Nutritional and Pharmacological Targeting of the Calcium-Sensing Receptor Influences Chemically Induced Colitis in Mice

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Handling and Maintenance
2.2. Experimental Design and Induction of Colitis
2.3. Clinical Assessment of Colitis
2.4. Histology Scoring
2.5. Mucin Quantification
2.6. Cytokine Multiplex Assay
2.7. Immunohistochemistry
2.8. Quantification of Immune Cell Infiltration
2.9. GSK3004774 Synthesis
2.10. Intracellular Ca2+ Measurements to Test the Activity of GSK3004774
2.11. Statistical Analysis
3. Results
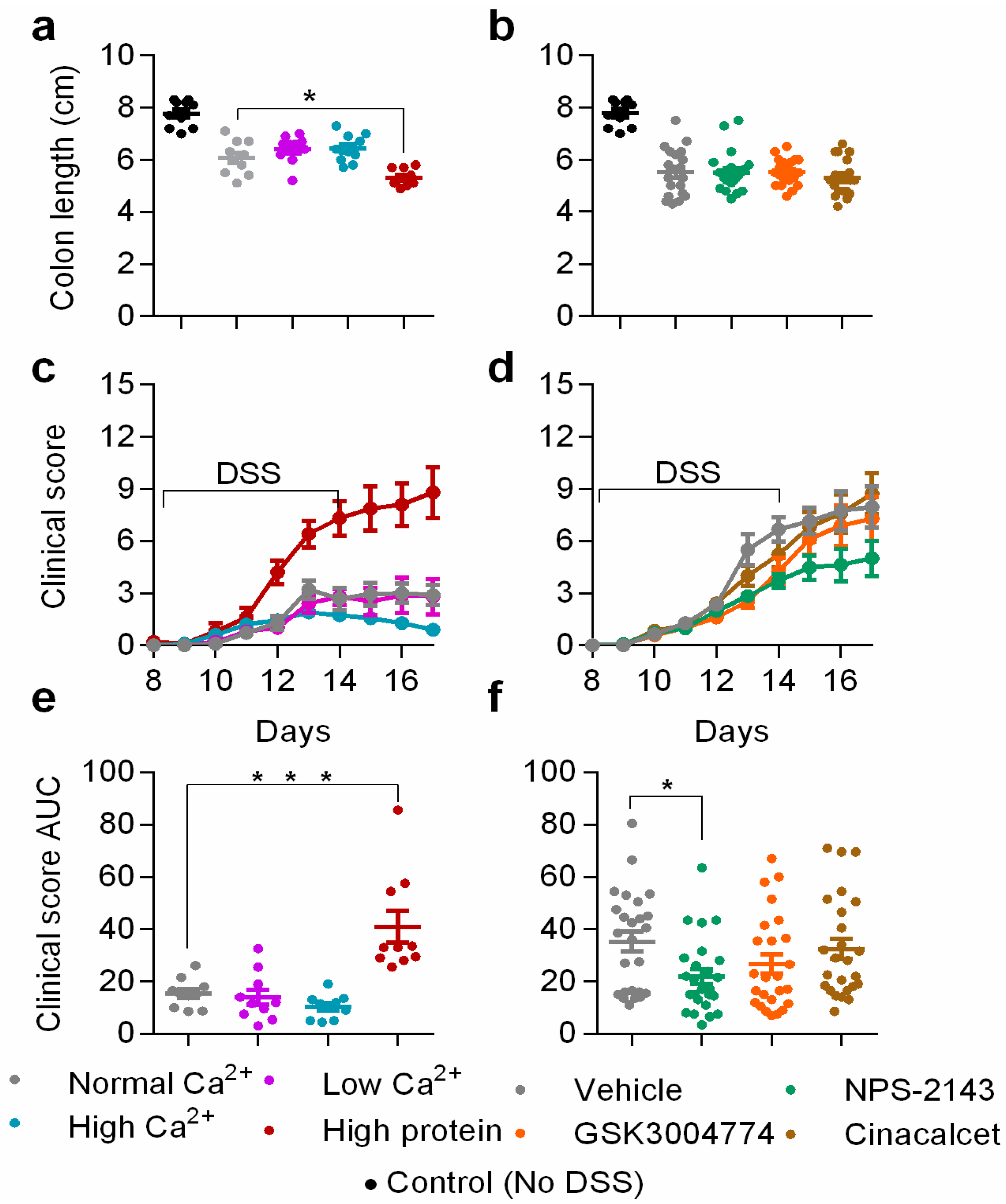
3.1. Clinical Symptoms of Colitis were Exacerbated by High Dietary Protein, but Improved by NPS-2143
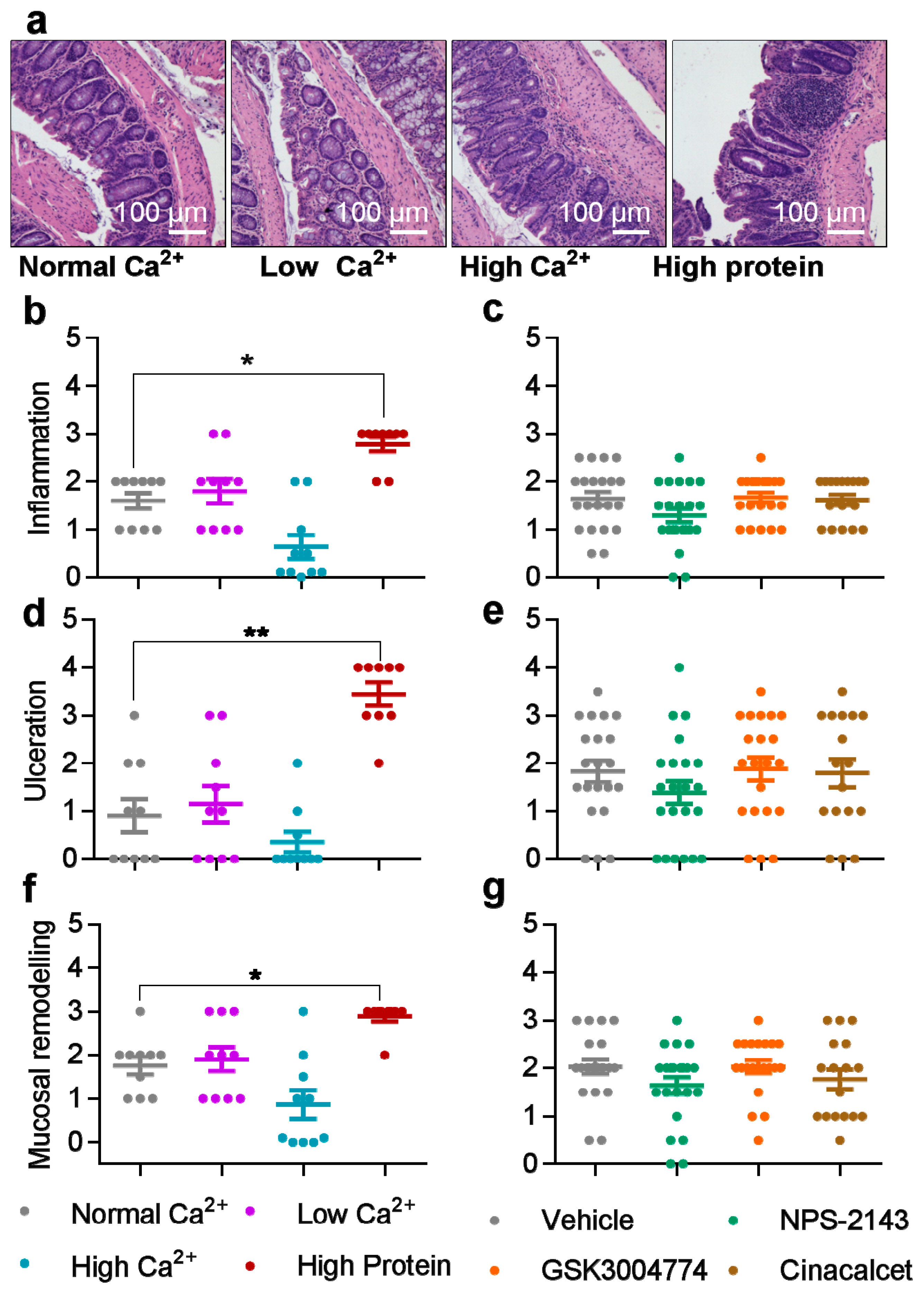
3.2. Histological Scores were Increased by High Dietary Protein, while Unaffected by Dietary Calcium and The Pharmacological Modulators of The CaSR
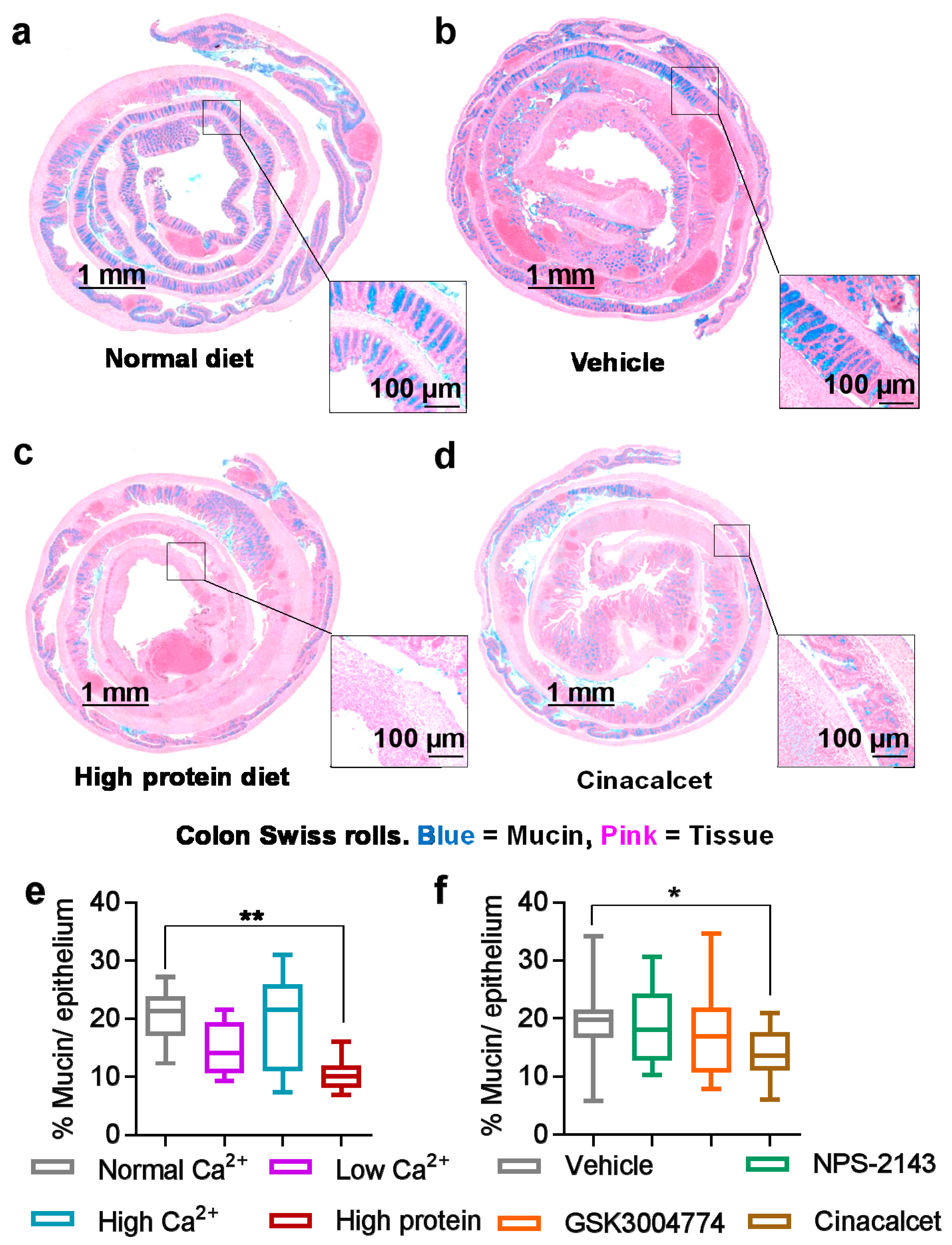
3.3. Mucin Was Reduced by High Dietary Protein and the Positive Allosteric Modulator Cinacalcet
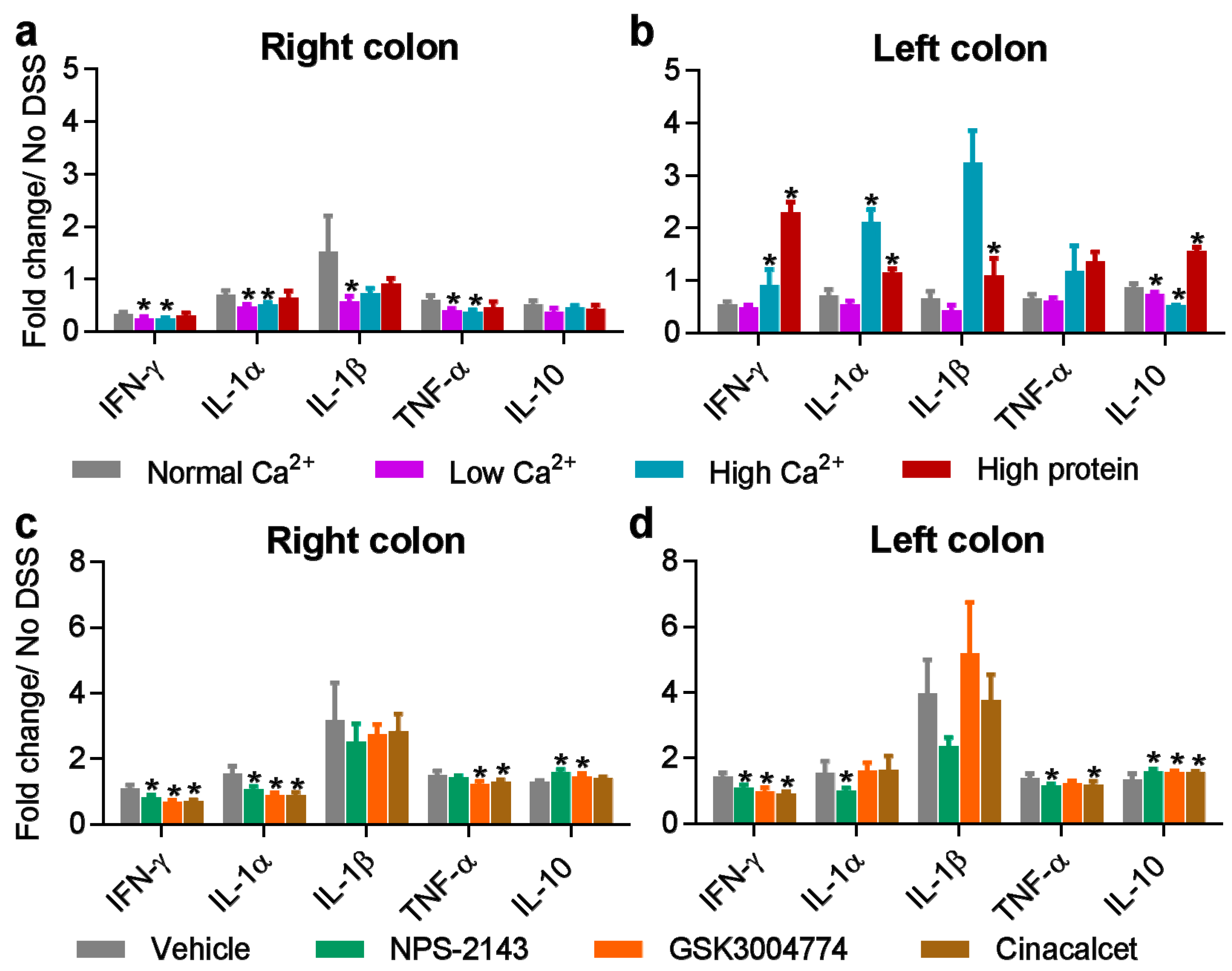
3.4. Inflammatory Cytokines are Differentially Affected by Nutritional and Pharmacological CaSR Ligands
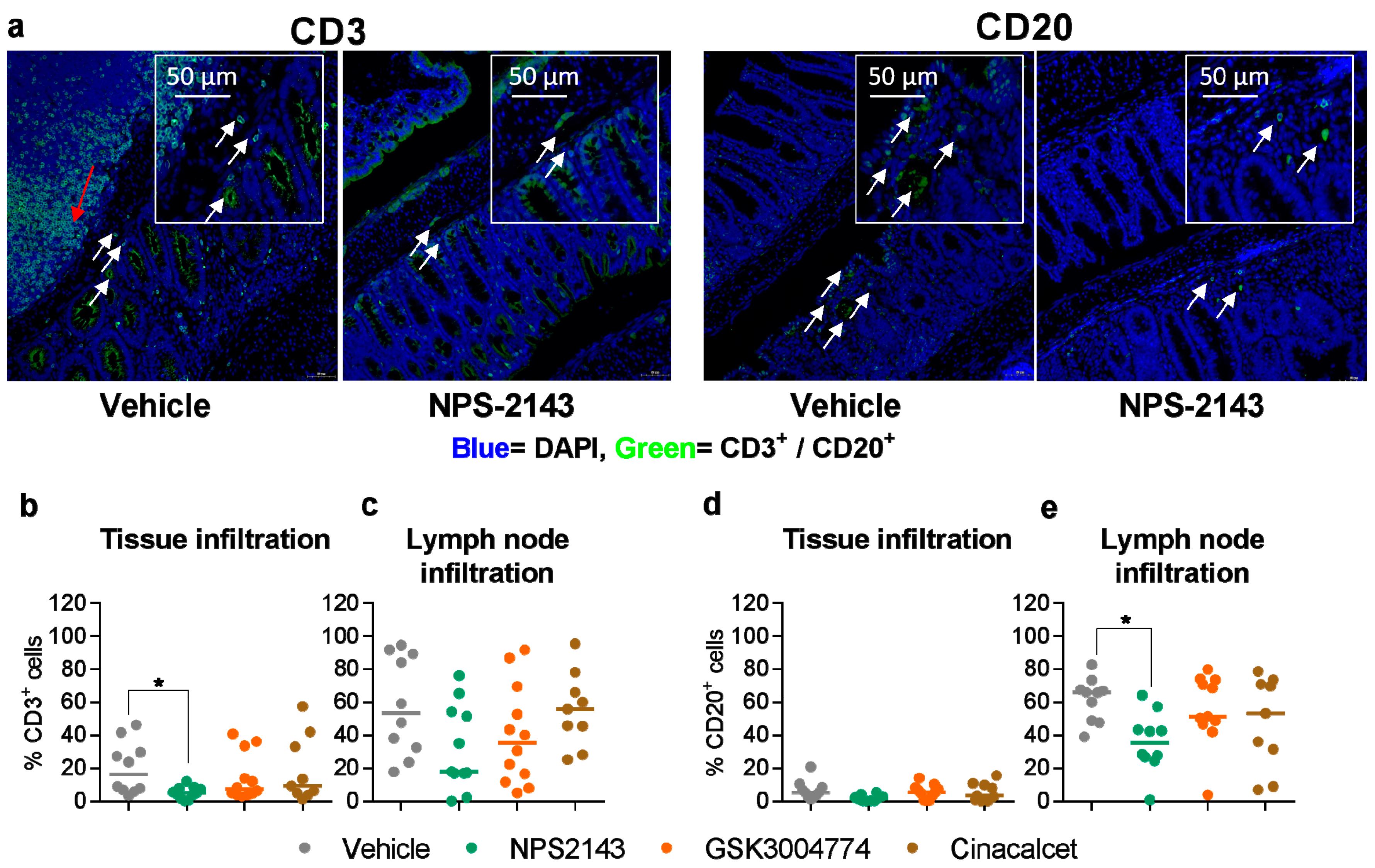
3.5. NPS-2143 Reduced Immune Cell Infiltration into the Colon
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hannan, F.M.; Kallay, E.; Chang, W.; Luisa Brandi, M.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. Nat. Rev. Endocrinol. 2018, 15, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, D.; Valenti, G. Localization and function of the renal calcium-sensing receptor. Nat. Rev. Nephrol. 2016, 12, 414–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conigrave, A.D. The calcium-sensing receptor and the parathyroid: Past, present, future. Front. Physiol. 2016, 7, 563. [Google Scholar] [CrossRef] [PubMed]
