1. Introduction
Leptin is an adipokine produced by adipocytes that influences food intake, satiety, body weight, and metabolic activity [
1]. Sexual dimorphism in leptin has been observed in rodents and humans, with females having more leptin than males [
2,
3]. Females secrete more leptin than males and this difference in leptin secretion is intensified with obesity [
4]. Besides its role in food intake control, leptin has also been implicated in many other processes, including cardiovascular risk [
4,
5], cancer [
6,
7], neurodegenerative diseases [
8,
9], and in inflammation and immunity [
10,
11]. For all those reasons, it is especially interesting to delve deeper into the molecular bases that may influence plasma leptin concentrations, as, despite the considerable progress made, they are still not fully understood. Moreover, insufficient attention has often been paid to studying sex-specific effects, which may have resulted in several biases in the estimates, hence the need to reconsider those effects.
The leptin gene (LEP), initially called “ob” in mice [
12], is located on chromosome 7 in humans (7q32.1) and consists of three exons [
13,
14]. Although several low-frequency mutations associated with very low leptin levels, and extreme obesity phenotypes [
15,
16] have been identified, the effect of common polymorphisms on circulating leptin levels and obesity phenotypes is less clear and inconsistent [
17,
18,
19,
20,
21,
22]. This is because regulation is highly complex and it is known that other genes may also influence leptin levels, as well as on the effect of leptin on obesity and related diseases [
23]. Among those genes, the most studied one is the so-called leptin receptor gene (LEPR). Leptin binds to a membrane protein, the leptin receptor that is a class I cytokine receptor. Just as with the leptin gene, several low-frequency variants have been reported in the LEPR gene that result in LEPR deficiency and are associated with severe early-onset obesity and hyperphagia [
24,
25]. Likewise, common polymorphisms in the LEPR gene have been less consistently associated with plasma leptin levels and obesity phenotypes, depending on the type of population analyzed [
26,
27,
28,
29]. In many studies, the polymorphisms in the LEP and LEPR genes are analyzed jointly [
30,
31,
32,
33,
34].
Other candidate genes that may influence plasma leptin levels have also been identified [
35], and genome-wide association studies (GWAS) provide an efficient tool for revealing the main genes associated with this phenotype. However, despite the advantages of this approach, there have been very few GWAS published that have analyzed circulating leptin [
36,
37]. Outstanding among those is the GWAS and GWAS meta-analysis carried out by Kilpeläin et al. [
37], who undertook a two-stage study. In Stage 1, they carried out a GWAS of circulating leptin levels from 32,161 individuals (including 23 studies). In Stage 2, they then tested the loci with
p < 1 × 10
−6, in 19,979 additional individuals. Finally, in the joint meta-analysis for men and women, they identified five SNPs reaching genome-wide significance (
p < 5 × 10
−8). These SNPs were in/near the following genes: LEP, SLC32A1 (solute carrier family 32 member 1), GCKR (glucokinase regulatory protein), CCNL1 (cyclin-L1), and FTO (fat mass and obesity-associated). In addition to these five robustly associated SNPs, the authors also included the COBLL1 (cordon-bleu WH2 repeat protein like 1) gene to the list of relevant leptin-associated loci from the GWAS. The SNP in this gene was close to genome-wide significance, but previous GWAS studies reported strong associations between this locus and obesity-related measures.
However, it is becoming increasingly important to undertake GWAS on different populations, as the most relevant SNPs in one population may not be the most important in another and so give rise to bias [
38]. Likewise, the genetic risk scores (GRS) derived from the top-ranked SNPs identified in a GWAS undertaken on certain populations may not be useful in other populations not included or little represented in the initial GWAS [
39]. The Spanish Mediterranean population is, in general, little represented in GWAS, including those carried on European populations, and so specific studies must be undertaken on that population before directly using the SNPs or GRS derived from those GWAS. Specifically, in the case of leptin levels, various studies on different populations [
40,
41] are using the SNPs reported in the Kilpeläin et al. [
37] GWAS as plasma leptin level proxies for Mendelian randomization studies. Therefore, the aims of our study for the whole population and by sex are as follows: (1) to estimate the association between the SNPs reported as most relevant (in/near the LEP, SLC32A1, GCKR, CCNL, COBLL1, and FTO genes) in the Kilpeläin et al. [
37] GWAS and leptin levels in a Mediterranean population; (2) to investigate whether other SNPs (after the whole gene-screening) in the 6 genes reported by Kilpeläin et al. [
37], plus one that we have added—the LEPR gene—are more highly associated with leptin levels in this population; and (3) to undertake a new GWAS for plasma leptin levels in this Mediterranean population.
