Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies


Abstract
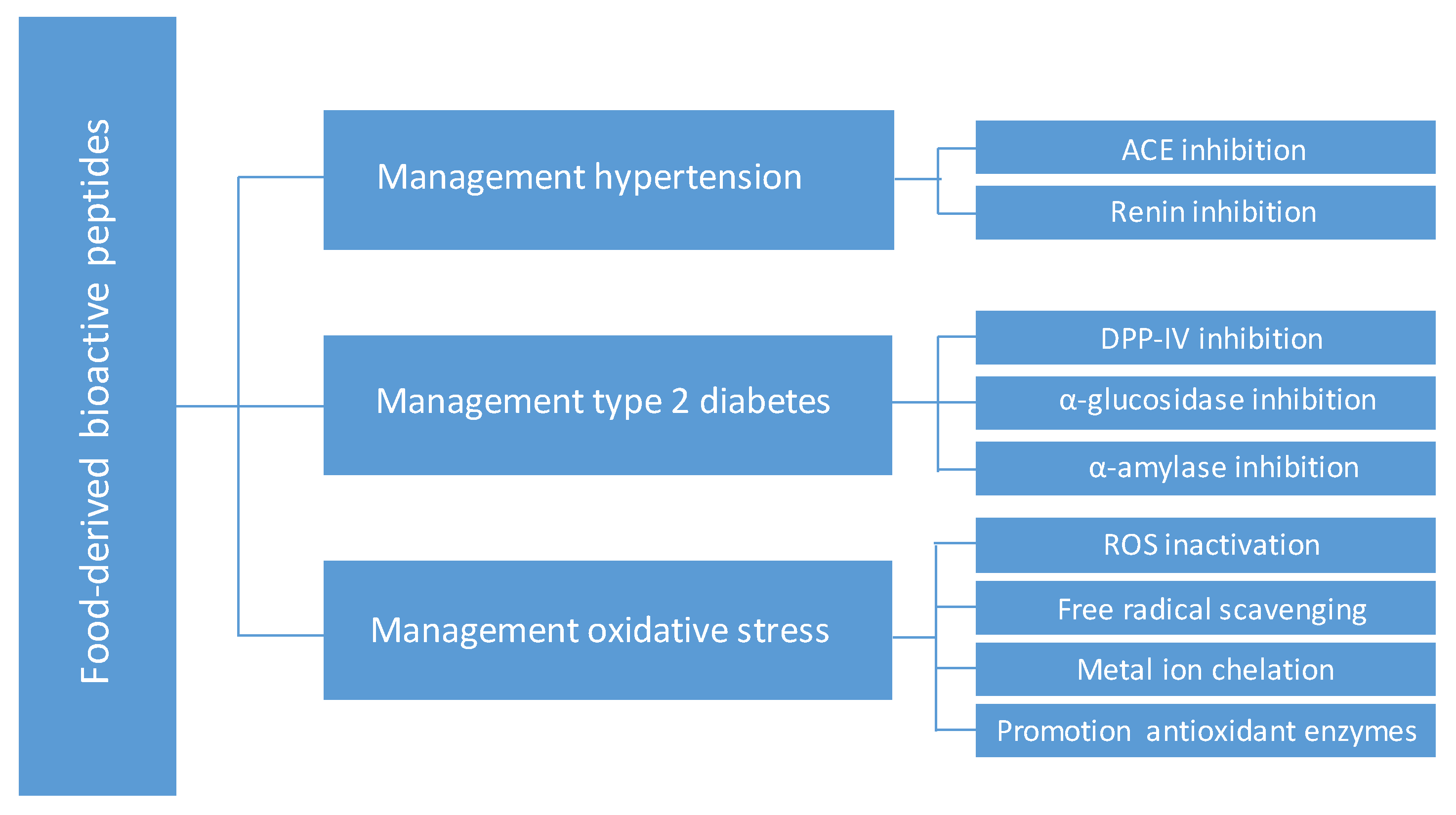
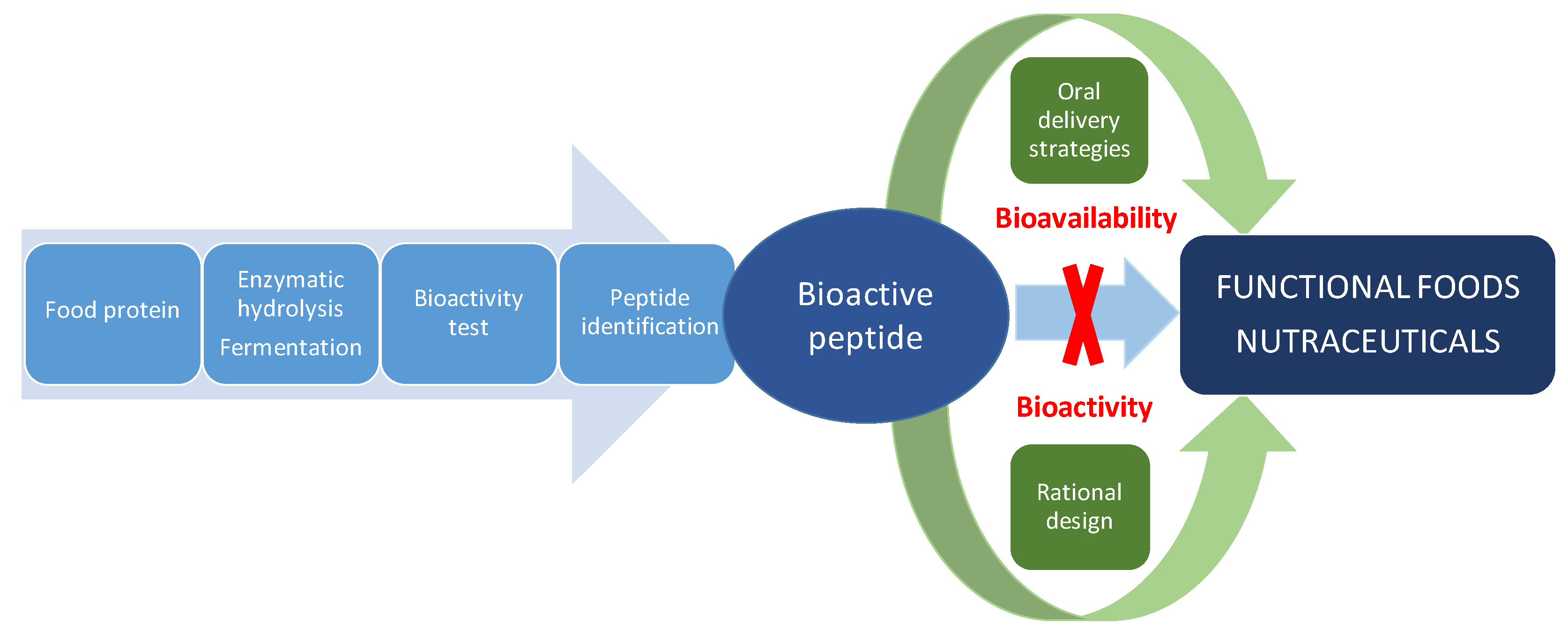
:1. Introduction
2. Structural Requirements of Bioactive Peptides
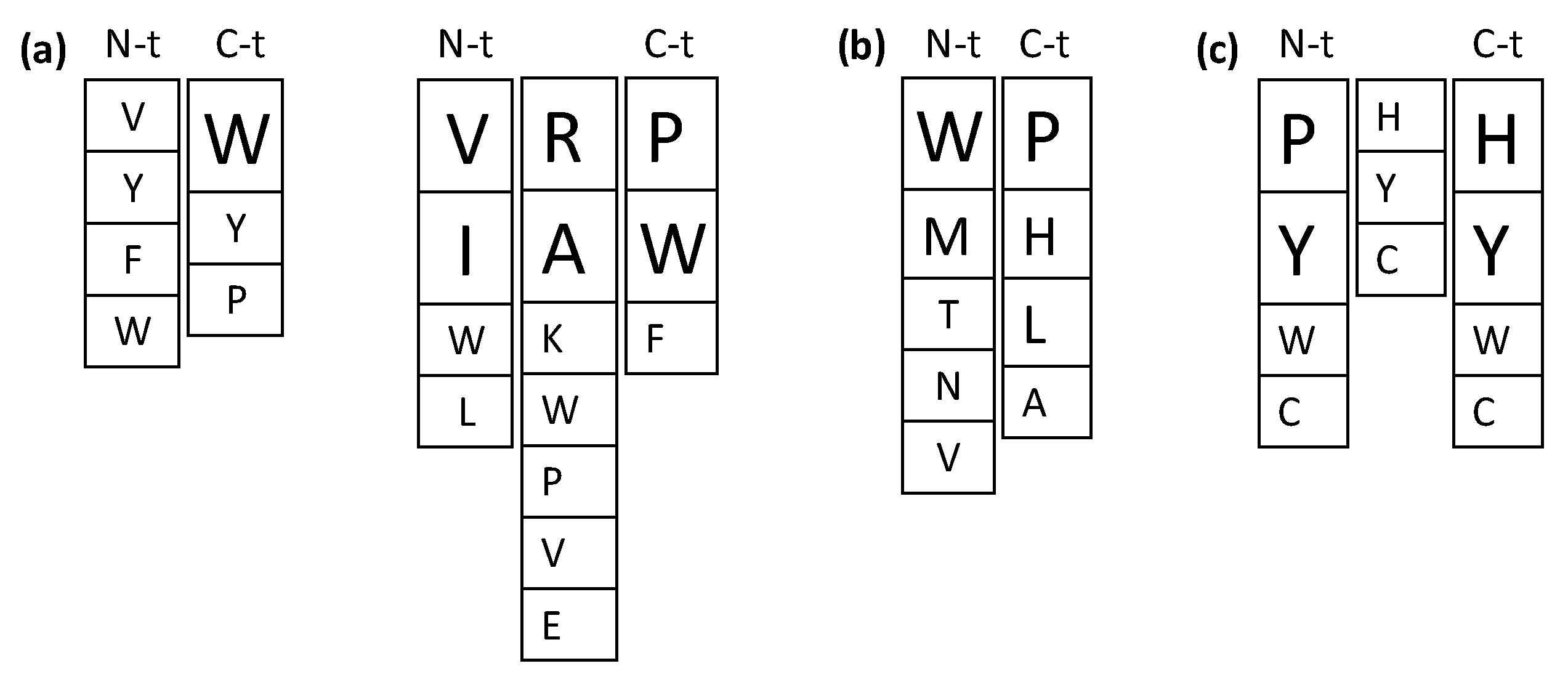
2.1. Antihypertensive Peptides
2.2. Antidiabetic Peptides
2.3. Antioxidant Peptides
3. Oral Delivery Strategies
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haller, C.A. Nutraceuticals: Has there been any progress? Clin. Pharmacol. Ther. 2010, 87, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, S.K.; Maiti, T.K. Targeting tumors with peptides from natural sources. Trends Biotechnol. 2008, 26, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.; Schroder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef]
- Manzanares, P.; Salom, J.B.; Garcia-Tejedor, A.; Fernandez-Musoles, R.; Ruiz-Gimenez, P.; Gimeno-Alcaniz, J.V. Unraveling the mechanisms of action of lactoferrin-derived antihypertensive peptides: ACE inhibition and beyond. Food Funct. 2015, 6, 2440–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, R.M.; Siragy, H.M. Newly recognized components of the renin-angiotensin system: Potential roles in cardiovascular and renal regulation. Endocr. Rev. 2003, 24, 261–271. [Google Scholar] [CrossRef]
- Fragasso, G.; Maranta, F.; Montanaro, C.; Salerno, A.; Torlasco, C.; Margonato, A. Pathophysiologic therapeutic targets in hypertension: A cardiological point of view. Expert Opin. Ther. Targets 2012, 16, 179–193. [Google Scholar] [CrossRef]
- Martinez-Maqueda, D.; Miralles, B.; Recio, I.; Hernandez-Ledesma, B. Antihypertensive peptides from food proteins: A review. Food Funct. 2012, 3, 350–361. [Google Scholar] [CrossRef]
- Murray, B.A.; FitzGerald, R.J. Angiotensin converting enzyme inhibitory peptides derived from food proteins: Biochemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods 2014, 8, 45–52. [Google Scholar] [CrossRef]
- Deacon, C.F. Dipeptidyl peptidase-IV inhibitors in the treatment of type 2 diabetes: A comparative review. Diabetes Obes. Metab. 2011, 13, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Kajaria, D.; Ranjana; Tripathi, J.; Tripathi, Y.; Tiwari, S. In-vitro α-amylase and glycosidase inhibitory effect of ethanolic extract of antiasthmatic drug—Shirishadi. J. Adv. Pharm. Technol. Res. 2013, 4, 206–209. [Google Scholar] [PubMed]
- Liu, R.; Cheng, J.; Wu, H. Discovery of food-derived dipeptidyl peptidase IV inhibitory peptides: A review. Int. J. Mol. Sci. 2019, 20, 463. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Prospects for the management of type 2 diabetes using food protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Curr. Opin. Food Sci. 2016, 8, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Bamdad, F.; Ahmed, S.; Chen, L. Specifically designed peptide structures effectively suppressed oxidative reactions in chemical and cellular systems. J. Funct. Foods 2015, 18, 35–46. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Marcos, J.F.; Muñoz, A.; Pérez-Payá, E.; Misra, S.; López-García, B. Identification and rational design of novel antimicrobial peptides for plant protection. Annu. Rev. Phytopathol. 2008, 46, 273–301. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; Ryan, B.; Duffy, A.; Brayden, D.J. Formulation strategies to improve oral peptide delivery. Pharm. Pat. Anal. 2014, 3, 313–336. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, P.; Pechenov, S.; Anand Subramony, J. Oral peptide delivery: Translational challenges due to physiological effects. J. Control. Release 2018, 287, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Cheng, S.; Lu, W.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. TrAC Trends Analyt. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- Majumder, K.; Wu, J. Molecular targets of antihypertensive peptides: Understanding the mechanisms of action based on the pathophysiology of hypertension. Int. J. Mol. Sci. 2014, 16, 256–283. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.S.; Cushman, D.W. Inhibition of homogeneous angiotensin I-converting enzyme of rabbit lung by synthetic venom peptides of Bothrops jararaca. Biochim. Biophys. Acta 1973, 293, 451–463. [Google Scholar] [CrossRef]