- Gerbino, A.; Colella, M. The Different Facets of Extracellular Calcium Sensors: Old and New Concepts in Calcium-Sensing Receptor Signalling and Pharmacology. Int. J. Mol. Sci. 2018, 19, 999. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Miller, C.L.; Gorkhali, R.; Zou, J.; Huang, K.; Brown, E.M.; Yang, J.J. Molecular Basis of the Extracellular Ligands Mediated Signaling by the Calcium Sensing Receptor. Front. Physiol. 2016, 7, 441. [Google Scholar] [CrossRef] [Green Version]
- Leach, K.; Conigrave, A.D.; Sexton, P.M.; Christopoulos, A. Towards tissue-specific pharmacology: Insights from the calcium-sensing receptor as a paradigm for GPCR (patho)physiological bias. Trends Pharmacol. Sci. 2015, 36, 215–225. [Google Scholar] [CrossRef]
- Leach, K.; Sexton, P.M.; Christopoulos, A.; Conigrave, A.D. Engendering biased signalling from the calcium-sensing receptor for the pharmacotherapy of diverse disorders. Br. J. Pharmacol. 2014, 171, 1142–1155. [Google Scholar] [CrossRef] [Green Version]
- Valenti, G.; Mira, A.; Mastrofrancesco, L.; Lasorsa, D.R.; Ranieri, M.; Svelto, M. Differential modulation of intracellular Ca2+ responses associated with calcium-sensing receptor activation in renal collecting duct cells. Cell. Physiol. Biochem. 2010, 26, 901–912. [Google Scholar] [CrossRef]
