Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease

 , , ,
, , ,
Abstract
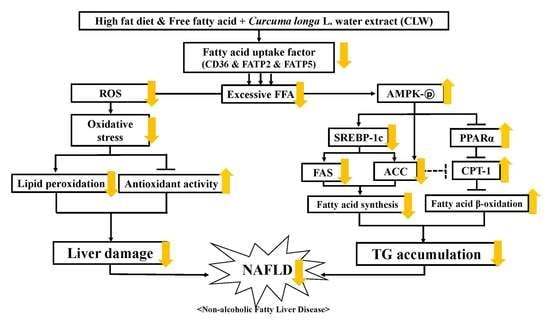
:
1. Introduction
2. Materials and Methods
2.1. Sample and Chemicals
2.2. Cell Culture
2.3. FFA-Induced Lipid Overloading in HepG2 Cells
2.4. Measurement of Intracellular ROS Formation
2.5. Measurement of Lipid Accumulation (Oil Red O Staining)
2.6. Animal Experiments
2.7. Assays for Serum Marker Enzymes, and Hepatic Triglyceride, and Total Cholesterol
2.8. Hematoxylin and Eosin Staining
2.9. Measurement of Antioxidant Enzyme Activity
2.10. Total RNA Isolation and Real-Time Polymerase Chain Reaction
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
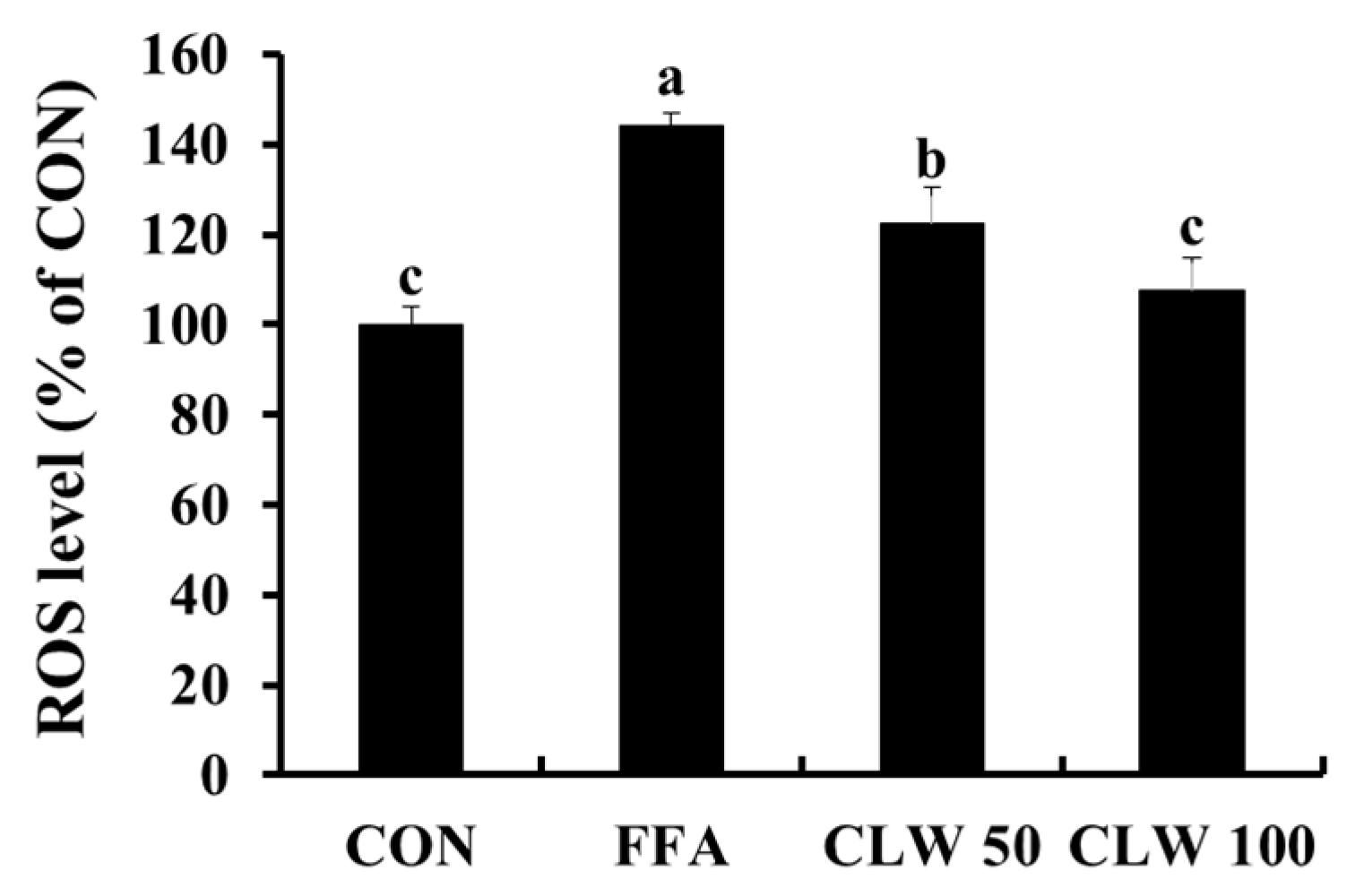
3.1. Effect of CLW on Antioxidant Activity in FFA-Treated Cells
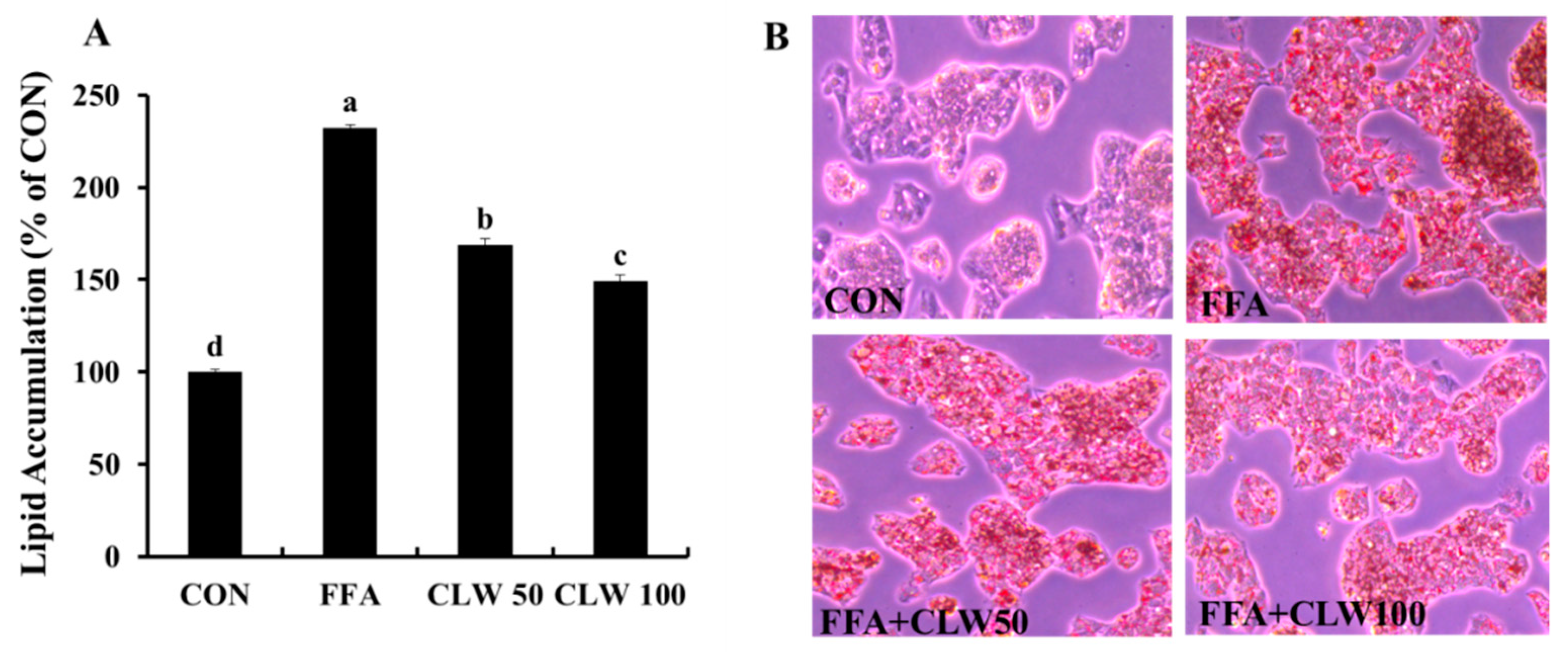
3.2. Effect of CLW on Lipid Accumulation in FFA-Treated Cells
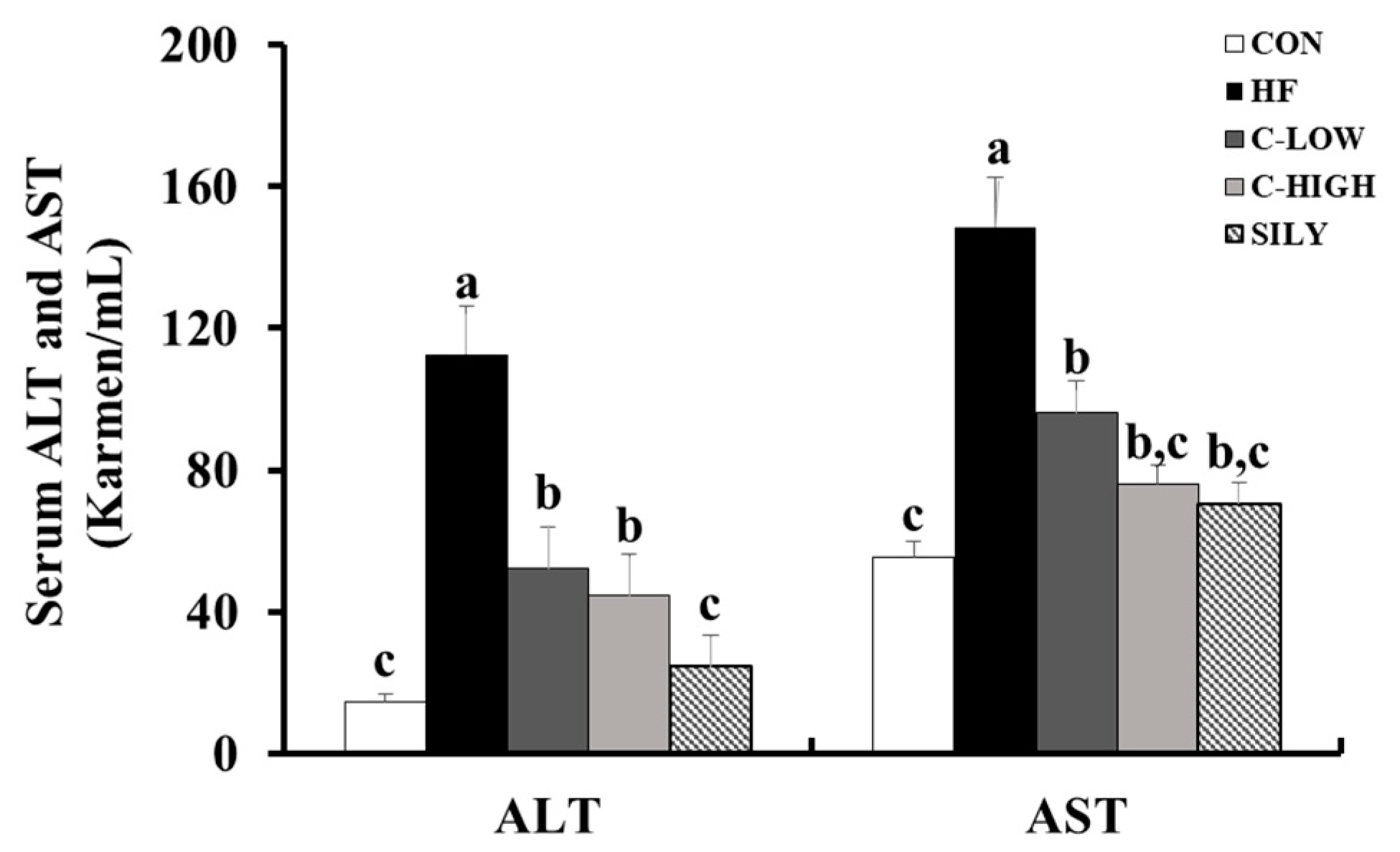
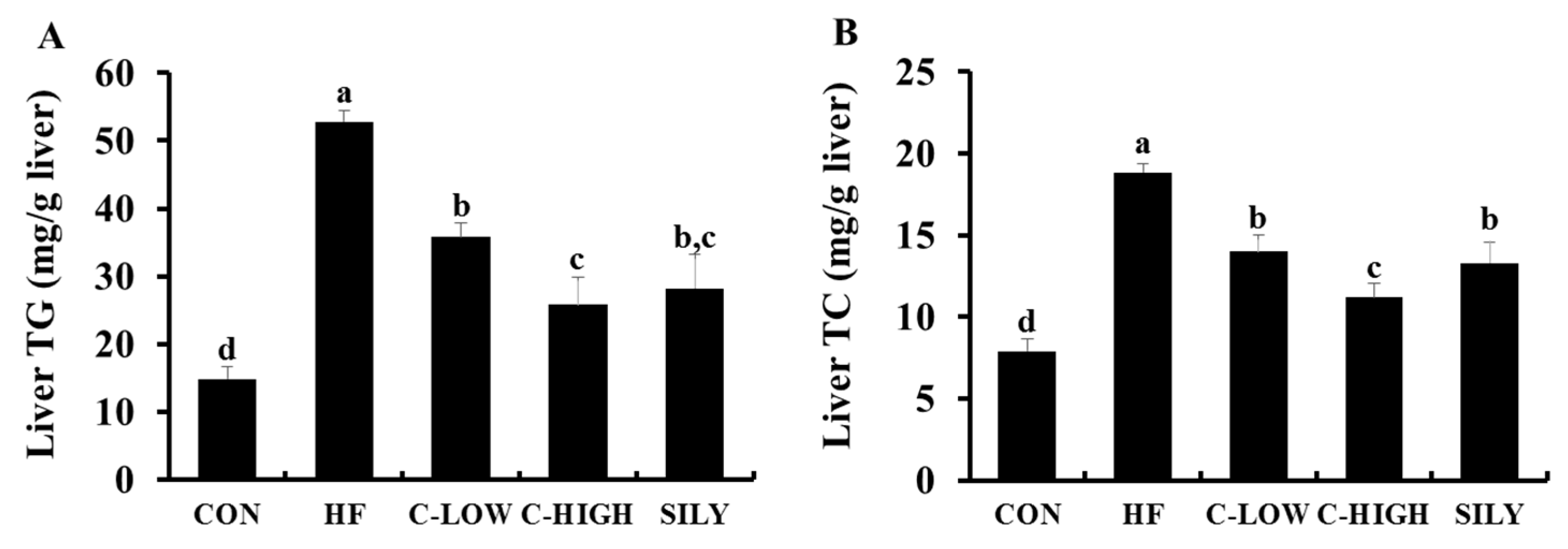
3.3. Effect of CLW on HF-Induced Hepatotoxicity in Mice
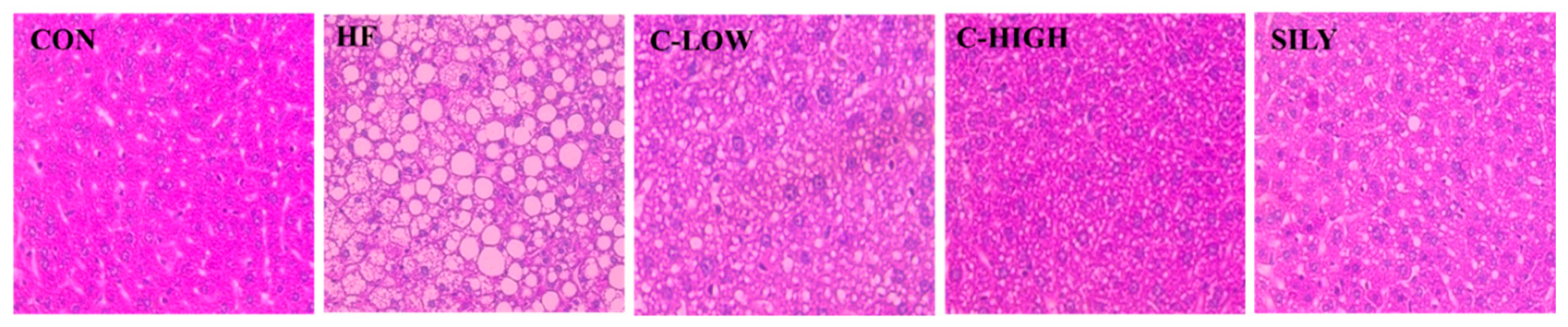
3.4. Effect of CLW on Histopathological Morphology
3.5. Changes in Antioxidant Activities in Fatty Liver by CLW Treatment
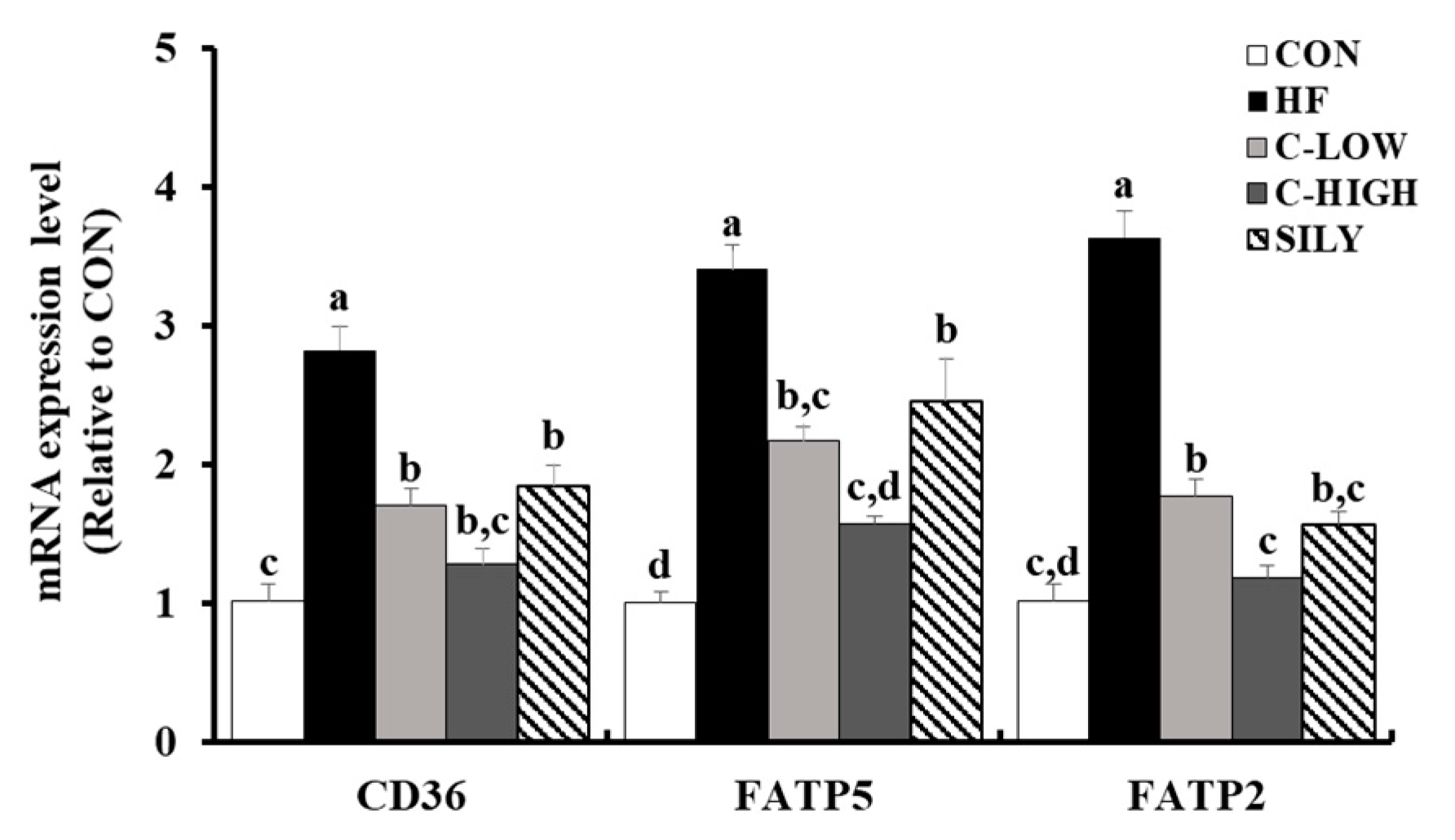
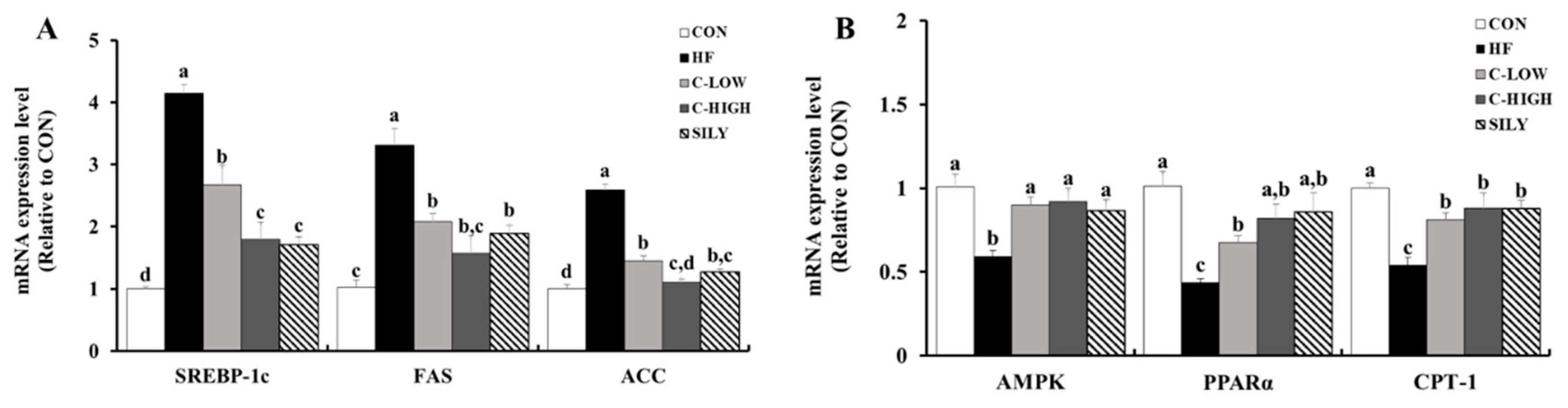
3.6. Effect of CLW on Fatty Acid Uptake-Related mRNA Expression in HF Diet-fed Mice
3.7. Effect of CLW on Lipid Accumulation-Related mRNA Expression in HF Diet-fed Mice