- Cushman, D.W.; Pluščec, J.; Williams, N.J.; Weaver, E.R.; Sabo, E.F.; Kocy, O.; Cheung, H.S.; Ondetti, M.A. Inhibition of angiotensin-converting enzyme by analogs of peptides from Bothrops jararaca venom. Experientia 1973, 29, 1032–1035. [Google Scholar] [CrossRef]
- Cushman, D.W.; Cheung, H.S.; Sabo, E.F.; Ondetti, M.A. Design of potent competitive inhibitors of angiotensin-converting enzyme: Carboxyalkanoyl and mercaptoalkanoyl amino acids. Biochemistry 1977, 16, 5484–5491. [Google Scholar] [CrossRef]
- Cheung, H.S.; Wang, F.L.; Ondetti, M.A.; Sabo, E.F.; Cushman, D.W. Binding of peptide-substrates and inhibitors of angiotensin-converting enzyme—Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar]
- Masuda, O.; Nakamura, Y.; Takano, T. Antihypertensive peptides are present in aorta after oral administration of sour milk containing these peptides to spontaneously hypertensive rats. J. Nutr. 1996, 126, 3063–3068. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and characterization of angiotensin I-converting enzyme-inhibitors from sour milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T. Antihypertensive effect of sour milk and peptides isolated from it that are inhibitors to angiotensin I-converting enzyme. J. Dairy Sci. 1995, 78, 1253–1257. [Google Scholar] [CrossRef]
- Turpeinen, A.M.; Järvenpää, S.; Kautiainen, H.; Korpela, R.; Vapaatalo, H. Antihypertensive effects of bioactive tripeptides—A random effects meta-analysis. Ann. Med. 2013, 45, 51–56. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure-sctivity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Deng, B.; Ni, X.; Zhai, Z.; Tang, T.; Tan, C.; Yan, Y.; Deng, J.; Yin, Y. New quantitative structure–activity relationship model for angiotensin-converting enzyme inhibitory dipeptides based on integrated descriptors. J. Agric. Food Chem. 2017, 65, 9774–9781. [Google Scholar] [CrossRef]
- Norris, R.; Casey, F.; FitzGerald, R.J.; Shields, D.; Mooney, C. Predictive modelling of angiotensin converting enzyme inhibitory dipeptides. Food Chem. 2012, 133, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Vukic, V.R.; Vukic, D.V.; Milanovic, S.D.; Ilicic, M.D.; Kanuric, K.G.; Johnson, M.S. In silico identification of milk antihypertensive di-and tripeptides involved in angiotensin I–converting enzyme inhibitory activity. Nutr. Res. 2017, 46, 22–30. [Google Scholar] [CrossRef]
- Panyayai, T.; Sangsawad, P.; Pacharawongsakda, E.; Sawatdichaikul, O.; Tongsima, S.; Choowongkomon, K. The potential peptides against angiotensin-I converting enzyme through a virtual tripeptide-constructing library. Comput. Biol. Chem. 2018, 77, 207–213. [Google Scholar] [CrossRef]
- Xie, C.L.; Choung, S.Y.; Cao, G.P.; Lee, K.W.; Choi, Y.J. In silico investigation of action mechanism of four novel angiotensin-I converting enzyme inhibitory peptides modified with Trp. J. Funct. Foods 2015, 17, 632–639. [Google Scholar] [CrossRef]
- Jing, P.; Qian, B.; He, Y.; Zhao, X.; Zhang, J.; Zhao, D.; Lv, Y.; Deng, Y. Screening milk-derived antihypertensive peptides using quantitative structure activity relationship (QSAR) modelling and in vitro/in vivo studies on their bioactivity. Int. Dairy J. 2014, 35, 95–101. [Google Scholar] [CrossRef]