- Ranieri, M.; Zahedi, K.; Tamma, G.; Centrone, M.; Di Mise, A.; Soleimani, M.; Valenti, G. CaSR signaling down-regulates AQP2 expression via a novel microRNA pathway in pendrin and NaCl cotransporter knockout mice. FASEB J. 2018, 32, 2148–2159. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, M. Renal Ca2+ and water handling in response to calcium sensing receptor signaling: Physiopathological aspects and role of CaSR-regulated microRNAs. Int. J. Mol. Sci. 2019, 20, 5341. [Google Scholar] [CrossRef] [Green Version]
- Iamartino, L.; Elajnaf, T.; Kallay, E.; Schepelmann, M. Calcium-sensing receptor in colorectal inflammation and cancer: Current insights and future perspectives. World J. Gastroenterol. 2018, 24, 4119–4131. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Peng, M.; Liu, L.; Chang, W.; Binder, H.J.; Cheng, S.X. Calcium-sensing receptor stimulates Cl (-)- and SCFA-dependent but inhibits cAMP-dependent HCO3 (-) secretion in colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G874–G883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Cheng, C.Y.; Sun, X.; Pedicone, A.J.; Mohamadzadeh, M.; Cheng, S.X. The Extracellular Calcium-Sensing Receptor in the Intestine: Evidence for Regulation of Colonic Absorption, Secretion, Motility, and Immunity. Front. Physiol. 2016, 7, 245. [Google Scholar] [PubMed] [Green Version]
- Barahona, M.J.; Maina, R.M.; Lysyy, T.; Finotti, M.; Caturegli, G.; Baratta, V.; D’Amico, F.; Mulligan, D.; Geibel, J.P. Activation of the Calcium Sensing Receptor Decreases Secretagogue-Induced Fluid Secretion in the Rat Small Intestine. Front. Physiol. 2019, 10, 439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barahona, M.J.; Baratta, V.; Ollodart, J.; Mulligan, D.; Geibel, J.P. Design and implementation of novel nutraceuticals and derivatives for treating intestinal disorders. Future Med. Chem. 2019, 11, 847–855. [Google Scholar] [CrossRef]
