Production of Bioactive Compounds by Food Associated Galactomyces geotrichum 38, as Determined by Proteome Analysis

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Microorganisms and Cultures
2.3. Identification of Proteins Produced by G. geotrichum 38
2.4. Determination of Sphingolipid Content
2.5. Determination of B Vitamin Content
2.6. Determination of Sterol Content of G. geotrichum 38 Biomass
2.7. Determination of Lipoic Acid Content
2.8. Determination of Trehalose Content of Culture
2.9. Production of Fried Cottage Cheese Using G. geotrichum 38 and Determination of Vitamin B2 and Ergosterol
3. Results and Discussion
3.1. Proteomic Analysis of Proteins Synthesized by G. geotrichum 38
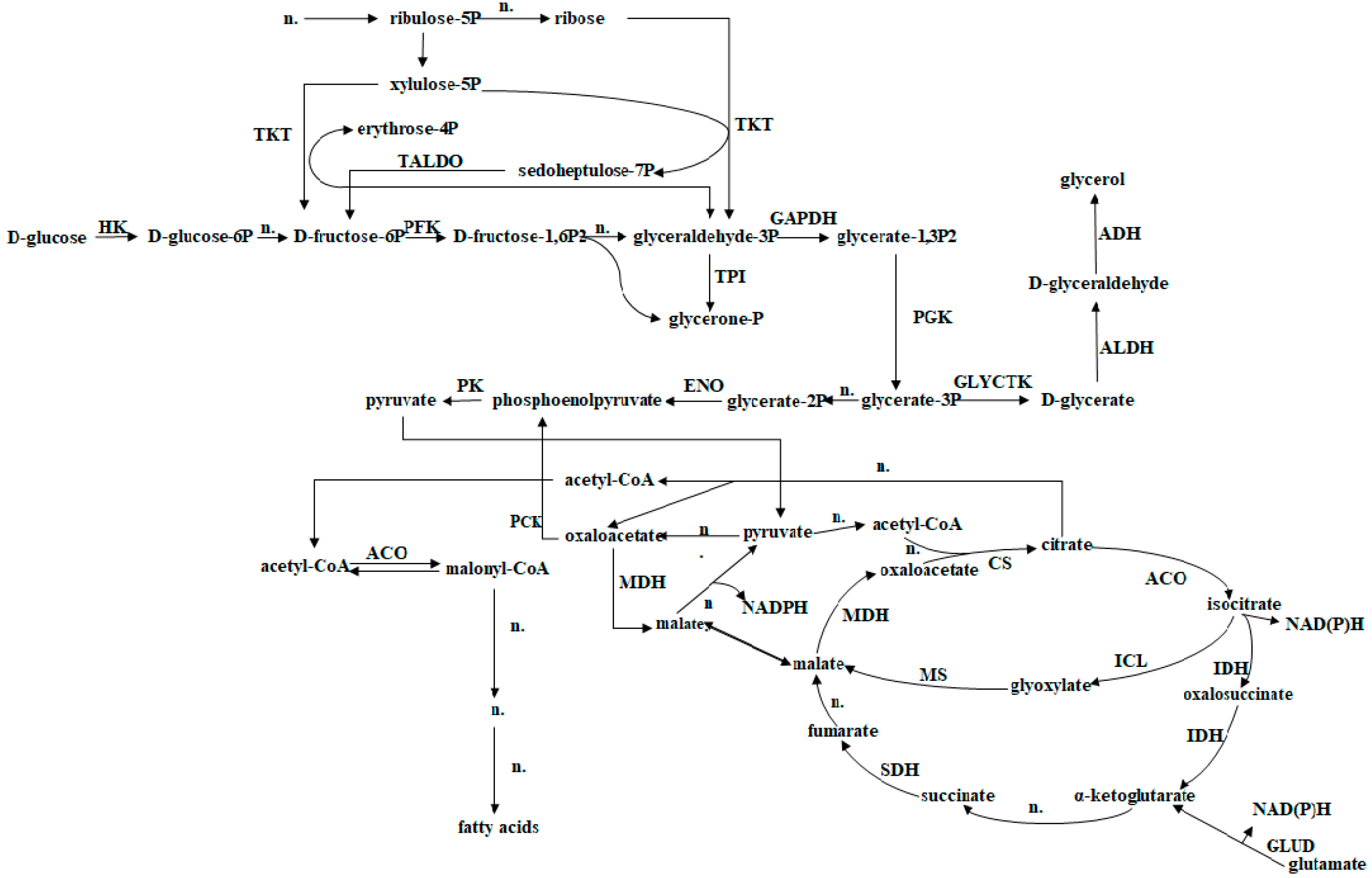
3.2. Hypothetical Fatty Acid Biosynthesis Pathway
3.3. Sphingolipids in G. geotrichum 38 Culture
3.4. Determination of B-Group Vitamins in G. geotrichum 38 Culture
3.5. Determination of Sterols in G. geotrichum 38 Biomass
3.6. Determination of Lipoic Acid Content of G. geotrichum 38 Culture
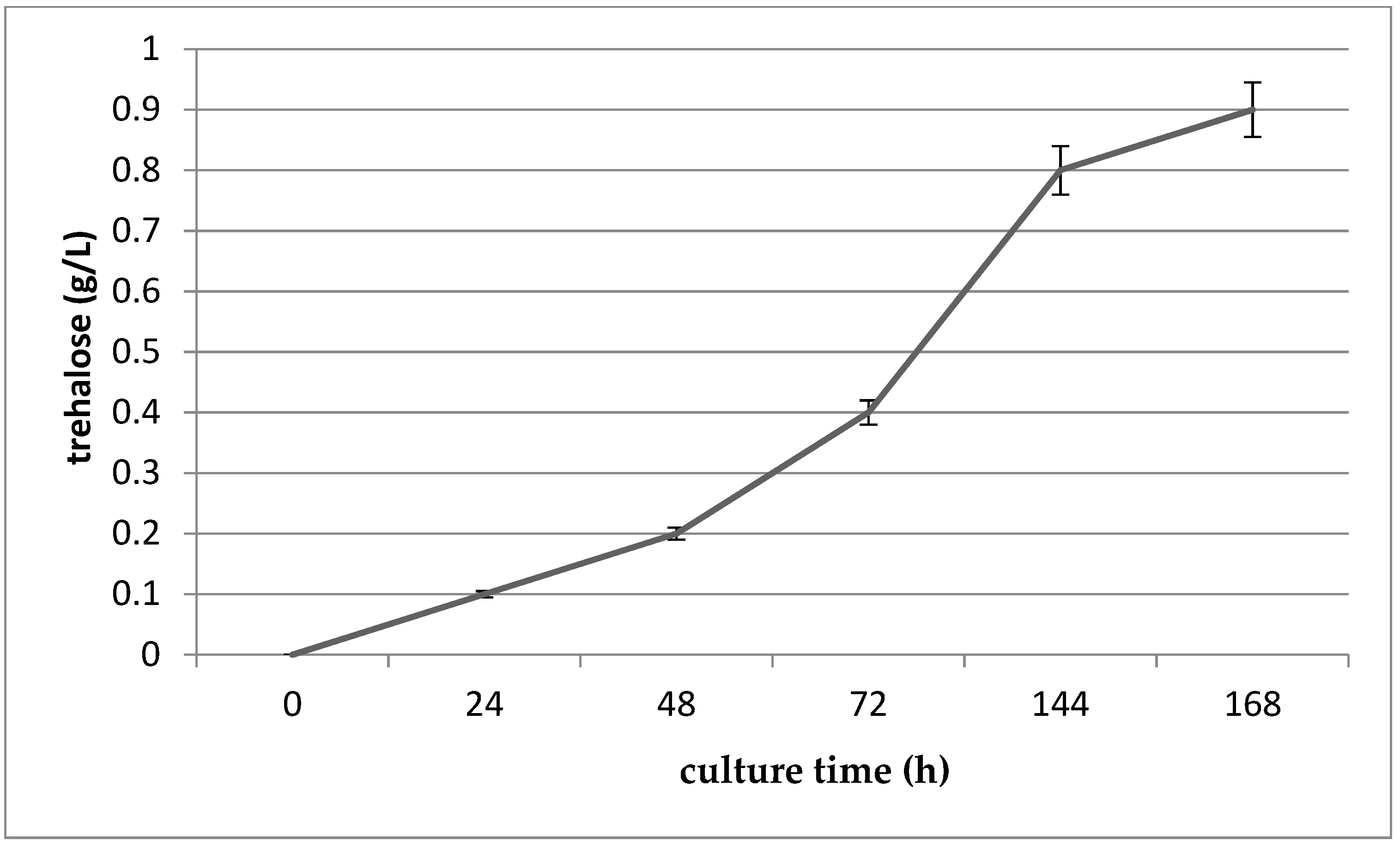
3.7. Determination of the Trehalose Content of G. geotrichum 38 Culture
3.8. Vitamin B2 and Ergosterol in Fried Cottage Cheese Produced by G. geotrichum 38
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shukla, R. Studies on bioactive compounds from different microorganisms. Int. J. Sci. Eng. Res. 2015, 6, 1225–1233. [Google Scholar]
- Chaves-Lopez, C.; Serio, A.; Rossi, C.; Pepe, A.; Compagnone, E.; Paparella, A. Interaction between Galactomyces geotrichum KL20B, Lactobacillus plantarum LAT3 and Enterococcus faecalis KE06 during milk fermentation. Fermentation 2017, 3, 52. [Google Scholar] [CrossRef]
- Grygier, A.; Myszka, K.; Rudzińska, M. Galactomyces geotrichum: Moulds from dairy products with high biotechnological potential. Acta Sci. Pol. Tecnol. Aliment. 2017, 16, 5–16. [Google Scholar] [CrossRef]
- Grygier, A.; Myszka, K.; Juzwa, W.; Białas, W.; Rudzińska, M. Ability of Galactomyces geotrichum for production of fatty acids. Folia Microbiol. under review.
- Szczepańska, E.; Robak, M. Proteomika w badaniu drobnoustrojów. Acta Sci. Pol. Biotech. 2013, 12, 25–40. [Google Scholar]