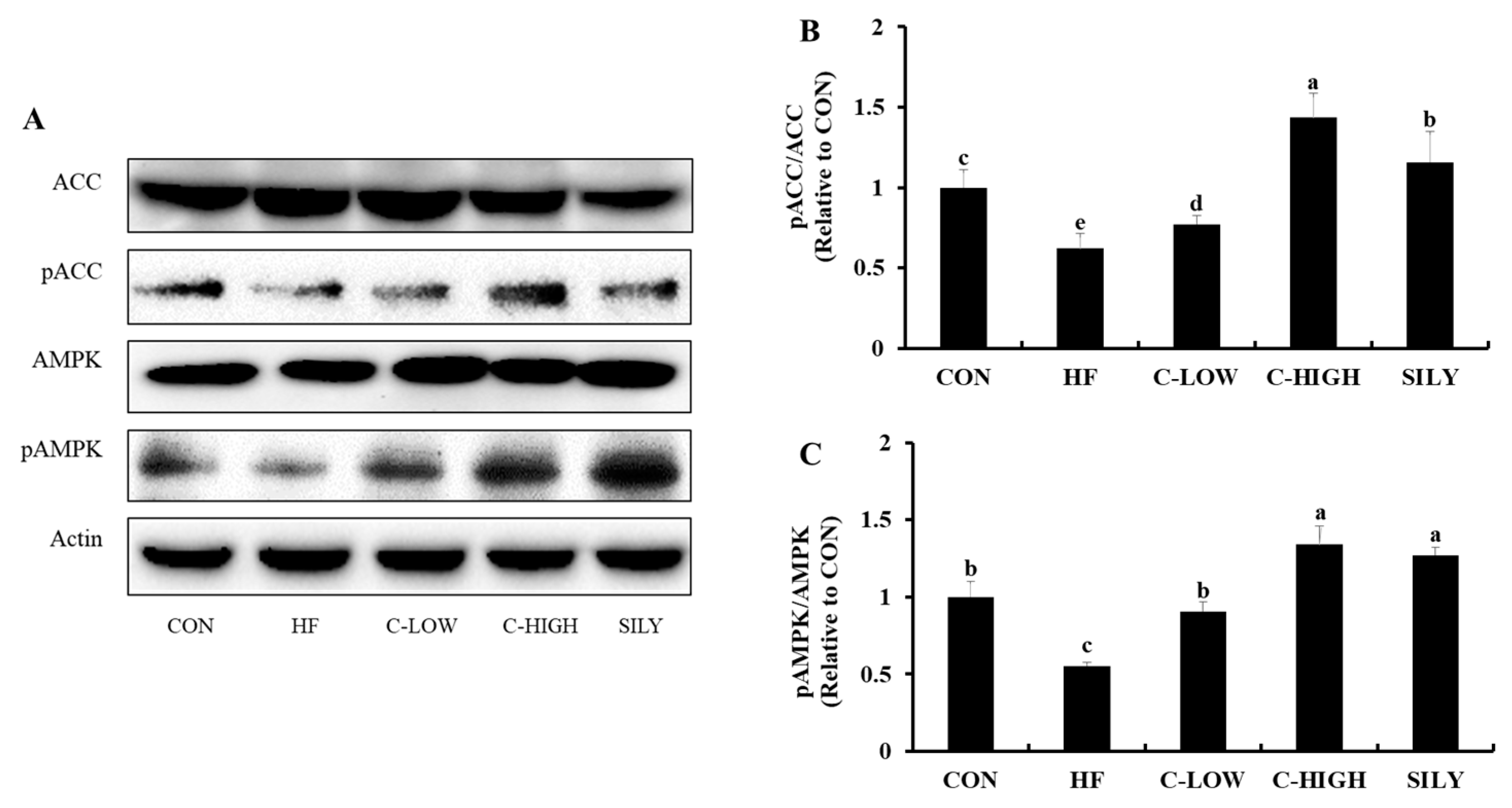
3.8. Effect of CLW on Activation of AMPK and ACC in HF Diet-fed Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Purohit, V.; Russo, D.; Coates, P.M. Role of fatty liver, dietary fatty acid supplements, and obesity in the progression of alcoholic liver disease: Introduction and summary of the symposium. Alcohol 2004, 34, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Yeh, M.M.; Brunt, E.M. Pathological features of fatty liver disease. Gastroenterology 2014, 147, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M. Pathology of fatty liver disease. Mod. Pathol. 2007, 20, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- Yasui, K.; Hashimoto, E.; Komorizono, Y.; Koike, K.; Arii, S.; Imai, Y.; Shima, T.; Kanbara, Y.; Saibara, T.; Mori, T.; et al. Characteristics of patients with nonalcoholic steatohepatitis who develop hepatocellular carcinoma. Clin. Gastroenterol. Hepatol. 2011, 9, 428–433. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Muralidharan, P.; Raj, J.P. Update in global trends and aetiology of hepatocellular carcinoma. Contemp. Oncol. 2018, 22, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Koonen, D.P.Y.; Jacobs, R.L.; Febbraio, M.; Young, M.E.; Soltys, C.L.M.; Ong, H.; Vance, D.E.; Dyck, J.R.B. Increased hepatic CD36 expression contributes to dyslipidemia associated with diet-induced obesity. Diabetes 2007, 56, 2863–2871. [Google Scholar] [CrossRef]