- Yarova, P.L.; Stewart, A.L.; Sathish, V.; Britt, R.D.; Thompson, M.A.; Lowe, A.P.P.; Freeman, M.; Aravamudan, B.; Kita, H.; Brennan, S.C.; et al. Calcium-sensing receptor antagonists abrogate airway hyperresponsiveness and inflammation in allergic asthma. Sci. Transl. Med. 2015, 7, ra60. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.X.; Lightfoot, Y.L.; Yang, T.; Zadeh, M.; Tang, L.; Sahay, B.; Wang, G.P.; Owen, J.L.; Mohamadzadeh, M. Epithelial CaSR deficiency alters intestinal integrity and promotes proinflammatory immune responses. FEBS Lett. 2014, 588, 4158–4166. [Google Scholar] [CrossRef] [Green Version]
- Mine, Y.; Zhang, H. Anti-inflammatory Effects of Poly-L-lysine in Intestinal Mucosal System Mediated by Calcium-Sensing Receptor Activation. J. Agric. Food Chem. 2015, 63, 10437–10447. [Google Scholar] [CrossRef]
- Zhang, H.; Kovacs-Nolan, J.; Kodera, T.; Eto, Y.; Mine, Y. γ-Glutamyl cysteine and γ-glutamyl valine inhibit TNF-α signaling in intestinal epithelial cells and reduce inflammation in a mouse model of colitis via allosteric activation of the calcium-sensing receptor. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 792–804. [Google Scholar] [CrossRef] [Green Version]
- Low, D.; Nguyen, D.D.; Mizoguchi, E. Animal models of ulcerative colitis and their application in drug research. Drug Des. Dev. Ther. 2013, 7, 1341–1356. [Google Scholar]
- Perera, P.S.; Thompson, R.L.; Wiseman, M.J. Recent Evidence for Colorectal Cancer Prevention Through Healthy Food, Nutrition, and Physical Activity: Implications for Recommendations. Curr. Nutr. Rep. 2012, 1, 44–54. [Google Scholar] [CrossRef]
