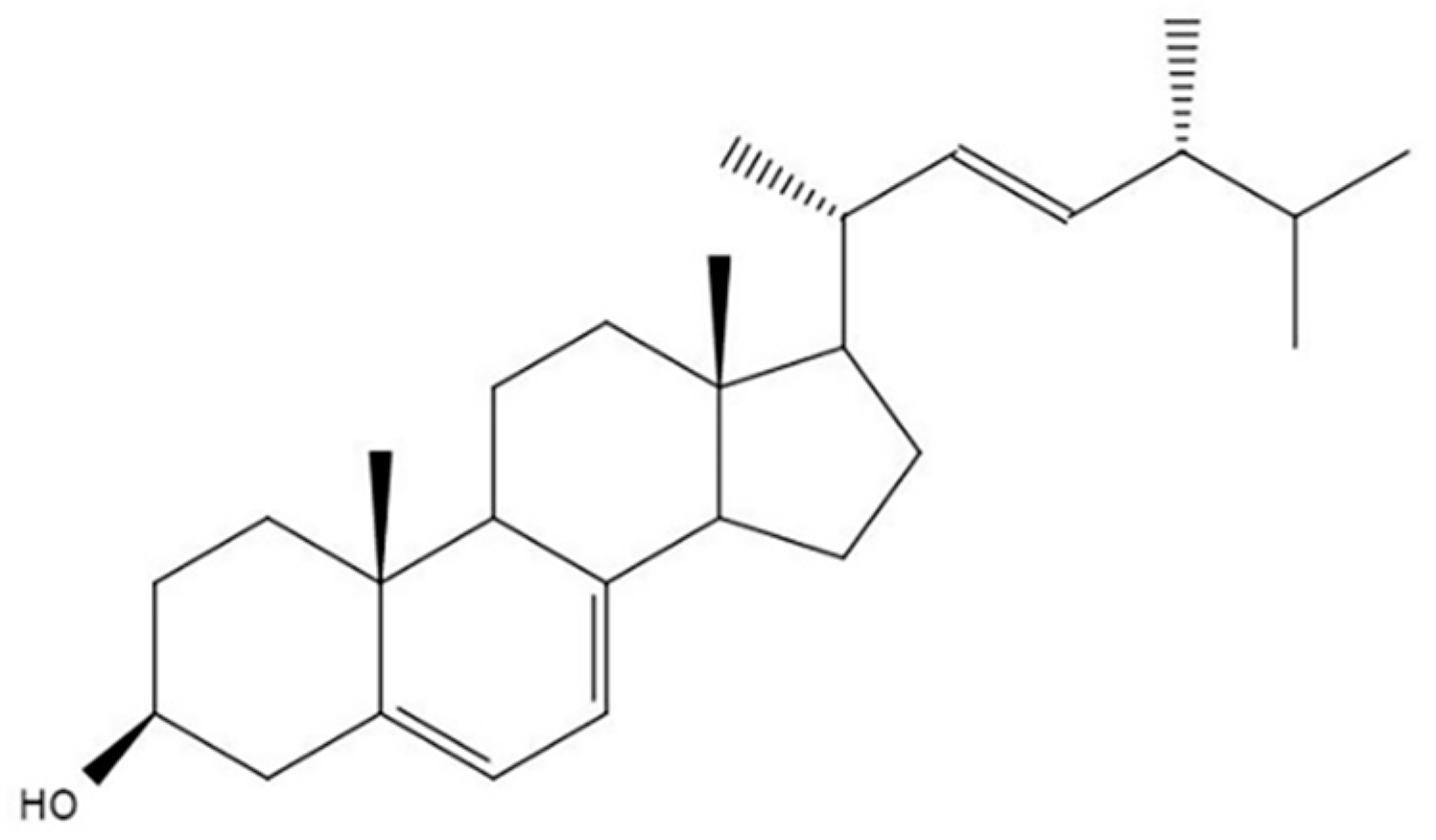
Ergosterol Ameliorates Diabetic Nephropathy by Attenuating Mesangial Cell Proliferation and Extracellular Matrix Deposition via the TGF-β1/Smad2 Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture and Treatment
2.3. Cell Proliferation Analysis
2.4. Animal Models
2.5. Histological and Morphological Examination
2.6. Immunohistochemical Analysis
2.7. Western Blot Analysis
2.8. Quantitative Real-Time RT-PCR
2.9. Statistical Analysis
3. Results
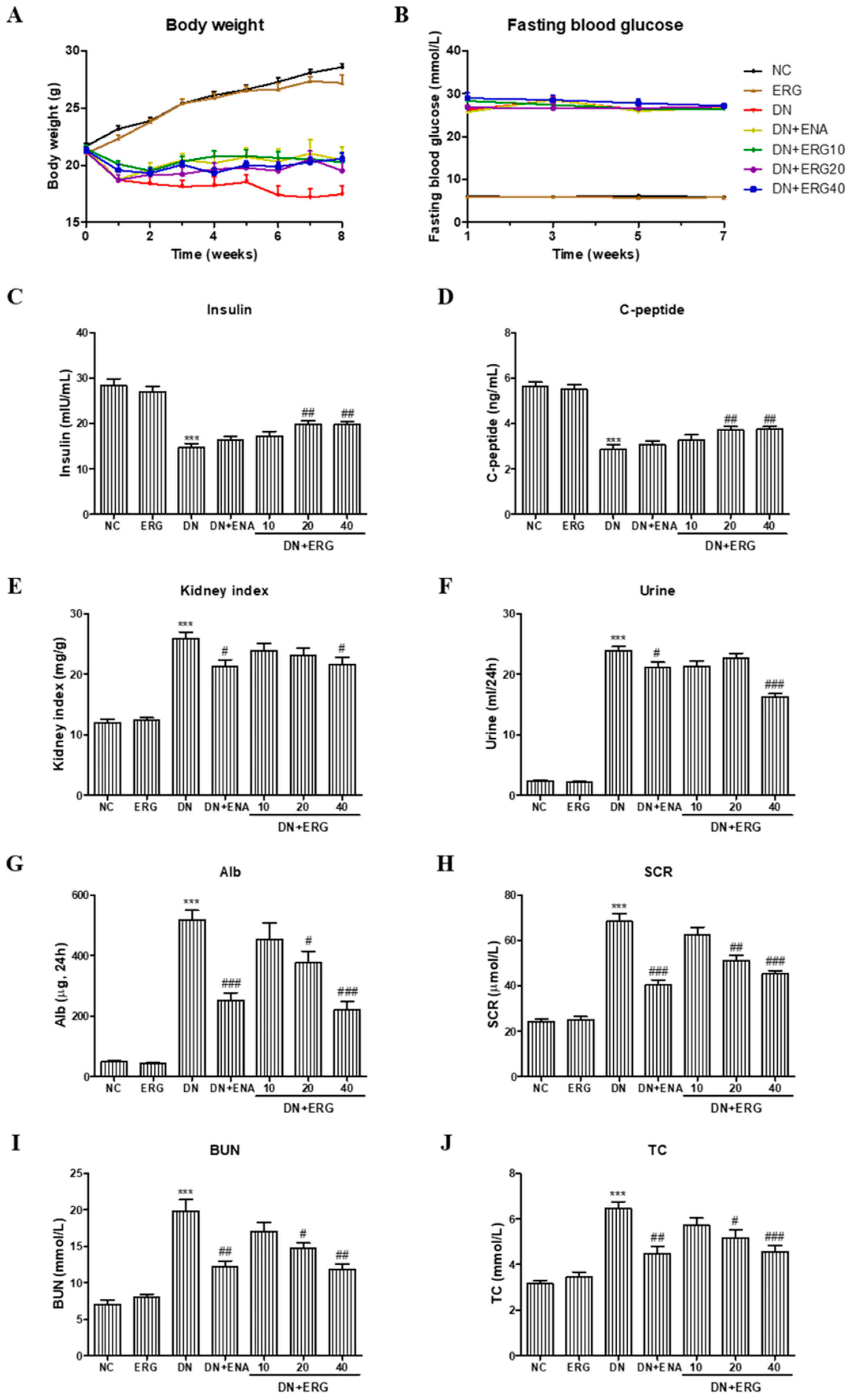
3.1. Effects of Ergosterol on