- Ledesma-Amaro, R.; Santos, M.A.; Jiménez, A.; Revuelta, J.L. Microbial production of vitamins. In Microbial Production of Food Ingredients, Enzymes and Nutraceuticals, 1st ed.; McNeil, B., Archer, D., Eds.; Woodhead Publishing: Cambridge, UK, 2013; pp. 571–594. [Google Scholar]
- Fogle, K.J.; Parson, K.G.; Dahm, N.A.; Holmes, T.C. Cryptochrome is a blue-light sensor that regulates neuronal firing rate. Science 2011, 6023, 1409–1413. [Google Scholar] [CrossRef] [PubMed]
- Nahlik, J.; Hrncirik, P.; Mares, J. Towards the design of an optimal strategy for the production of ergosterol from Saccharomyces cerevisiae yeasts. Biotechnol. Prog. 2017, 33, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Sułkowska-Ziaja, K.; Hałaszuk, P.; Mastej, M.; Piechaczek, M.; Muszyńska, B. Mycosteroles: Characteristics and biological importance. Med. Internacia Revuo 2016, 106, 26–34. [Google Scholar]
- Ostlund, R.E.; Racette, S.B.; Stenson, W.F. Effects of trace components of dietary fat on cholesterol metabolism: Phytosterols, oxysterols, and squalene. Nutr. Rev. 2002, 60, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Chalfant, C.; Del Poeta, M. Sphingolipids as Signaling and Regulatory Molecules, 1st ed.; Springer Science+Business Media: New York, NY, USA, 2010. [Google Scholar]
- Singh, A.; Del Poeta, M. Sphingolipidomics: An important mechanistic tool for studying fungal pathogens. Front. Microbiol. 2016, 7, 501. [Google Scholar] [CrossRef] [PubMed]
- Vesper, H.; Schmelz, E.M.; Nikolova-Karakashian, M.N.; Dillehay, D.L.; Lynch, D.V.; Merrill, A.H. Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J. Nutr. 1999, 129, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.; Deangelis, B.; Baker, E.R.; Hutner, S.H. A practical assay of lipoate in biologic fluids and liver in health and disease. Free Radic. Biol. Med. 1998, 25, 473–479. [Google Scholar] [CrossRef]
- Chen, Q.; Haddad, G.G. Role of trehalose phosphate synthase and trehalose during hypoxia: From flies to mammals. J. Exp. Biol. 2004, 207, 3125–3129. [Google Scholar] [CrossRef] [PubMed]
- Deesuth, O.; Laopaiboon, P.; Laopaiboon, L. High ethanol production under optimal aeration conditions and yeast composition in a very high gravity fermentation from sweet sorghum juice by Saccharomyces cerevisiae. Ind. Crop Prod. 2016, 92, 263–270. [Google Scholar] [CrossRef]