- Ouwens, D.M.; Diamant, M.; Fodor, M.; Habets, D.D.J.; Pelsers, M.M.A.L.; El Hasnaoui, M.; Dang, Z.C.; van den Brom, C.E.; Vlasblom, R.; Rietdijk, A.; et al. Cardiac contractile dysfunction in insulin-resistant rats fed a high-fat diet is associated with elevated CD36-mediated fatty acid uptake and esterification. Diabetologia 2007, 50, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Vallve, J.C.; Uliaque, K.; Girona, J.; Cabre, A.; Ribalta, J.; Heras, M.; Masana, L. Unsaturated fatty acids and their oxidation products stimulate CD36 gene expression in human macrophages. Atherosclerosis 2002, 164, 45–56. [Google Scholar] [CrossRef]
- Zhong, S.; Zhao, L.; Wang, Y.; Zhang, C.; Liu, J.; Wang, P.; Zhou, W.; Yang, P.; Varghese, Z.; Moorhead, J.F.; et al. Cluster of differentiation 36 deficiency aggravates macrophage infiltration and hepatic inflammation by upregulating monocyte chemotactic protein-1 expression of hepatocytes through histone deacetylase 2-dependent pathway. Antioxid. Redox Signal. 2017, 27, 201–214. [Google Scholar] [CrossRef]
- You, Y.; Min, S.; Lee, Y.-H.; Hwang, K.; Jun, W. Hepatoprotective effect of 10% ethanolic extract from Curdrania tricuspidata leaves against ethanol-induced oxidative stress through suppression of CYP2E1. Food Chem. Toxicol. 2017, 108, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Shin, H.-S.; Choi, H.S.; Park, J.-W.; Jo, I.; Oh, E.-S.; Lee, K.-Y.; Lee, B.-H.; Johnson, R.J.; Kang, D.