- Ding, F.Y.; Qian, B.J.; Zhao, X.; Shen, S.Q.; Deng, Y.; Wang, D.F.; Zhang, F.; Sui, Z.Q.; Jing, P. VPPIPP and IPPVPP: Two hexapeptides innovated to exert antihypertensive activity. PLoS ONE 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Lin, G.; Zhang, R.; Wu, W. Studies on the bioactivities of ACE-inhibitory peptides with phenylalanine C-terminus using 3D-QSAR, molecular docking and in vitro evaluation. Mol. Inf. 2017, 36, 1600157. [Google Scholar] [CrossRef] [PubMed]
- Centeno, J.M.; Burguete, M.C.; Castello-Ruiz, M.; Enrique, M.; Valles, S.; Salom, J.B.; Torregrosa, G.; Marcos, J.F.; Alborch, E.; Manzanares, P. Lactoferricin-related peptides with inhibitory effects on ACE-dependent vasoconstriction. J. Agric. Food Chem. 2006, 54, 5323–5329. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Giménez, P.; Marcos, J.F.; Torregrosa, G.; Lahoz, A.; Fernández-Musoles, R.; Vallés, S.; Alborch, E.; Manzanares, P.; Salom, J.B. Novel antihypertensive hexa-and heptapeptides with ACE-inhibiting properties: From the in vitro ACE assay to the spontaneously hypertensive rat. Peptides 2011, 32, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Quiros, A.; Contreras, M.D.; Ramos, M.; Amigo, L.; Recio, I. Stability to gastrointestinal enzymes and structure-activity relationship of β-casein-peptides with antihypertensive properties. Peptides 2009, 30, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Li, H.; Aluko, R.E. Quantitative structure-activity relationship modeling of renin-inhibiting dipeptides. Amino Acids 2012, 42, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Cheng, C.; Li, Y. Structure-based design and optimization of antihypertensive peptides to obtain high inhibitory potency against both renin and angiotensin I-converting enzyme. SAR QSAR Environ. Res. 2015, 26, 1001–1016. [Google Scholar] [CrossRef]
- Xie, C.-L.; Kim, J.-S.; Ha, J.-M.; Choung, S.-Y.; Choi, Y.-J. Angiotensin I-converting enzyme inhibitor derived from cross-linked oyster protein. BioMed Res. Int. 2014, 2014, 9. [Google Scholar] [CrossRef]
- Fujita, H.; Yokoyama, K.; Yoshikawa, M. Classification and anthihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar]
- Nii, Y.; Fukuta, K.; Yoshimoto, R.; Sakai, K.; Ogawa, T. Determination of antihypertensive peptides from an izumi shrimp hydrolysate. Biosci. Biotechnol. Biochem. 2008, 72, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A. Angiotensin I-converting enzyme inhibitory peptides derived from wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.; Martin, M.; Lunow, D.; Rudolph, S.; Mertten, S.; Möckel, U.; Deußen, A.; Henle, T. Tryptophan-containing dipeptides are bioavailable and inhibit plasma human angiotensin-converting enzyme in vivo. Int. Dairy J. 2016, 52, 107–114. [Google Scholar] [CrossRef]
- Khedr, S.; Deussen, A.; Kopaliani, I.; Zatschler, B.; Martin, M. Effects of tryptophan-containing peptides on angiotensin-converting enzyme activity and vessel tone ex vivo and in vivo. Eur. J. Nutr. 2018, 57, 907–915. [Google Scholar] [CrossRef]
- Tian, F.; Zhou, P.; Lv, F.; Song, R.; Li, Z. Three-dimensional holograph vector of atomic interaction field (3D-HoVAIF): A novel rotation–translation invariant 3D structure descriptor and its applications to peptides. J. Peptide Sci. 2007, 13, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Liu, Y.-L.; Ning, J.-H.; Yu, J.; Li, X.-H.; Wang, F.-X. Is the structural diversity of tripeptides sufficient for developing functional food additives with satisfactory multiple bioactivities? J. Mol. Struct. 2013, 1040, 164–170. [Google Scholar] [CrossRef]