- Bald, E.; Sypniewski, S.; Drzewoski, J.; Stępień, M. Application of 2-halopyridinium salts as ultraviolet derivatization reagents and solid-phase extraction for determination of captopril in human plasma by high-performance liquid chromatography. J. Chromatogr. B 1996, 681, 283–289. [Google Scholar] [CrossRef]
- Bajpai, P.; Bajpai, P.K.; Ward, O.P. Eicosapentaenoic acid (EPA) formation: Comparative studies with Mortierella strains and production by Mortierella elongate. Mycol. Res. 1996, 95, 1294–1298. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Celińska, E.; Olkowicz, M.; Grajek, W. L-Phenylalanine catabolism and 2-phenylethanol synthesis in Yarrowia lipolytica—Mapping molecular identities through whole-proteome quantitative mass spectrometry analysis. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef]
- Zhang, W.; Quinn, B.; Barnes, S.; Grabowski, G.; Sun, Y.; Setchell, K. Metabolic profiling and quantification of sphingolipids by liquid chromatography-tandem mass spectrometry. J. Glycomics Lipidomics 2013, 3, 170. [Google Scholar] [CrossRef]
- Zand, N.; Chowdhry, B.Z.; Pullen, F.S.; Snowden, M.J.; Tetteh, J. Simultaneous determination of riboflavin and pyridoxine by UHPLC/LC–MS in UK commercial infant meal food products. Food Chem. 2012, 135, 2743–2749. [Google Scholar] [CrossRef] [PubMed]
- Perkowski, J.; Wiwart, M.; Buśko, M.; Laskowska, M.; Berthiller, F.; Kandler, W.; Krska, R. Fusarium toxins and total fungal biomass indicators in naturally contaminated wheat samples from north-eastern Poland in 2003. Food Addit. Contam. 2007, 24, 1292–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chwatko, G.; Krawczyk, M.; Iciek, M.; Kamińska, A.; Bilska-Wilkosz, A.; Marcykiewicz, B.; Głowacki, R. Determination of lipoic acid in human plasma by high-performance liquid chromatography with ultraviolet detection. Arab. J. Chem. 2016. [Google Scholar] [CrossRef] [Green Version]
- Pawlicka, J.; Drożdżyńska, A.; Kośmider, A.; Czaczyk, K. Wpływ buforu fosforanowego na produkcję biomasy, kwasu propionowego i trehalozy przez Propionibacterium freudenreichii ssp. shermanii 1. EPISTEME. Czasopismo Naukowo Kulturalne 2015, 26, 85–93. [Google Scholar]