- Ahearn, T.U.; Mccullough, M.L.; Flanders, W.D.; Long, Q.; Sidelnikov, E.; Fedirko, V.; Daniel, C.R.; Rutherford, R.E.; Shaukat, A.; Bostick, R.M. A Randomized Clinical Trial of the Effects of Supplemental Calcium and Vitamin D 3 on Markers of Their Metabolism in Normal Mucosa of Colorectal Adenoma Patients. Prev. Epidemiol. 2011, 25, 413–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mine, Y.; Zhang, H. Calcium-Sensing Receptor (CaSR)-Mediated Anti-inflammatory Effects of l-Amino Acids in Intestinal Epithelial Cells. J. Agric. Food Chem. 2015, 63, 9987–9995. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tan, B.; Huang, B.; Li, J.; Wang, J.; Liao, P.; Guan, G.; Ji, P.; Yin, Y. Involvement of calcium-sensing receptor activation in the alleviation of intestinal inflammation in a piglet model by dietary aromatic amino acid supplementation. Br. J. Nutr. 2018, 120, 1321–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, O.; Young, S.H.; Jacamo, R.; Moyer, M.P.; Rozengurt, E. Extracellular calcium sensing receptor stimulation in human colonic epithelial cells induces intracellular calcium oscillations and proliferation inhibition. J. Cell. Physiol. 2010, 225, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, S.M.; Spearing, P.K.; Diaz, C.J.; Cowan, D.J.; Jayawickreme, C.; Chen, G.; Rimele, T.J.; Generaux, C.; Harston, L.T.; Roller, S.G. Identification of potent, nonabsorbable agonists of the calcium-sensing receptor for GI-specific administration. Bioorgan Med. Chem. Lett. 2017, 27, 4673–4677. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. In Current Protocols in Immunology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; Volume 104, pp. 15.25.1–15.25.14. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hall, M.; Frank, E.; Holmes, G.; Pfahringer, B.; Reutemann, P.; Witten, I.H. The WEKA data mining software. ACM SIGKDD Explor. Newsl. 2009, 11, 10–18. [Google Scholar] [CrossRef]