- Takano, T. Milk derived peptides and hypertension reduction. Int. Dairy J. 1998, 8, 375–381. [Google Scholar] [CrossRef]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sorhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure-activity relationship modeling of peptides containing 4–10 amino acid residues. QSAR Comb. Sci. 2006, 25, 873–880. [Google Scholar] [CrossRef]
- Sagardia, I.; Roa-Ureta, R.H.; Bald, C. A new QSAR model, for angiotensin I-converting enzyme inhibitory oligopeptides. Food Chem. 2013, 136, 1370–1376. [Google Scholar] [CrossRef]
- Garcia-Tejedor, A.; Sanchez-Rivera, L.; Castello-Ruiz, M.; Recio, I.; Salom, J.B.; Manzanares, P. Novel antihypertensive lactoferrin-derived peptides produced by Kluyveromyces marxianus: Gastrointestinal stability profile and in vivo angiotensin I-converting enzyme (ACE) inhibition. J. Agric. Food Chem. 2014, 62, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Savitha, M.N.; Siddesha, J.M.; Suvilesha, K.N.; Yariswamy, M.; Vivek, H.K.; D’Souza, C.J.M.; Umashankar, M.; Vishwanath, B.S. Active-site directed peptide L-Phe-D-His-L-Leu inhibits angiotensin converting enzyme activity and dexamethasone-induced hypertension in rats. Peptides 2019, 112, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Giménez, P.; Ibañez, A.; Salom, J.B.; Marcos, J.F.; López-Díez, J.J.; Vallés, S.; Torregrosa, G.; Alborch, E.; Manzanares, P. Antihypertensive properties of lactoferricin B-derived peptides. J. Agric. Food Chem. 2010, 58, 6721–6727. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, R.J.; Meisel, H. Milk protein-derived peptide inhibitors of angiotensin-I-converting enzyme. Br. J. Nutr. 2000, 84, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagebayashi, T.; Kontani, N.; Yamada, Y.; Mizushige, T.; Arai, T.; Kino, K.; Ohinata, K. Novel CCK-dependent vasorelaxing dipeptide, Arg-Phe, decreases blood pressure and food intake in rodents. Mol. Nutr. Food Res. 2012, 56, 1456–1463. [Google Scholar] [CrossRef] [PubMed]
- Kontani, N.; Omae, R.; Kagebayashi, T.; Kaneko, K.; Yamada, Y.; Mizushige, T.; Kanamoto, R.; Ohinata, K. Characterization of Ile-His-Arg-Phe, a novel rice-derived vasorelaxing peptide with hypotensive and anorexigenic activities. Mol. Nutr. Food Res. 2014, 58, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Sasai, M.; Sun, X.; Okuda, C.; Nakato, J.; Kanamoto, R.; Ohinata, K. Orally active anti-hypertensive peptides found based on enteroendocrine cell responses to a dipeptide library. Biochem. Biophys. Res. Commun. 2018, 503, 1070–1074. [Google Scholar] [CrossRef]
- Foltz, M.; van Buren, L.; Klaffke, W.; Duchateau, G.S. Modeling of the relationship between dipeptide structure and dipeptide stability, permeability, and ACE inhibitory activity. J. Food Sci. 2009, 74, H243–H251. [Google Scholar] [CrossRef]
- Mentlein, R. Dipeptidyl-peptidase IV (CD26)-role in the inactivation of regulatory peptides. Regul. Pept. 1999, 85, 9–24. [Google Scholar] [CrossRef]
- Hikida, A.; Ito, K.; Motoyama, T.; Kato, R.; Kawarasaki, Y. Systematic analysis of a dipeptide library for inhibitor development using human dipeptidyl peptidase IV produced by a Saccharomyces cerevisiae expression system. Biochem. Biophys. Res. Commun. 2013, 430, 1217–1222. [Google Scholar] [CrossRef]
- Lan, V.T.T.; Ito, K.; Ito, S.; Kawarasaki, Y. Trp-Arg-Xaa tripeptides act as uncompetitive-type inhibitors of human dipeptidyl peptidase IV. Peptides 2014, 54, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Lan, V.T.T.; Ito, K.; Ohno, M.; Motoyama, T.; Ito, S.; Kawarasaki, Y. Analyzing a dipeptide library to identify human dipeptidyl peptidase IV inhibitor. Food Chem. 2015, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterisation of the potential of β-lactoglobulin and α-lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Structure activity relationship modelling of milk protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Peptides 2016, 79, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R.M. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef]