- Schmidt, A.; Schreiner, M.G.; Mayer, H.K. Rapid determination of the various native forms of vitamin B6 and B2 in cow’s milk using ultra-high performance liquid chromatography. J. Chromatog. A 2017, 1500, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Stancher, B.; Zonta, F. High performance liquid chromatographic analysis of riboflavin (vitamin B2) with visible absorbance detection in Italian cheeses. J. Food Sci. 1986, 51, 857–858. [Google Scholar] [CrossRef]
- Tong, L. Acetyl-coenzyme A carboxylase: Crucial metabolic enzyme and attractive target for drug discovery. Cell. Mol. Life Sci. 2005, 62, 1784–1803. [Google Scholar] [CrossRef] [PubMed]
- Besada-Lombana, P.; Fernandez-Moya, R.; Fenster, J.; Da Silva, N.A. Engineering Saccharomyces cerevisiae fatty acid composition for increased tolerance to octanoic acid. Biotechnol. Bioeng. 2017, 114, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Yang, J.; Cao, Y.; Li, L.; Jiang, X.; Xu, X.; Liu, W.; Xian, M.; Zhang, Y. Increasing fatty acid production in E. coli by simulating the lipid accumulation of oleaginous microorganisms. J. Ind. Microbiol. Biot. 2011, 38, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Simopoulus, A.P. Omega-3 fatty acids in inflammation and autoimmution disease. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- Okamoto, T.; Sekiyama, N.; Otsu, M.; Shimada, Y.; Sato, A.; Nakanishi, S.; Jingami, H. Expression and purification of the extracellular ligand binding region of metabotropic glutamate receptor subtype 1. J. Biol. Chem. 1998, 273, 13089–13096. [Google Scholar] [CrossRef] [PubMed]
- Sofia, H.J.; Chen, G.; Hetzler, B.G.; Reyes-Spindola, J.F.; Miller, N.E. Radical SAM, a novel protein superfamily linking unresolved steps in familiar biosynthetic pathways with radical mechanisms: Functional characterization using new analysis and information visualization methods. Nucleic Acid Res. 2001, 29, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Sandoval, G.; Ambrogelly, A.; Rinehart, J.; Wei, D.; Cruz-Vera, L.R.; Graham, D.E.; Stetter, K.O.; Guarneros, G.; Söll, D. Orthologs of a novel archaeal and of the bacterial peptidyl–tRNA hydrolase are nonessential in yeast. Proc. Natl. Acad. Sci. USA 2002, 99, 16707–16712. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/uniprot/A0A2T5P0V1 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/P09980 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/Q9FVX1 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/Q92II9 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/U5QN56 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/A0A2T3QGM2 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/A0A0F2KUL7 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/A7NPP8 (accessed on 10 February 2019).