Metabolic and Biochemical Parameters
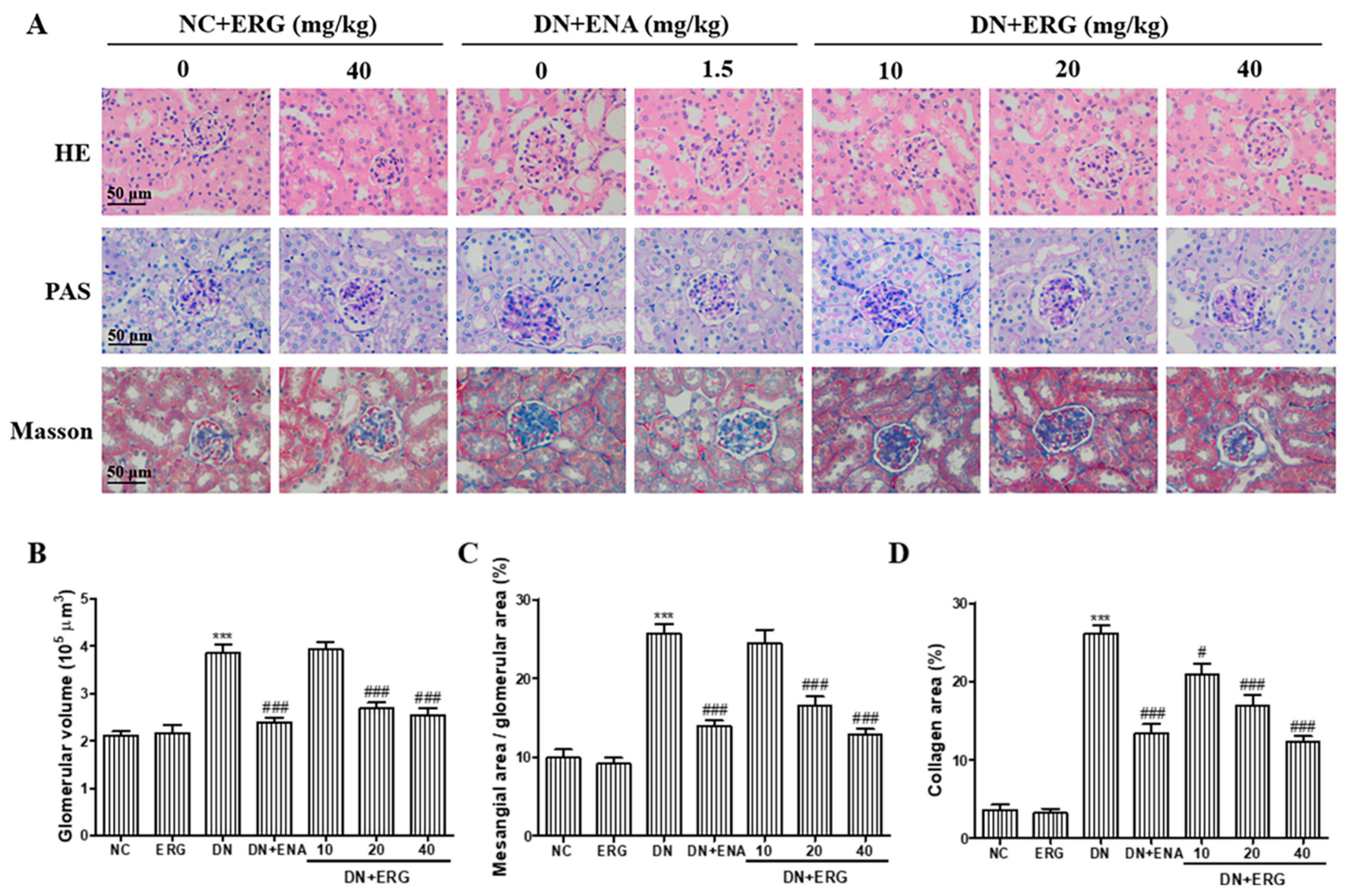
3.2. Effects of Ergosterol on Histopathological Changes in Renal Tissue
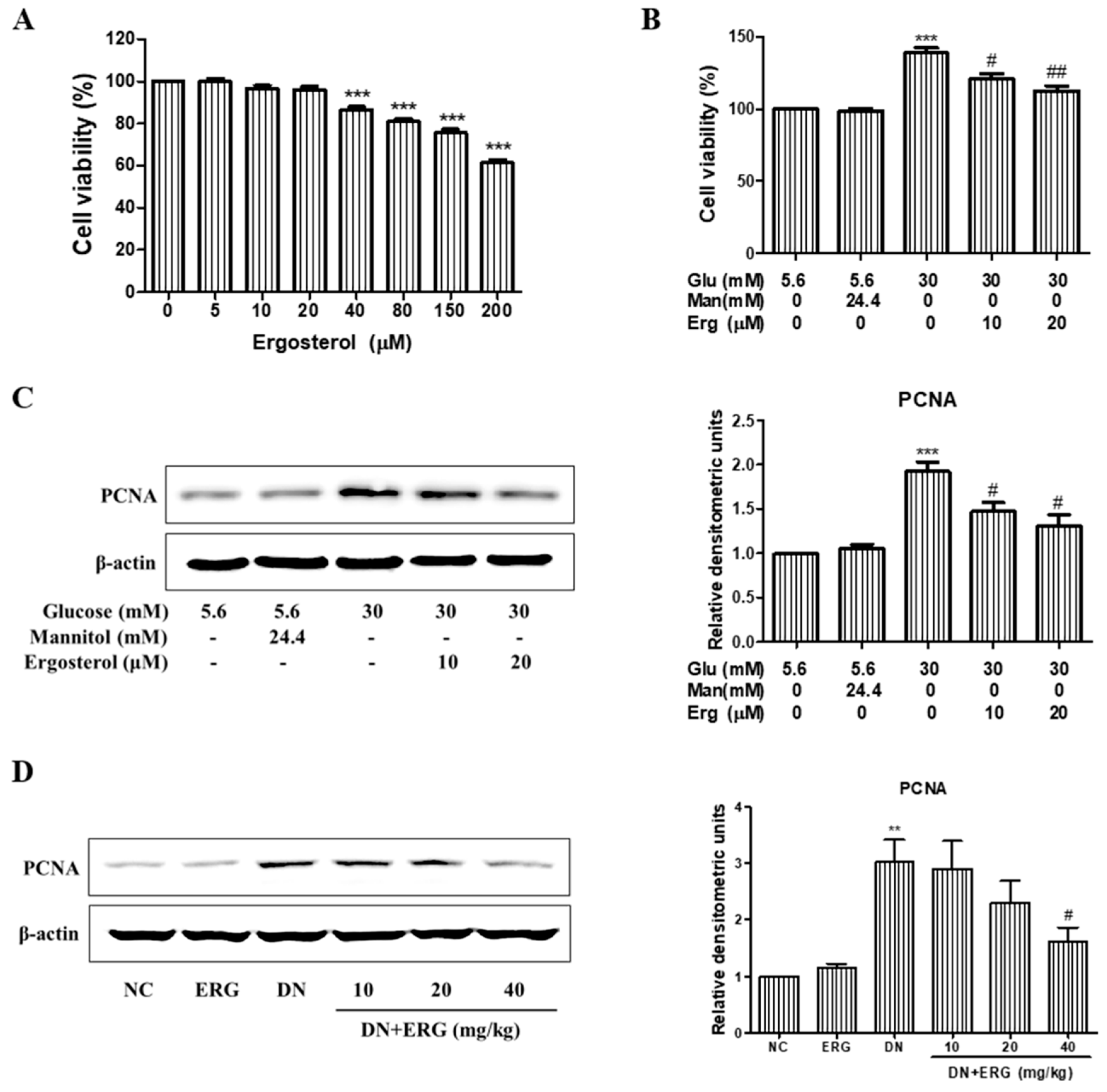