- Mojica, L.; de Mejia, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Marcuse, R. Antioxidative effect of amino-acids. Nature 1960, 186, 886–887. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-activity relationship of the antioxidant peptides from natural proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxidant activity of designed peptides based on the antioxidative peptide isolated from digests of a soybean protein. J. Agric. Food Chem. 1996, 44, 2619–2623. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef]
- Saito, K.; Jin, D.H.; Ogawa, T.; Muramoto, K.; Hatakeyama, E.; Yasuhara, T.; Nokihara, K. Antioxidative properties of tripeptide libraries prepared by the combinatorial chemistry. J. Agric. Food Chem. 2003, 51, 3668–3674. [Google Scholar] [CrossRef]
- Chen, N.; Chen, J.; Yao, B.; Li, Z.G. QSAR study on antioxidant tripeptides and the antioxidant activity of the designed tripeptides in free radical systems. Molecules 2018, 23, 12. [Google Scholar] [CrossRef]
- Gu, L.; Zhao, M.; Li, W.; You, L.; Wang, J.; Wang, H.; Ren, J. Chemical and cellular antioxidant activity of two novel peptides designed based on glutathione structure. Food Chem. Toxicol. 2012, 50, 4085–4091. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, Y.; Dong, H.; Su, G.; Zhao, M. Structure–activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. J. Funct. Foods 2016, 21, 485–496. [Google Scholar] [CrossRef]
- Matsui, R.; Honda, R.; Kanome, M.; Hagiwara, A.; Matsuda, Y.; Togitani, T.; Ikemoto, N.; Terashima, M. Designing antioxidant peptides based on the antioxidant properties of the amino acid side-chains. Food Chem. 2018, 245, 750–755. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Sun, Q.; Song, G.; Huang, J. Extraction, identification and structure-activity relationship of antioxidant peptides from sesame (Sesamum indicum L.) protein hydrolysate. Food Res. Int. 2019, 116, 707–716. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef]
- Renukuntla, J.; Vadlapudi, A.D.; Patel, A.; Boddu, S.H.S.; Mitra, A.K. Approaches for enhancing oral bioavailability of peptides and proteins. Int. J. Pharm. 2013, 447, 75–93. [Google Scholar] [CrossRef] [Green Version]
- McClements, D.J. Enhancing nutraceutical bioavailability through food matrix design. Curr. Opin. Food Sci. 2015, 4, 1–6. [Google Scholar] [CrossRef]
- Ting, Y.; Jiang, Y.; Ho, C.-T.; Huang, Q. Common delivery systems for enhancing in vivo bioavailability and biological efficacy of nutraceuticals. J. Funct. Foods 2014, 7, 112–128. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Ryan, S.M.; Brayden, D.J. Oral delivery strategies for nutraceuticals: Delivery vehicles and absorption enhancers. Trends Food Sci. Technol. 2016, 53, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Gleeson, J.P.; Heade, J.; Ryan, S.M.; Brayden, D.J. Stability, toxicity and intestinal permeation enhancement of two food-derived antihypertensive tripeptides, Ile-Pro-Pro and Leu-Lys-Pro. Peptides 2015, 71, 1–7. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Brayden, D.J.; Ryan, S.M. Evaluation of PepT1 transport of food-derived antihypertensive peptides, Ile-Pro-Pro and Leu-Lys-Pro using in vitro, ex vivo and in vivo transport models. Eur. J. Pharm. Biopharm. 2017, 115, 276–284. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Frías, J.M.; Ryan, S.M.; Brayden, D.J. Sodium caprate enables the blood pressure-lowering effect of Ile-Pro-Pro and Leu-Lys-Pro in spontaneously hypertensive rats by indirectly overcoming PepT1 inhibition. Eur. J. Pharm. Biopharm. 2018, 128, 179–187. [Google Scholar] [CrossRef] [Green Version]