- Sreekumar, P.G.; Hinton, D.R.; Kannan, R. Methionine sulfoxide reductase A: Structure, function and role in ocular pathology. World J. Biol. Chem. 2011, 2, 184–192. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/uniprot/Q8EQ75 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/B7L440 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/C4ZRR2 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/W0M2E9 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/P0ABM9 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/E8W756 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/B9MG17 (accessed on 10 February 2019).
- Du, C.; Wenzel, G.P. Mining for microbial gems: Integrating proteomics in the postgenomic natural product discovery pipeline. Proteomics 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Wallis, J.G.; Browse, J. The Δ8-desaturase of Euglena gracilis: An alternate pathway for synthesis of 20-carbon polyunsaturated fatty acids. Arch. Biochem. Biophys. 1999, 365, 307–316. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/uniprot/A0A0Y9XSK8 (accessed on 10 February 2019).
- Kumar, V.P.; West, A.H.; Cook, P.F. Supporting role of lysine 13 and glutamate 16 in the acid-base mechanism of saccharopine dehydrogenase from Saccharomyces cerevisiae. Arch. Biochem. Biophys. 2012, 522, 57–61. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/uniprot/P32626 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/P33734 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/P40207 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/Q92410 (accessed on 10 February 2019).
- UniProt. Available online: https://www.uniprot.org/uniprot/P32875 (accessed on 10 February 2019).
- Miyazaki, K. Bifunctional isocitrate–homoisocitrate dehydrogenase: A missing link in the evolution of β-decarboxylating dehydrogenase. Biochem. Bioph. Res. Co. 2005, 331, 341–346. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/uniprot/P0CM18 (accessed on 10 February 2019).