4. Discussion
In this study, we have investigated the SNPs most associated with plasma leptin concentrations in a high cardiovascular risk Mediterranean population. We have shown that there are several sex-specific genetic associations, as well as several population-specific-differences, concerning the central genes and SNPs that are most significantly associated. These results contribute, on one hand, to emphasizing the essential need to study the homogeneity or heterogeneity per sex of genetic associations (in this case, the SNPs associated with leptin levels) in order to obtain fewer biased results; thus contributing to more profound knowledge for future precision medicine [
62,
63]. On the other hand, these results also underscore the importance of studying specific populations instead of extrapolating results of gene variants obtained in studies undertaken in other populations [
38,
64,
65,
66,
67]. Several publications have pointed out that, for precision medicine, disease risk is likely to be miscalculated if GWAS results obtained in one population are naively used to compute GRS for a geographically different population [
38,
66]. It has been pointed out that most GWAS have been undertaken using European ancestry samples [
39]; however, even those GWAS have mainly been conducted in populations based in the United States, Germany, England, Finland, Sweden, Denmark, France, Austria, or Italy, among others [
39,
66,
67], and very few GWAS have analyzed the Spanish Mediterranean population. That means that information is unavailable on whether the gene variants that are associated to a higher degree in the GWAS and meta-analysis of GWAS in other countries are also the most relevant for the Spanish Mediterranean population. Previous studies indicate that there may be substantial differences in the predictive value of gene variants depending on different populations [
39,
66]. In this regard, European north–south gradients have even been described for several polymorphisms and ancestry markers [
68,
69,
70,
71,
72,
73]. These small genetic differences may influence the top-ranked SNPs in the corresponding GWAS, requiring finer tuning for the genetic structure in precision medicine/nutrition. In the specific case of leptin, very few GWAS have been undertaken worldwide, given that it is not a measurement readily available in large epidemiological studies. Outstanding among the few GWAS that have been undertaken is that of Kilpeläin et al. [
37], who in stage 1 analyzed plasma leptin including up to 32,161 individuals of European descent from 23 studies (mainly from Europe and the United States), but none of those 23 included a Spanish population and neither did their Stage 2 meta-analysis, which analyzed up to 19,979 additional individuals of European descent from 13 studies. Therefore, it is not surprising that in our Spanish Mediterranean population, the effect of the six SNPs (rs10487505-LEP, rs6071166- SLC32A, rs780093- GCKR, rs900400-CCNL1, rs6738627-COBLL1, and rs8043757-FTO), reported as most significant by Kilpeläin et al. [
37], did not reach statistical significance in our whole population. By sex, only one SNP was statistically significant in women after the adjustment for BMI. However, when we carried out, in our Mediterranean population, a whole gene screening to capture a more complete set of SNPs in the same candidate genes (LEP, SLC32A, GCKR, CCNL1, COBLL1, and FTO), we did obtain several statistically significant association between other SNPs in these genes and plasma leptin concentrations. In addition to those genes, we included the LEPR gene that, although not showing up as relevant in the GWAS de Kilpeläin et al. [
37], has been reported in other studies as having significant associations of SNPs in the LEPR gene with circulating leptin levels [
24,
26,
33,
74]. In the LEPR screening, we also found several SNPs showing statistically significant associations with leptin levels in our population. These results are relevant because they highlight the pitfalls of using SNPs or GRS, derived from the above mentioned GWAS [
37], as proxies for plasma leptin concentrations for setting individual-level metrics of genetic risk in Mendelian randomization studies [
75], in this, and other populations (i.e., the SNPs obtained in the GWAS undertaken by Kilpeläin et al. [
37], have been used in Mendelian randomization studies for the association between leptin and Alzheimer’s disease [
40] or bone mineral density [
41]).
Another factor that could act independently, or jointly with geographical differences, is population characteristics. In our case, we did not study a general Mediterranean population, but an elderly, high cardiovascular risk population in which there may be factors that also modulate the association between the candidate SNPs [
37] and plasma leptin levels. These gene-health status-environment interactions should be additionally analyzed in other studies to better evaluate the genetic contribution. We must underscore that candidate SNPs in some studies may not be the most relevant in other studies with a population of different age, environment, or health status characteristics. This adds then a more dynamic component to the effect of SNPs on a phenotype, in this case, leptin, which may vary with age, sex, and population traits. As an example of this, we could point to the results of Helgeland et al. [
76], where they analyzed, at the GWAS level, the associations between the genome and BMI in children at 12 time points, from birth to eight years (9,286 children, more than 70,000 measurements) in a Norwegian cohort. Their results clearly show that the effect of an SNP on BMI is variable and either can show associations with BMI or not, depending on when it is measured. For example, for the SNPs in the LEPR, they identified a transient effect with no effect at birth, increasing effect in infancy, and slight effect after 5 years. A similar transient effect near the leptin gene was identified.