- de Vos, P.; Faas, M.M.; Spasojevic, M.; Sikkema, J. Encapsulation for preservation of functionality and targeted delivery of bioactive food components. Int. Dairy J. 2010, 20, 292–302. [Google Scholar] [CrossRef]
- Mohan, A.; Rajendran, S.R.C.K.; He, Q.S.; Bazinet, L.; Udenigwe, C.C. Encapsulation of food protein hydrolysates and peptides: A review. RSC Adv. 2015, 5, 79270–79278. [Google Scholar] [CrossRef]
- Yu, T.; Zhao, S.; Li, Z.; Wang, Y.; Xu, B.; Fang, D.; Wang, F.; Zhang, Z.; He, L.; Song, X.; et al. Enhanced and extended anti-hypertensive effect of VP5 nanoparticles. Int. J. Mol. Sci. 2016, 17, 1977. [Google Scholar] [CrossRef]
- Kecel-Gündüz, S.; Budama-Kilinc, Y.; Cakir Koc, R.; Kökcü, Y.; Bicak, B.; Aslan, B.; Özel, A.E. Computational design of Phe-Tyr dipeptide and preparation, characterization, cytotoxicity studies of Phe-Tyr dipeptide loaded PLGA nanoparticles for the treatment of hypertension. J. Biomol. Struct. Dyn. 2018, 36, 2893–2907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Su, M.; Jiang, X.X.; Xue, Y.Q.; Zhang, J.X.; Zeng, X.Q.; Wu, Z.; Guo, Y.X.; Pan, D.D. Transepithelial transport route and liposome encapsulation of milk-derived ACE-inhibitory peptide Arg-Leu-Ser-Phe-Asn-Pro. J. Agric. Food Chem. 2019, 67, 5544–5551. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.-C.; Cheng, M.-L.; Hsu, K.-C.; Hwang, J.-S. Absorption-enhancing treatments for antihypertensive activity of oligopeptides from tuna cooking juice: In vivo evaluation in spontaneously hypertensive rats. J. Food Sci. 2006, 71, S13–S17. [Google Scholar] [CrossRef]
- Hwang, J.S.; Tsai, Y.L.; Hsu, K.C. The feasibility of antihypertensive oligopeptides encapsulated in liposomes prepared with phytosterols-β-sitosterol or stigmasterol. Food Res. Int. 2010, 43, 133–139. [Google Scholar] [CrossRef]
- Li, Z.; Paulson, A.T.; Gill, T.A. Encapsulation of bioactive salmon protein hydrolysates with chitosan-coated liposomes. J. Funct. Foods 2015, 19, 733–743. [Google Scholar] [CrossRef]
- Montero, P.; Mosquera, M.; Marin-Penalver, D.; Aleman, A.; Martinez-Alvarez, O.; Gomez-Guillen, M.C. Changes in structural integrity of sodium caseinate films by the addition of nanoliposomes encapsulating an active shrimp peptide fraction. J. Food Eng. 2019, 244, 47–54. [Google Scholar] [CrossRef]
- Zhao, S.; Li, J.; Zhou, Y.; Huang, L.; Li, Y.; Xu, J.; Fu, C.; Guo, X.; Yang, J. Lipid nanoparticles-encapsulated YF4: A potential therapeutic oral peptide delivery system for hypertension treatment. Front. Pharmacol. 2019, 10, 102. [Google Scholar] [CrossRef]
- Gong, K.J.; Shi, A.M.; Liu, H.Z.; Liu, L.; Hu, H.; Yang, Y.; Adhikari, B.; Wang, Q. Preparation of nanoliposome loaded with peanut peptide fraction: Stability and bioavailability. Food Funct. 2016, 7, 2034–2042. [Google Scholar] [CrossRef]
- Mosquera, M.; Gimenez, B.; Montero, P.; Gomez-Guillen, M.C. Incorporation of liposomes containing squid tunic ACE-inhibitory peptides into fish gelatin. J. Sci. Food Agric. 2016, 96, 769–776. [Google Scholar] [CrossRef]
- Auwal, S.M.; Zarei, M.; Tan, C.P.; Basri, M.; Saari, N. Enhanced physicochemical stability and efficacy of angiotensin I-converting enzyme (ACE)-inhibitory biopeptides by chitosan nanoparticles optimized using Box-Behnken design. Sci. Rep. 2018, 8, 11. [Google Scholar] [CrossRef]
- Auwal, S.M.; Zarei, M.; Tan, C.P.; Basri, M.; Saari, N. Improved in vivo efficacy of anti-hypertensive biopeptides encapsulated in chitosan nanoparticles fabricated by ionotropic gelation on spontaneously hypertensive rats. Nanomaterials 2017, 7, 421. [Google Scholar] [CrossRef] [PubMed]