-H. Uric acid induces fat accumulation via generation of endoplasmic reticulum stress and SREBP-1c activation in hepatocytes. Lab. Investig. 2014, 94, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, R.S.; Surya, D.; Nalini, N. Antioxidant efficacy of black pepper (Piper nigrum L.) and piperine in rats with high fat diet induced oxidative stress. Redox Rep. 2004, 9, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Song, F.L.; Gan, R.Y.; Zhang, Y.; Xiao, Q.; Kuang, L.; Li, H.-B. Total phenolic contents and antioxidant capacities of selected chinese medicinal plants. Int. J. Mol. Sci. 2010, 11, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, Y.; Yoon, H.-G.; You, Y.; Park, J.; Lee, Y.-H.; Kim, S.; Hwang, K.; Lee, J.; Jun, W. Prevention of ethanol-induced hepatotoxicity by fermented Curcuma longa L. in C57BL/6 Mice. Food Sci. Biotechnol. 2014, 23, 925–930. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm J. 2013, 21, 143–152. [Google Scholar] [CrossRef]
- Garg, S.; Garg, A.; Shukla, A.; Dev, S.K.; Bishnoi, R.S.; Kumar, M. Review on antioxidant evaluation methods: And models In-vitro in-vivo. Asian J. Pharm. Pharmacol. 2018, 4, 147–154. [Google Scholar] [CrossRef]
- Molina, M.F.; Sanchez-Reus, I.; Iglesias, I.; Benedi, J. Quercetin, a flavonoid antioxidant, prevents and protects against ethanol-induced oxidative stress in mouse liver. Biol. Pharm. Bull. 2003, 26, 1398–1402. [Google Scholar] [CrossRef]