- Ruifrok, A.C.; Johnston, D.A. Quantification of histochemical staining by color deconvolution. Anal. Quant. Cytol. Histol. 2001, 23, 291–299. [Google Scholar]
- Miyazaki, H.; Tsubakimoto, J.; Yasuda, K.; Takamuro, I.; Sakurai, O.; Yanagida, T.; Hisada, Y. Preparation of Arylalkylamines as Calcium-Sensing Receptor (CaSR) Activators. WO2005115975A1, 8 December 2005. [Google Scholar]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating intestinal inflammation in DSS-induced model of IBD. J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepens, M.A.A.; Schonewille, A.J.; Vink, C.; Van Schothorst, E.M.; Kramer, E.; Hendriks, T.; Brummer, R.; Keijer, J.; Van Der Meer, R.; Bovee-oudenhoven, I.M.J. Supplemental Calcium Attenuates the Colitis-Related Increase in Diarrhea, Intestinal Permeability, and Extracellular Matrix Breakdown in HLA-B27 Transgenic Rats. J. Nutr. 2009, 139, 1525–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszak, I.T.; Poznansky, M.C.; Evans, R.H.; Olson, D.; Kos, C.; Pollak, M.R.; Brown, E.M.; Scadden, D.T. Extracellular calcium elicits a chemokinetic response from monocytes in vitro and in vivo. J. Clin. Investig. 2000, 105, 1299–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, A.; Blais, A.; Coelho, D.; Capron, J.; Maarouf, M.; Benamouzig, R.; Lancha, A.H.; Walker, F.; Tomé, D.; Blachier, F. Dual effects of a high-protein diet on DSS-treated mice during colitis resolution phase. Am. J. Physiol. Liver Physiol. 2016, 311, G624–G633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Lletjós, S.; Andriamihaja, M.; Blais, A.; Grauso, M.; Lepage, P.; Davila, A.