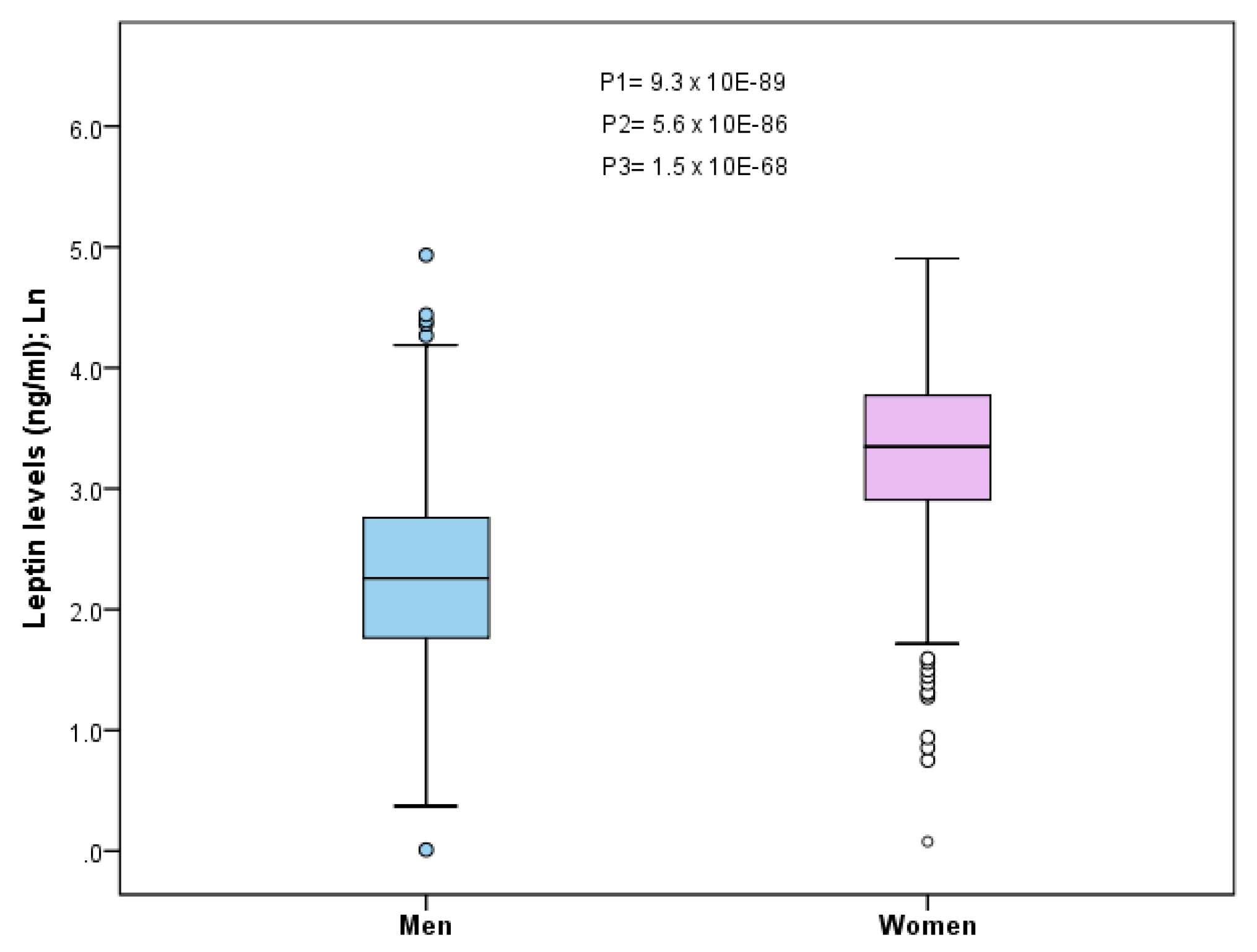
Therefore, in generating results for future precision medicine and nutrition, it will be necessary to pay more attention to the possible dynamic characteristics of SNP associations with the traits that determine them, as well as the characteristics of the populations analyzed. Of relevance will be the analysis of sex-specific effects [
62,
63,
77]. In the case of leptin, its levels are higher in women than in men at all ages (1–5). In our analysis of SNP*sex interactions, we found various SNPs in candidate genes with statistically significant interactions (at the nominal
p-value) with some LEPR and FTO SNPs. We also found sex-specific effects concerning the candidate genes/SNPs that are more significantly associated with plasma leptin levels. In men, the FTO gene presents the most significant associations, whereas, in women, it was found to be the LEPR gene. Currently, we do not know the potential mechanisms behind these sex-specific differences. Few studies have carried out formal analysis of the gene–sex interactions in determining leptin level and this information is lacking. There have been studies associating polymorphisms in the FTO gene with plasma leptin levels in various populations [
37,
78,
79,
80,
81,
82], concluding that leptin could be a possible intermediary contributing to the association between the FTO polymorphism and adiposity. However, those studies did not properly investigate potential sex-related differences. Therefore, sex-specific mechanisms need to be better studied.