- Al Batran, R.; Al-Bayaty, F.; Al-Obaidi, M.M.J.; Abdualkader, A.M.; Hadi, H.A.; Ali, H.M.; Abdulla, M.A. In vivo antioxidant and antiulcer activity of Parkia speciosa ethanolic leaf extract against ethanol-induced gastric ulcer in rats. PLoS ONE 2013, 8, e64751. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [PubMed]
- Akerboom, T.P.; Sies, H. Assay of glutathione, glutathione disulfide, and glutathione mixed disulfides in biological samples. Methods Enzymol. 1981, 77, 373–382. [Google Scholar]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Garcia, M.C.; Amankwa-Sakyi, M.; Flynn, T.J. Cellular glutathione in fatty liver in vitro models. Toxicol. Vitr. 2011, 25, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Xu, S.; Choi, S.-K.; Ha, C.-M.; Thoudam, T.; Cha, S.-K.; Wiederkehr, A.; Wollheim, C.B.; Lee, I.-K.; Park, K.-S. Oxidative stress and calcium dysregulation by palmitate in type 2 diabetes. Exp. Mol. Med. 2017, 49, 291. [Google Scholar]
- Gomez-Lechon, M.J.; Donato, M.T.; Martinez-Romero, A.; Jimenez, N.; Castell, J.V.; O’Connor, J.E. A human hepatocellular in vitro model to investigate steatosis. Chem. Biol. Interact. 2007, 165, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metabolism 2008, 57, 1071–1077. [Google Scholar] [CrossRef]
- Clugston, R.D.; Yuen, J.J.; Hu, Y.; Abumrad, N.A.; Berk, P.D.; Goldberg, I.J.; Blaner, W.S.; Huang, L.-S. CD36-deficient mice are resistant to alcohol- and high-carbohydrate-induced hepatic steatosis. J. Lipid Res. 2013, 55, 239–246. [Google Scholar] [CrossRef]