-M.; Viel, R.; Gaudichon, C.; Leclerc, M.; Blachier, F.; et al. Dietary Protein Intake Level Modulates Mucosal Healing and Mucosa-Adherent Microbiota in Mouse Model of Colitis. Nutrients 2019, 11, 514. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, S.R.; Britton, G.J.; Contijoch, E.J.; Vennaro, O.H.; Mortha, A.; Colombel, J.F.; Grinspan, A.; Clemente, J.C.; Merad, M.; Faith, J.J. Interactions Between Diet and the Intestinal Microbiota Alter Intestinal Permeability and Colitis Severity in Mice. Gastroenterology 2018, 154, 1037–1046. [Google Scholar] [CrossRef]
- Morens, C.; Gaudichon, C.; Fromentin, G.; Marsset-Baglieri, A.; Bensaïd, A.; Larue-Achagiotis, C.; Luengo, C.; Tomé, D. Daily delivery of dietary nitrogen to the periphery is stable in rats adapted to increased protein intake. Am. J. Physiol. Metab. 2001, 281, E826–E836. [Google Scholar] [CrossRef]
- Gäbele, E.; Dostert, K.; Hofmann, C.; Wiest, R.; Schölmerich, J.; Hellerbrand, C.; Obermeier, F. DSS induced colitis increases portal LPS levels and enhances hepatic inflammation and fibrogenesis in experimental NASH. J. Hepatol. 2011, 55, 1391–1399. [Google Scholar] [CrossRef]
- Paccou, J.; Boudot, C.; Renard, C.; Liabeuf, S.; Kamel, S.; Fardellone, P.; Massy, Z.; Brazier, M.; Mentaverri, R. Total calcium-sensing receptor expression in circulating monocytes is increased in rheumatoid arthritis patients with severe coronary artery calcification. Arthritis Res. Ther. 2014, 16, 412. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.-S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The calcium-sensing receptor regulates the NLRP3 inflammasome through Ca2+ and cAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Petersen, C.D.; Coy, D.H.; Jiang, J.