- Auwal, S.M.; Zarei, M.; Tand, C.P.; Saari, N. Comparative physicochemical stability and efficacy study of lipoid S75-biopeptides nanoliposome composite produced by conventional and direct heating methods. Int. J. Food Prop. 2018, 21, 1646–1660. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Mascaraque, L.G.; Miralles, B.; Recio, I.; Lopez-Rubio, A. Microencapsulation of a whey protein hydrolysate within micro-hydrogels: Impact on gastrointestinal stability and potential for functional yoghurt development. J. Funct. Foods 2016, 26, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.-J.; Mao, X.-Y.; Wang, Q.; Yang, S.; Zhang, D.; Chen, S.-W.; Li, Y.-H. Effect of spray drying and freeze drying on the immunomodulatory activity, bitter taste and hygroscopicity of hydrolysate derived from whey protein concentrate. LWT-Food Sci. Technol. 2014, 56, 296–302. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Manzanares, P.; Martínez, R.; Garrigues, S.; Genovés, S.; Ramón, D.; Marcos, J.; Martorell, P. Tryptophan-containing dual neuroprotective peptides: Prolyl endopeptidase inhibition and Caenorhabditis elegans protection from β-Amyloid Peptide Toxicity. Int. J. Mol. Sci. 2018, 19, 1491. [Google Scholar] [CrossRef]
- Muñoz, A.; Gandía, M.; Harries, E.; Carmona, L.; Read, N.D.; Marcos, J.F. Understanding the mechanism of action of cell-penetrating antifungal peptides using the rationally designed hexapeptide PAF26 as a model. Fungal Biol. Rev. 2013, 26, 146–155. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Milk proteins as a source of tryptophan-containing bioactive peptides. Food Funct. 2015, 6, 2115–2127. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Table | Predicted/Novel Sequence 1 | Methodology | Evaluation | Ref. |
|---|---|---|---|---|
| ACE inhibition | FW, WW, YW, VRF, IKP, LRW, LRF | QSAR modeling | In vitro IC50 | [35] |
| CW, TW, HW, QW, CY | QSAR modeling | In vitro IC50 | [36] | |
| DW, WP | Molecular docking | In vitro IC50 | [37] | |
| KW, VW, MKP, FAP, VAP, | Molecular docking | In silico IC50 | [38] | |
| IPP, VPP, LRP, IVY | ||||
| WCW, IWW, WWW, WWI, WLW | Tripeptide library, molecular docking | In vitro IC50 | [39] | |
| VKW, YAW, KYW, TAW | Rational design | In vitro IC50, cell toxicity | [40] | |
| IVP, INP, IQP, VIP | QSAR modeling | In vitro IC50, SHRs | [41] | |
| VPPIPP, IPPVPP | Rational design | In vitro IC50, SHRs | [42] | |
| GEF, VEF, VRF, VKF | QSAR modeling, molecular docking | In vitro IC50 | [43] | |
| RKWHFW, RKWLFW | Partial hexapeptide library | In vitro IC50, vasoconstriction, SHRs | [44] | |
| RKWHFLW | Rational design | In vitro IC50, vasoconstriction, | [45] | |
| SHRs, toxicity | ||||
| LHLPGP, LHLPLR | Rational design | In vitro IC50 | [46] | |
| Renin inhibition | IW, LW, VW, AW | QSAR modeling | In vitro renin activity | [47] |
| ACE & renin | RYLP, YTAWVP, YRAWVL | QSAR modeling, molecular dynamics, | In vitro IC50 | [48] |
| inhibition | peptide binding free energy |
| Table | Sequence 1 | Methodology | Evaluation | Ref. |
|---|---|---|---|---|
| DPP-IV inhibition | WP, WA, WR | XP and XA library | In vitro inhibitory effect | [70] |
| WRE | WRX library | In vitro inhibitory effect | [71] | |
| TH, NH, VL, | Dipeptide library | In vitro inhibitory effect | [72] | |
| ML, MM | ||||
| WP, YP | Sequence alignment | In vitro inhibitory effect | [73] | |
| α-glucosidase | SVPA, SEPA | Tri-tetra- and | In vitro IC50 | [77] |
| inhibition | pentapeptide library | |||
| DPP-IV, α-glucosidase, | AKSPLF, QTPF, | Computational docking | In vitro inhibitory effect | [78] |
| α-amylase imhibition | FEELN, LSKSVL | analysis |
| Sequence 1 | Methodology | Evaluation | Ref. |
|---|---|---|---|
| PHH | LLPHH-related peptides | Activity against peroxidation of linoleic acid | [82] |
| YHY, XXW, | Tripeptide library, | Activity against peroxidation of linoleic acid, | [84,85] |