- Tanikawa, K.; Torimura, T. Studies on oxidative stress in liver diseases: Important future trends in liver research. Med. Mol. Morphol. 2006, 39, 22–27. [Google Scholar] [CrossRef]
- Kim, Y.; You, Y.; Yoon, H.G.; Lee, Y.H.; Kim, K.; Lee, J.; Kim, M.S.; Kim, J.C.; Jun, W. Hepatoprotective effects of fermented Curcuma longa L. on carbon tetrachloride-induced oxidative stress in rats. Food Chem. 2014, 15, 148–153. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Hassan, W.; Rongyin, G.; Daoud, A.; Ding, L.; Wang, L.; Liu, J.; Shang, J. Reduced Oxidative Stress Contributes to the Lipid Lowering Effects of Isoquercitrin in Free Fatty Acids Induced Hepatocytes. Oxid. Med. Cell. Longev. 2014, 2014, 313602. [Google Scholar] [CrossRef] [PubMed]
- Song, C.Y.; Shi, J.; Zeng, X.; Zhang, Y.; Xie, W.F.; Chen, Y.X. Sophocarpine alleviates hepatocyte steatosis through activating AMPK signaling pathway. Toxicol. In Vitro 2013, 27, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Park, K.I.; Ma, J.Y. Leonurus japonicus houtt attenuates nonalcoholic fatty liver disease in free fatty acid-induced HepG2 cells and mice fed a high-fat diet. Nutrients 2018, 10, 20. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, J.-J.; Cho, W.-K.; Yim, N.-H.; Kim, H.-K.; Yun, B.; Ma, J.Y. KBH-1, an herbal composition, improves hepatic steatosis and leptin resistance in high-fat diet-induced obese rats. BMC Complement. Altern. Med. 2016, 16, 1–12. [Google Scholar]