In the GWAS analysis undertaken to reveal the SNPs most associated with plasma leptin levels in the Spanish Mediterranean population, we did not identify any association at the conventional GWAS level (5 × 10
−8). However, we identified some SNPs associated with plasma leptin levels at the suggestive GWAS level (1 × 10
−5) in the whole population as well as in men and women. Although our sample size was not large, this being a limitation, the homogeneity in the geographic origin as well as the homogeneity in the population characteristics and the duplicate measurements of leptin concentrations, increased statistical power. Furthermore, we can state that the percentage of variability explained by the top-ranked SNPs is very low. These data are in agreement with other GWAS studies [
37] and support the finding that the genetic associations obtained did not reach a great statistical significance, suggesting an important modulation by environmental factors and/or population characteristics. The additional adjustments for BMI in our models have allowed us to identify which SNPs seem to have an effect on leptin regardless of weight and those that are dependent. This adjustment was proposed by Kilpeläinen et al. [
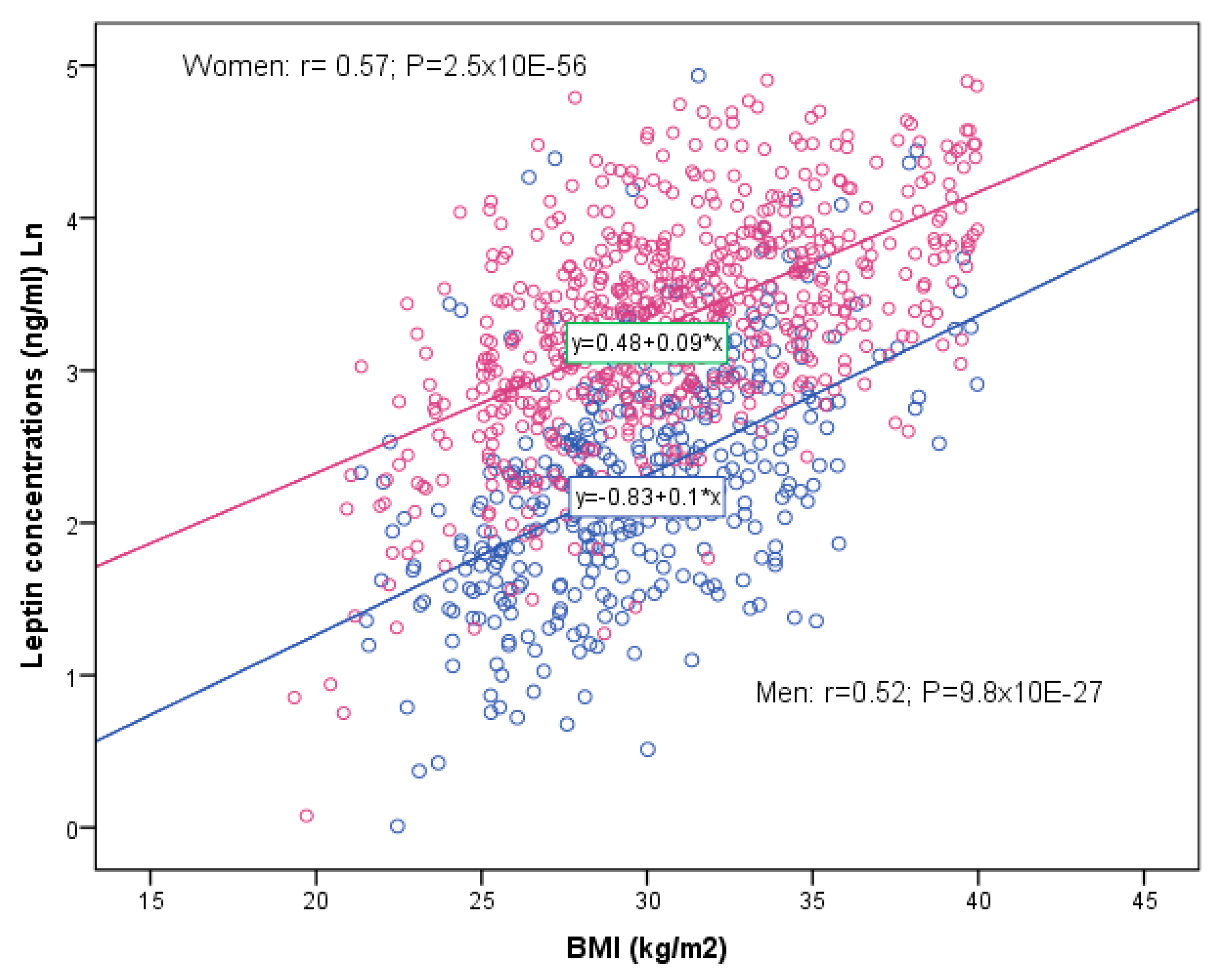
37], although they had already stated that one possible limitation may be that BMI is not the best measurement of adiposity. They undertook a secondary analysis in a subsample consisting of 13 studies that had data on both BMI and body fat percentage assessed by dual-energy X-ray absorptiometry or bioimpedance analysis. In that study, they were able to observe that the effects were similar to those adjusted for BMI or adjusting the models for body fat %, so suggesting that adjustment for BMI as compared with a more direct measure of adiposity did not compromise the ability to identify adiposity-independent leptin-associated loci. In our case, we did not have data on body fat % measurements available and so were unable to test the possible differential influence or not.
