamp, G.; Molke, B.; Herebian, D.; Klee, D.; Mayatepek, E.; Spiekerkoetter, U. Neurocognitive Outcome in Patients with Hypertyrosinemia Type I After Long-Term Treatment with NTBC. J. Inherit. Metab. Dis. 2012, 35, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Bendadi, F.; de Koning, T.J.; Visser, G.; Prinsen, H.C.; de Sain, M.G.; Verhoeven-Duif, N.; Sinnema, G.; van Spronsen, F.J.; van Hasselt, P.M. Impaired Cognitive Functioning in Patients with Tyrosinemia Type I Receiving Nitisinone. J. Pediatr. 2014, 164, 398–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.I.; de la Parra, A.; Arias, C.; Arredondo, M.; Cabello, J.F. Long-Term Cognitive Functioning in Individuals with Tyrosinemia Type 1 Treated with Nitisinone and Protein-Restricted Diet. Mol. Genet. Metab. Rep. 2017, 11, 12–16. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Ginkel, W.G.; van Vliet, D.; van der Goot, E.; Faassen, M.H.J.R.; Vogel, A.; Heiner-Fokkema, M.R.; van der Zee, E.A.; van Spronsen, F.J. Blood and Brain Biochemistry and Behaviour in NTBC and Dietary Treated Tyrosinemia Type 1 Mice. Nutrients 2019, 11, 2486. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102486
van Ginkel WG, van Vliet D, van der Goot E, Faassen MHJR, Vogel A, Heiner-Fokkema MR, van der Zee EA, van Spronsen FJ. Blood and Brain Biochemistry and Behaviour in NTBC and Dietary Treated Tyrosinemia Type 1 Mice. Nutrients. 2019; 11(10):2486. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102486
Chicago/Turabian Stylevan Ginkel, Willem G., Danique van Vliet, Els van der Goot, Martijn H. J. R. Faassen, Arndt Vogel, M. Rebecca Heiner-Fokkema, Eddy. A. van der Zee, and Francjan J. van Spronsen. 2019. "Blood and Brain Biochemistry and Behaviour in NTBC and Dietary Treated Tyrosinemia Type 1 Mice" Nutrients 11, no. 10: 2486. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102486