- Kim, M.K.; Kim, S.H.; Yu, H.S.; Park, H.G.; Kang, U.G.; Ahn, Y.M.; Kim, Y.S. The effect of clozapine on the AMPK-ACC-CPT1 pathway in the rat frontal cortex. Int. J. Neuropsychopharmacol. 2012, 15, 907–917. [Google Scholar] [CrossRef]
- Tailleux, A.; Wouters, K.; Staels, B. Roles of PPARs in NAFLD: Potential therapeutic targets. BBA Mol. Cell Biol. Lipids 2012, 1821, 809–818. [Google Scholar] [CrossRef]
- Cacciapuoti, F.; Scognamiglio, A.; Palumbo, R.; Forte, R.; Cacciapuoti, F. Silymarin in non alcoholic fatty liver disease. World J. Hepatol. 2013, 5, 109–113. [Google Scholar] [CrossRef]
- Kim, M.H.; Seong, J.B.; Huh, J.W.; Bae, Y.C.; Lee, H.S.; Lee, D.S. Peroxiredoxin 5 ameliorates obesity-induced non-alcoholic fatty liver disease through the regulation of oxidative stress and AMP-activated protein kinase signaling. Redox. Biol. 2020, 3, 101315. [Google Scholar] [CrossRef]
- Saitta, C.; Pollicino, T.; Raimondo, G. Obesity and liver cancer. Ann. Hepatol. 2019, in press. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Sequence (5′ to 3′) |
|---|---|---|
| CYP2E1 | Forward Reverse | 5′- CGTGGAAATGGAGAAGGAAA-3′ 5′- GGTGATGAACCGCTGAATCT-3′ |
| AMPK | Forward Reverse | 5′- GGCACCCTCCCATTTGATG-3′ 5′- ACACCCCCTCGGATCTTCTT-3′ |
| ACC | Forward Reverse | 5′- TGCAGATCTTAGCGGACCAA-3′ 5′- GCCTGCGTTGTACAGAGCAA-3′ |