-K.; Thomas, C.J.; Pollak, M.R.; Wank, S.A. Calcium-sensing receptor is a physiologic multimodal chemosensor regulating gastric G-cell growth and gastrin secretion. Proc. Natl. Acad. Sci. USA 2010, 107, 17791–17796. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Amount (g/kg Diet) | |||
|---|---|---|---|---|
| Normal Ca2+ | Low Ca2+ | High Ca2+ | High Protein | |
| Casein (> or =85% protein) | 140.000 | 140.000 | 140.000 | 260.000 |
| Supplemental CaCO3 | 12.495 | 0.000 | 37.485 | 12.495 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elajnaf, T.; Iamartino, L.; Mesteri, I.; Müller, C.; Bassetto, M.; Manhardt, T.; Baumgartner-Parzer, S.; Kallay, E.; Schepelmann, M. Nutritional and Pharmacological Targeting of the Calcium-Sensing Receptor Influences Chemically Induced Colitis in Mice. Nutrients 2019, 11, 3072. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123072
Elajnaf T, Iamartino L, Mesteri I, Müller C, Bassetto M, Manhardt T, Baumgartner-Parzer S, Kallay E, Schepelmann M. Nutritional and Pharmacological Targeting of the Calcium-Sensing Receptor Influences Chemically Induced Colitis in Mice. Nutrients. 2019; 11(12):3072. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123072
Chicago/Turabian StyleElajnaf, Taha, Luca Iamartino, Ildiko Mesteri, Christian Müller, Marcella Bassetto, Teresa Manhardt, Sabina Baumgartner-Parzer, Enikö Kallay, and Martin Schepelmann. 2019. "Nutritional and Pharmacological Targeting of the Calcium-Sensing Receptor Influences Chemically Induced Colitis in Mice" Nutrients 11, no. 12: 3072. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123072