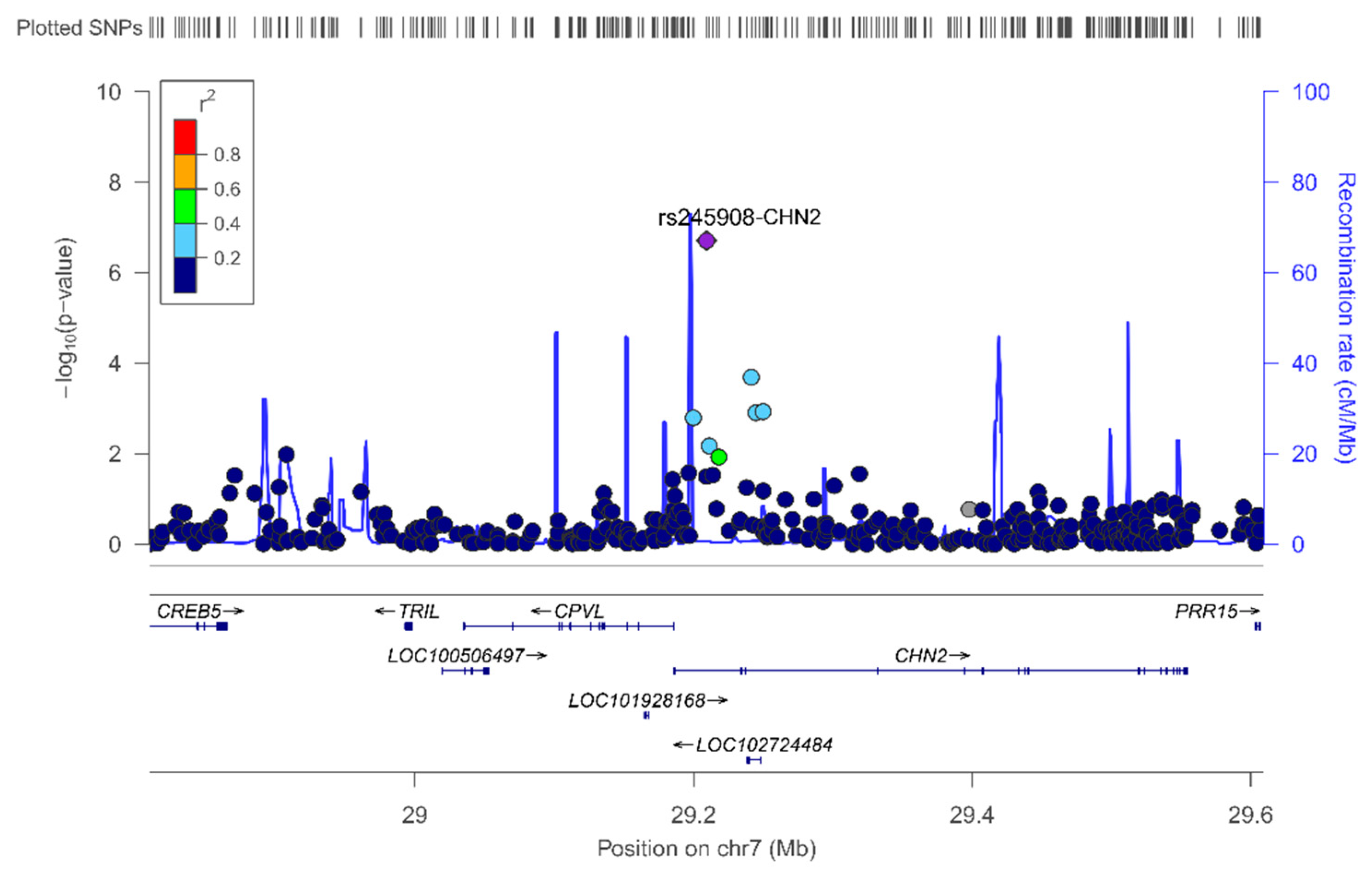
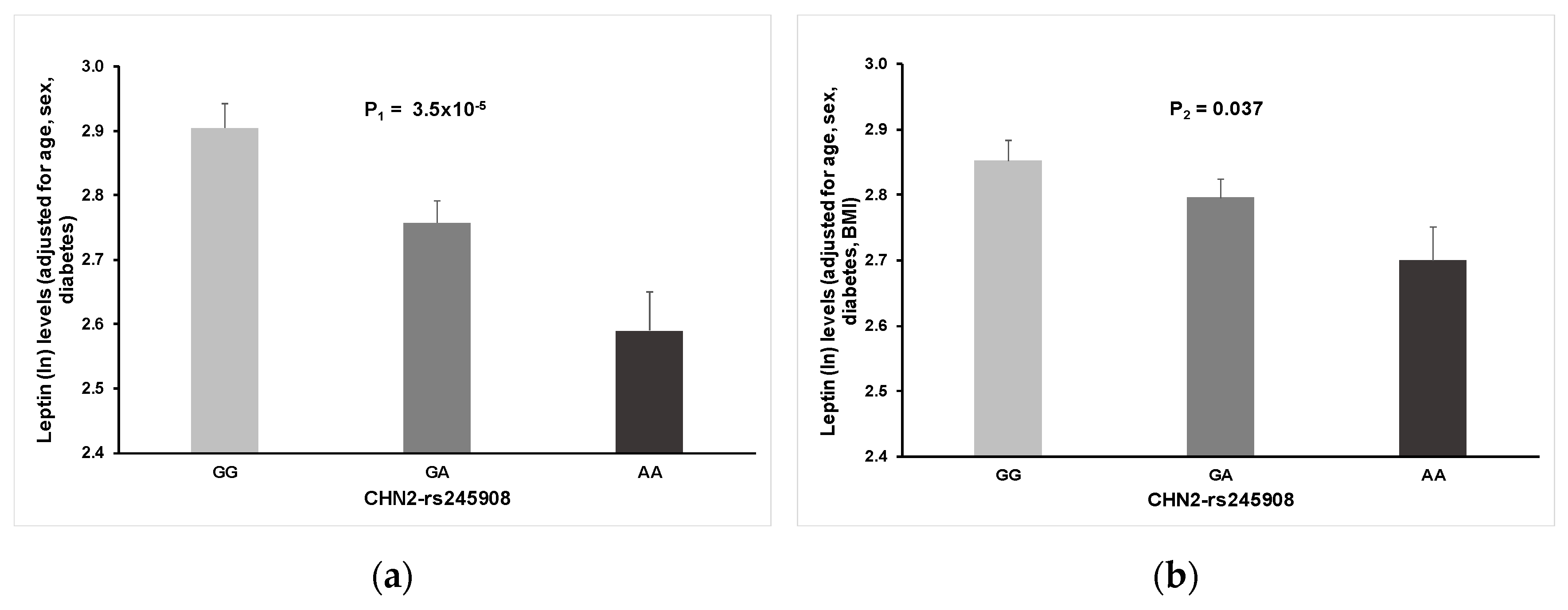
As far as we know, this is the first GWAS carried out on a Spanish Mediterranean population for the circulating leptin phenotype. Detailed analysis and some discussion of the top-ranked SNPs, both in the whole population and by sex, are provided in the results section (see Results for more information). We have outlined the novelty of our findings regarding the association between the rs245908-CHN2 SNP and plasma leptin concentration. The minor allele of this common variant (MAF: 0.38) is associated with lower leptin concentrations. This association, although attenuated, remained statistically significant after BMI adjustment. Curiously, it was observed for both men and women in the stratified analysis (not shown). Previous studies reported associations between the CHN2 gene and insulin-resistance, diabetes, and obesity phenotypes [
50,
51,
83] as well as with addiction and neurological diseases [
84,
85,
86].