| XXY, XXC | QSAR modeling | reducing activity, radical and peroxynitrite | |
| scavenging activity, Trolox equivalent | |||
| antioxidant capacity (TEAC), ferric | |||
| reducing antioxidant activity | |||
| ECH, YECG | Rational design | Radical scavenging activity, reducing power, | [86] |
| activity against peroxidation of linoleic acid, | |||
| oxygen radical absorbance capacity (ORAC), | |||
| TEAC, protection on H2O2-induced cytotoxicity | |||
| YX, XY, | Library of Y-, W-, C- | Radical scavenging activities, reducing power, | [87] |
| WX, XW | or M-containing dipeptides, | iron chelating activity, protective effect on | |
| QSAR modeling | erythrocyte hemolysis | ||
| YGY, YGGY, | Rational design | Antioxidant activities against hypochlorite ion, | [88] |
| GYYG, GWWW | hydroxyl radical, peroxynitrite | ||
| QP, PY | Rational design | Radical scavenging activity, iron chelating | [18] |
| activity, ORAC, cell response studies | |||
| ECRMR | Rational design, | Radical scavenging activity | [89] |
| 3D-QSAR modeling |
| Delivery Strategy | Peptide (Origin)/Hydrolysate | Bioactivity | Evaluated Functionality | In vitro/In vivo Model | Ref. |
|---|---|---|---|---|---|
| Sodium caprate | VPP (milk) and LKP (chicken, fish) | Antihypertensive | Intestinal permeability, antihypertensive effect | Rat jejunal tissue, plasma levels, SHRs | [95,96,97] |
| PLGA-based nanoparticles | VLPVP (synthetic) | Antihypertensive | Antihypertensive effect | SHRs | [100] |
| FY (seaweed) | Antihypertensive | Peptide toxicity | Fibroblast cells | [101] | |
| Liposomes | RLSFNP (milk) | ACE-inhibitory | Intestinal transport | Caco-2 cells | [102] |
| Tuna cooking juice oligopeptides | Antihypertensive | Antihypertensive effect | SHRs | [103,104] | |
| Chitosan coated liposomes | Salmon protein hydrolysate | Antidiabetic | In vitro release | Simulated biological fluids | [105] |
| Liposomes in sodium caseinate films | Shrimp peptide fraction | Antioxidant, ACE- and DPP-IV inhibitory | Solubility, palatability | Sensory evaluation | [106] |
| Nanoliposomes | YGLF (milk) | Antihypertensive | In vitro release, antihypertensive effect | SHRs | [107] |
| Peanut peptide fraction | ACE inhibitory | In vitro release, stability, bioavailability | Gastrointestinal digestion | [108] | |
| Nanoliposomes in fish gelatin | Squid tunic hydrolysate | ACE inhibitory | Stability, ACE inhibition | In vitro ACE inhibition | [109] |
| Nanoliposomes & chitosan nanoparticles | Stone fish-derived peptides | ACE inhibitory | In vitro release, stability, ACE inhibition, antihypertensive effect | Gastrointestinal digestion, SHRs | [110,111,112] |
| Microencapsulation in gelatin and chitosan | Whey protein hydrolysate | ACE-, DPP-IV inhibitory, hypocholesterolemic, antimicrobial | Bioaccesibility, stability | Gastrointestinal digestion, fermentation | [113] |
| Microencapsulation in sodium alginate and whey protein concentrate | Whey protein hydrolysate | Immunomodulatory | Immunomodulation, bitterness, hygroscopicity | In vitro splenocyte proliferation | [114] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzanares, P.; Gandía, M.; Garrigues, S.; Marcos, J.F. Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies. Nutrients 2019, 11, 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102545
Manzanares P, Gandía M, Garrigues S, Marcos JF. Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies. Nutrients. 2019; 11(10):2545. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102545
Chicago/Turabian StyleManzanares, Paloma, Mónica Gandía, Sandra Garrigues, and Jose F. Marcos. 2019. "Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies" Nutrients 11, no. 10: 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11102545