| CPT-1 | Forward Reverse | 5′- TGTTGGGTATGCTGTTCATGACA-3′ 5′- GCGGCCTGGGTAGGAAGA-3′ |
| SREBP-1c | Forward Reverse | 5′- CGGAACCATCTTGGCAACA-3′ 5′- GCCGGTTGATAGGCAGCTT-3′ |
| PPAR-α | Forward Reverse | 5′- AACATCCAAGAGATTTCGCAATC-3′ 5′- CCGTAAAGCCAAAGCTTCCA-3′ |
| CD36 | Forward Reverse | 5′- TGGAACAGAGGCTGACAACT-3′ 5′- TTGATTTTTGATAGATATGGG-3′ |
| FATP2 | Forward Reverse | 5′- CTTTCAGCACATTGCTGATTACCT-3′ 5′-CAGTGATCTCAATGGTGTCCTGTAT-3′ |
| FATP5 | Forward Reverse | 5′- AGCTCCTGCGGTACTTGTGT-3′ 5′- AAGGTCTCCCACACATCAGC-3′ |
| β-Actin | Forward Reverse | 5′- ACGGCCAGGTCATCACTATTG-3′ 5′- CAAGAAGGAAGGCTGGAAAAGA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mun, J.; Kim, S.; Yoon, H.-G.; You, Y.; Kim, O.-K.; Choi, K.-C.; Lee, Y.-H.; Lee, J.; Park, J.; Jun, W. Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease. Nutrients 2019, 11, 2536. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102536
Mun J, Kim S, Yoon H-G, You Y, Kim O-K, Choi K-C, Lee Y-H, Lee J, Park J, Jun W. Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease. Nutrients. 2019; 11(10):2536. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102536
Chicago/Turabian StyleMun, Jeongeun, Shintae Kim, Ho-Geun Yoon, Yanghee You, Ok-Kyung Kim, Kyung-Chul Choi, Yoo-Hyun Lee, Jeongmin Lee, Jeongjin Park, and Woojin Jun. 2019. "Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease" Nutrients 11, no. 10: 2536. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102536