- Hego, E.; Bes, C.M.; Bedon, F.; Palagi, P.M.; Chaumeil, P.; Barré, A.; Claverol, S.; Dupuy, J.W.; Bonneu, M.; Lalanne, C.; et al. Differential accumulation of soluble proteins in roots of metallicolous and nonmetallicolous populations of Agrostis capillaris L. exposed to Cu. Proteomics 2014, 14, 1746–1758. [Google Scholar] [CrossRef] [PubMed]
- Hames, B.D.; Hooper, N.M. Krótkie wykłady. Biochemia, 3rd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2004. [Google Scholar]
- Wang, L.; Chen, W.; Feng, Y.; Ren, Y.; Gu, Z.; Chen, H.; Wang, H.; Thomas, M.J.; Zhang, B.; Berquin, I.M.; et al. Genome characterization of the oleaginous fungus Mortierella alpina. PLoS ONE 2011, 6, e28319. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.A.; Mokhtar, N.F.; Taha, E.M.; Omar, O.; Yusoff, W.M.W. The role of ATP citrate lyase, malic enzyme and fatty acid synthase in the regulation of lipid accumulation in Cunninghamella sp. 2A1. Ann. Microbiol. 2011, 61, 463–468. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Techol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Alosta, H.A. Riboflavin Production by Encapsulated Candida flareri. PhD. Thesis, University of Oklahoma, Norman, OK, USA, 2007. [Google Scholar]
- Stahmann, K.P.; Revuelta, J.L.; Seulberger, H. Three biotechnical processes using Ashbya gossypii, Candida famata, or Bacillus subtilis compete with chemical riboflavin production. Appl. Microbiol. Biot. 2000, 53, 509–516. [Google Scholar] [CrossRef]
- Contreras, G.; Barahona, S.; Sepúlveda, D.; Baeza, M.; Cifuentes, V.; Alcaíno, J. Identification and analysis of metabolite production with biotechnological potential in Xanthophyllomyces dendrorhous isolates. World J. Microb. Biot. 2015, 31, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, E.J. Production of vitamins, coenzymes and related biochemicals by biotechnological processes. J. Chem. Technol. Biot. 1992, 53, 313–327. [Google Scholar] [CrossRef]
- Herbert, A.A.; Guest, J.R. Lipoic acid content of Escherichia coli and other microorganisms. Arch. Microbiol. 1975, 106, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Borowczyk, K.; Krawczyk, M.; Kubalczyk, P.; Chwatko, G. Determination of lipoic acid in biological samples. Bioanalysis 2015, 14, 1785–1798. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.S.; Gaspar, P.; Hugenholtz, J.; Ramos, A.; Santos, H. Enhancement of trehalose production in dairy Propionibacteria through manipulation of environmental conditions. Int. J. Food Microbiol. 2004, 91, 195–204. [Google Scholar] [CrossRef]
- Pawlicka-Karczorowska, J.; Czaczyk, K. Effect of crude and pure glycerol on biomass production and trehalose accumulation by Propionibacterium freudenreichii ssp. shermanii 1. Acta Biochim. Pol. 2017, 64, 621–629. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Proteins Extracted from Hyphae | Possible Function |
| extracellular ligand-binding receptor | extracellular signal molecule [32] |
| radical sterile alpha motif domain protein | reducing S-adenosyl L-methionine [33] |
| peptidyl-tRNA hydrolase | cell cycle [34] |
| flavin adenine dinucleotide dependent oxidoreductase | catalyzing the oxidation of amino acids [35] |
| ATP-dependent DNA helicase Rep | cell cycle [36] |
| glutaredoxin C3 | electron carrier in the synthesis of deoxyribonucleotides [37] |
| protein kinase C inhibitor 1 | inhibiting protein kinase C activity [38] |
| excinuclease ATP-binding cassette subunit C | repairing DNA [39] |
| nicotinate (nicotinamide) nucleotide adenylyltransferase | metabolism of nicotine and nicotinamide [40] |
| acetyl co-enzyme A carboxylase carboxyltransferase alpha subunit | biosynthesis of fatty acids [28] |