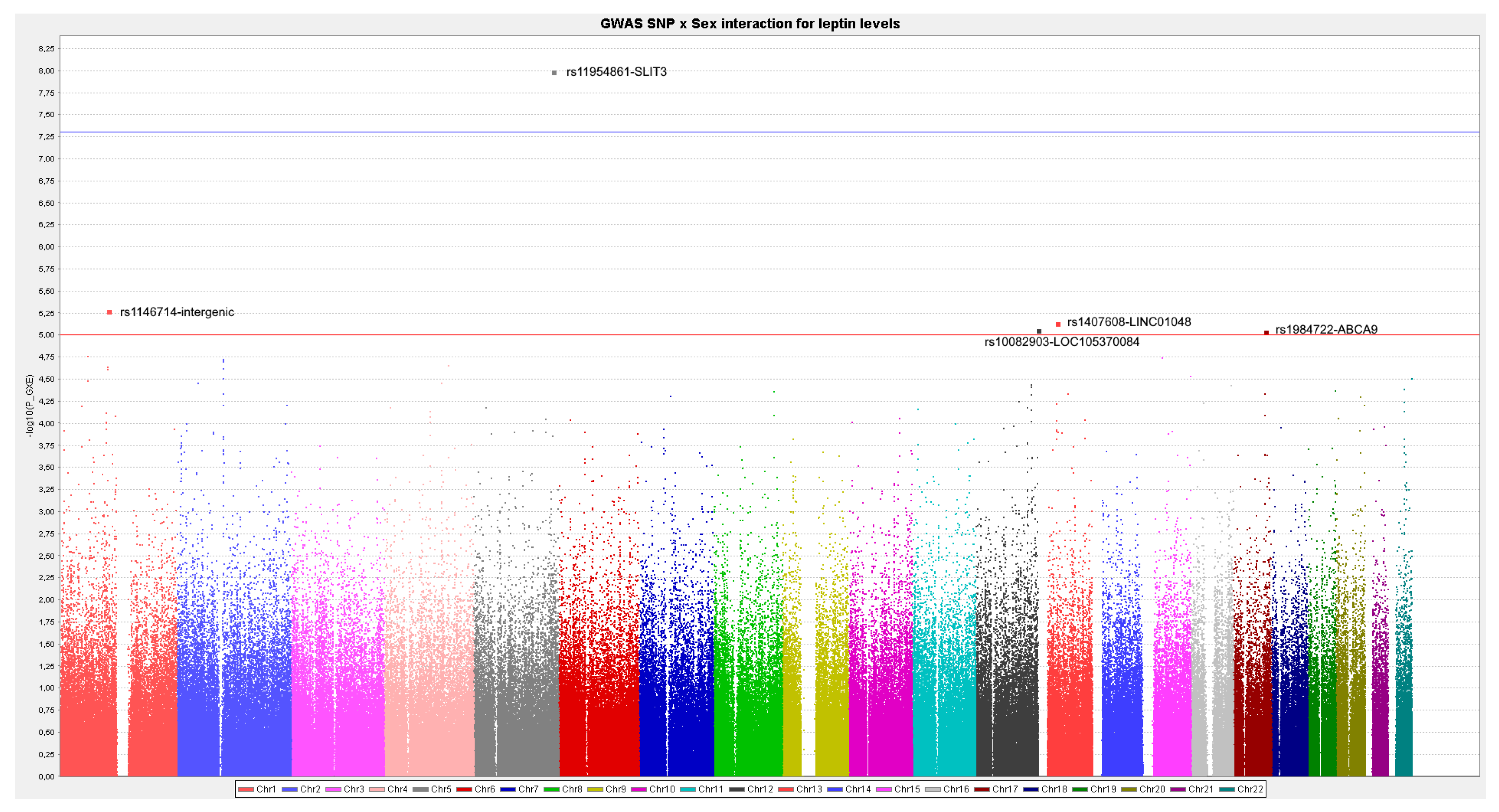
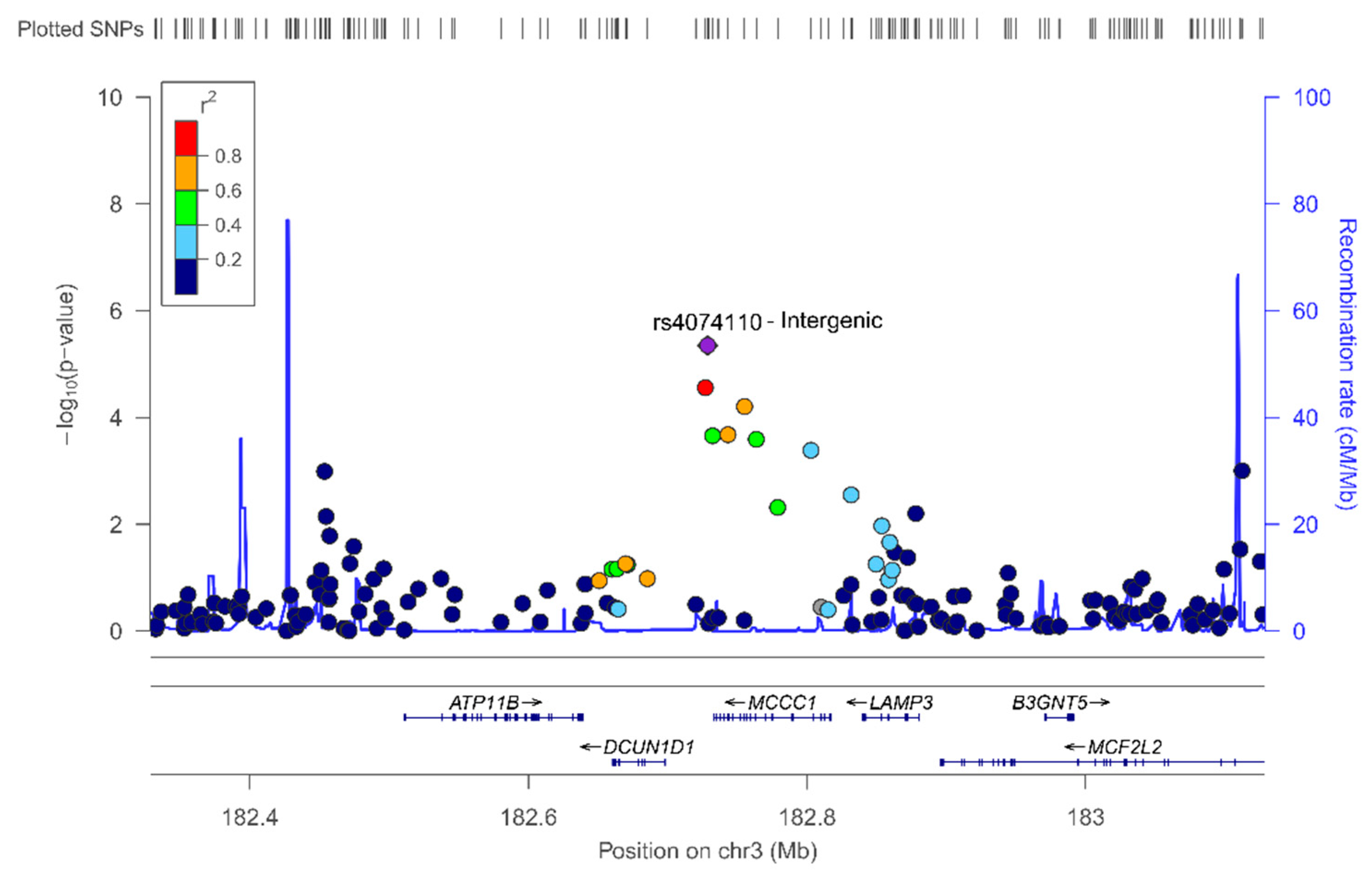
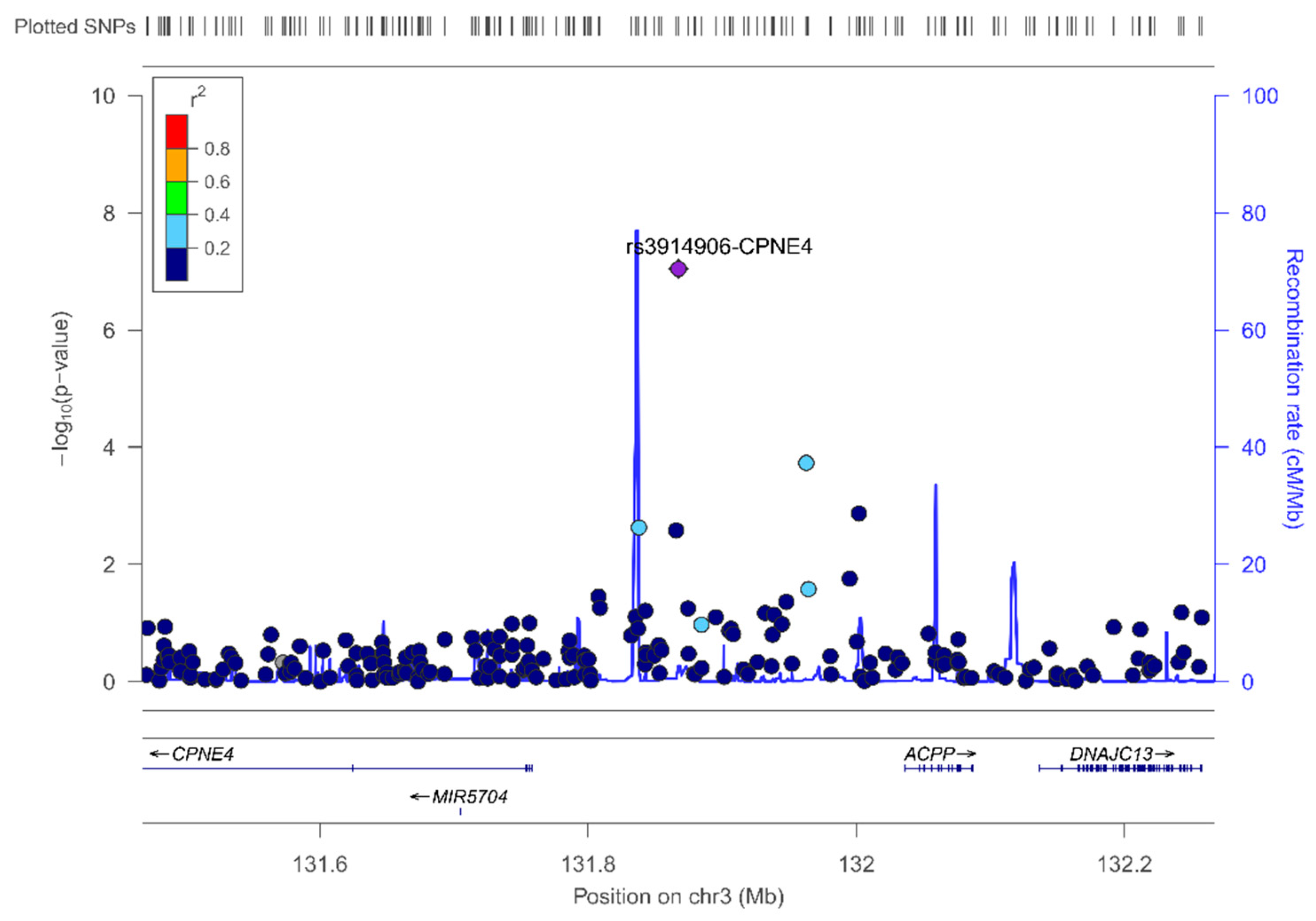
In the sex-stratified GWAS for men and women, we have identified novel sex-specific associations with plasma leptin. Hence, we identified a suggestive male-specific loci at chromosome 3 involving the SNP rs4074110 (intergenic) near the methylcrotonoyl-CoA carboxylase 1 (MCCC1) and defective in cullin neddylation 1 domain containing 1 (DCUN1D1) genes. Likewise, we identified a suggestive female-specific loci at chromosome 3 in the CPNE4 gene (rs3914096). Previous studies have associated this gene with cardiovascular and neurodegenerative phenotypes [
60,
61], but did not explore plasma leptin concentrations. Although these SNPs did not reach statistical significance at
p < 5 × 10
−8, they did reach a suggestive
p-value (
p = 3.28 × 10
−6 for the rs4074110 and
p = 9.04 × 10