| Silent Information Regulator A family protein | inducing arthrospore creation [41] |
| pyruvate carboxyltransferase | carboxylation of pyruvate [42] |
| methionine sulfoxide reductase A | reducing methionine sulfoxide to methionine [43] |
| transposase for IS660 | cell cycle [44] |
| transposase, IS4 family | cell cycle [45] |
| Proteins Extracted from Culture | Possible Function |
| elongation factor thermo stable | cell cycle [46] |
| putative endolysin | catabolism of chitin [47] |
| cytochrome C biogenesis protein, cytochromes c maturation protein H family protein | hem lyase subunit [48] |
| helix-turn-helix, AraC domain protein | cell cycle [49] |
| heavy metal transport/detoxification protein | metal transport [50] |
| Proteins with an Unidentified Function | |
| hypothetical protein HMPREF7215_1957 | |
| hypothetical protein bthur0004_61160 | |
| hypothetical protein Veis_3482 | |
| PREDICTED: similar to hCG1786587 | |
| hypothetical protein XF2003 | |
| hypothetical protein RPC_3615 | |
| putative endolysin of prophage CP-933O | |
| Proteins Extracted from Hyphae | Possible Protein Function |
| delta(8)-fatty-acid desaturase | sphingolipid biosynthesis [52] |
| pyridoxine biosynthesis PDX1-like protein | biosynthesis of B-group vitamins [53] |
| saccharopine dehydrogenase | lysine biosynthesis [54] |
| enolase-phosphatase E1 | methionine biosynthesis [55] |
| imidazole glycerol phosphate synthase hisHF | histidine biosynthesis [56] |
| Proteins Extracted from Culture Fluid | Possible Protein Function |
| pyridoxine biosynthesis PDX1-like protein | biosynthesis of B-group vitamins [53] |
| uncharacterized protein YMR134W | ergosterol biosynthesis [57] |
| alpha,alpha-trehalose-phosphate synthase | trehalose biosynthesis [58] |
| lipoyl synthase, mitochondrial | lipoic acid biosynthesis [59] |
| 2-isopropylmalate synthase;3-isopropylmalate dehydrogenase | leucine biosynthesis [60] |
| amino-acid acetyltransferase, mitochondrial | arginine biosynthesis [61] |
| probable 5-methyltetrahydropteroyltriglutamatehomocysteine methyltransferase | methionine biosynthesis [62] |
| Sphingolipid {%} | |||||||
|---|---|---|---|---|---|---|---|
| Culture Time (h) | (1.) | (2.) | (3.) | (4.) | (5.) | (6.) | (7.) |
| 48 | 25.7 a ± 1.0 | 0.2 a ± 0.0 | 1.1 a ± 0.0 | 0.5 b ± 0.0 | 8.7 b ± 0.1 | 3.0 b ± 0.0 | 60.8 a ± 1.7 |
| 216 | 28.2 b ± 0.3 | 0.9 b ± 0.0 | 0.6 a ± 0.1 | 0.2 a ± 0.0 | 5.9 a ± 0.1 | 0.3 a ± 0.0 | 63.9 b ± 0.5 |
| Culture Time {h} | Vitamin B2 Content µg/L |
|---|---|
| 0 | 92 a ± 4 |
| 216 | 224 b ± 10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grygier, A.; Myszka, K.; Szwengiel, A.; Stuper-Szablewska, K.; Pawlicka-Kaczorowska, J.; Chwatko, G.; Rudzińska, M. Production of Bioactive Compounds by Food Associated Galactomyces geotrichum 38, as Determined by Proteome Analysis. Nutrients 2019, 11, 471. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020471
Grygier A, Myszka K, Szwengiel A, Stuper-Szablewska K, Pawlicka-Kaczorowska J, Chwatko G, Rudzińska M. Production of Bioactive Compounds by Food Associated Galactomyces geotrichum 38, as Determined by Proteome Analysis. Nutrients. 2019; 11(2):471. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020471
Chicago/Turabian StyleGrygier, Anna, Kamila Myszka, Artur Szwengiel, Kinga Stuper-Szablewska, Joanna Pawlicka-Kaczorowska, Grażyna Chwatko, and Magdalena Rudzińska. 2019. "Production of Bioactive Compounds by Food Associated Galactomyces geotrichum 38, as Determined by Proteome Analysis" Nutrients 11, no. 2: 471. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020471