−8 for the rs3914096), respectively. As GWAS on leptin concentrations are very scarce, the interest in showing these results lies in the fact that they will provide information to compare with other studies and for inclusion in meta-analyses. On analyzing the statistical significance of the gene–sex interaction terms at the genome-wide level, we detected several SNPs having sex-specific effects at
p < 1 × 10
−5. Moreover, we detected a SLIT3 gene–sex interaction at the GWAS level of significance (
p < 5 × 10
−8). Interestingly, SLIT3 is regulated by estrogens [
52]. The SLIT family interacts with the ROBO family of transmembrane receptors in a wide variety of physiological processes. SLIT3 is widely expressed in human tissues and its deregulation has been associated with cancer, including breast cancer [
87]. SLIT3 also has been related to several processes in the ovary including follicle development, among others [
52,
88,
89]. This gene presents a higher association with leptin concentrations in women than in men, and in opposite directions for the top-ranked SNP (rs11954861-SLIT3). Although the SLIT3–sex interaction in determining leptin levels is reported here for the first time, several studies have shown other sex-specific effects for this gene. Thus, Park et al. [
53], showed that Slit3-KO mice displayed anxiety-like behaviors, and these effects were mainly observed in female KO mice. Likewise, Chung et al. [
90], in a genome-wide interaction analysis between sex and SNPs on intracerebral hemorrhage risk, obtained a SNP*sex interaction at the GWAS level with an SNP in the SLIT3 gene (rs2337552;
p = 1.0 × 10
−8). In the sex-stratified association tests, in agreement with our results, the effect directions differed between men and women. Additional studies are required to check the consistency of these observations which, together with our study, will contribute to a better understanding of the genetics of leptin and subsequently for obesity-cardiovascular phenotypes and interactions with diet.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}