3.3. Ergosterol Significantly Inhibited the Proliferation of Renal Mesangial Cells
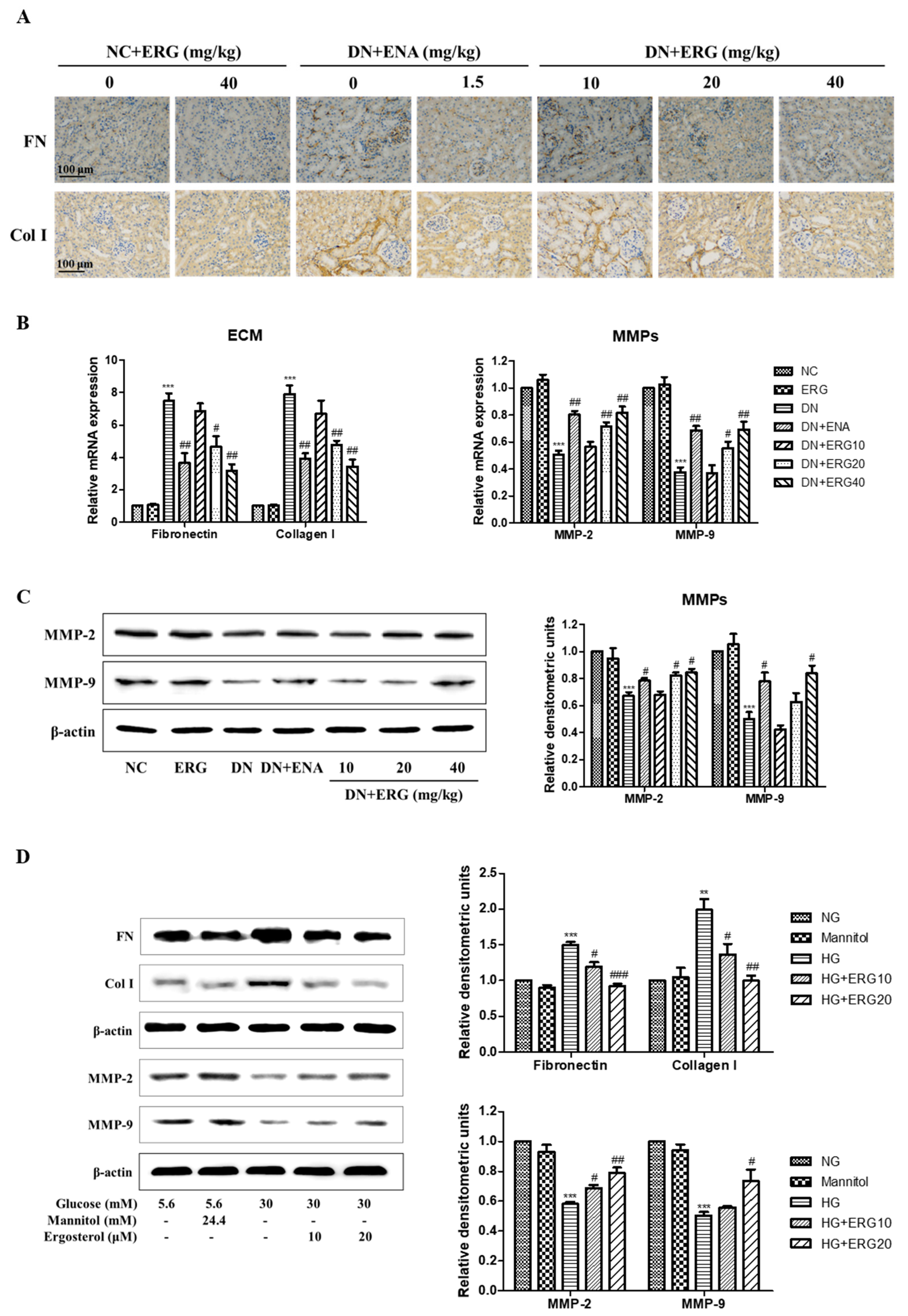
3.4. Ergosterol Significantly Alleviated ECM Accumulation
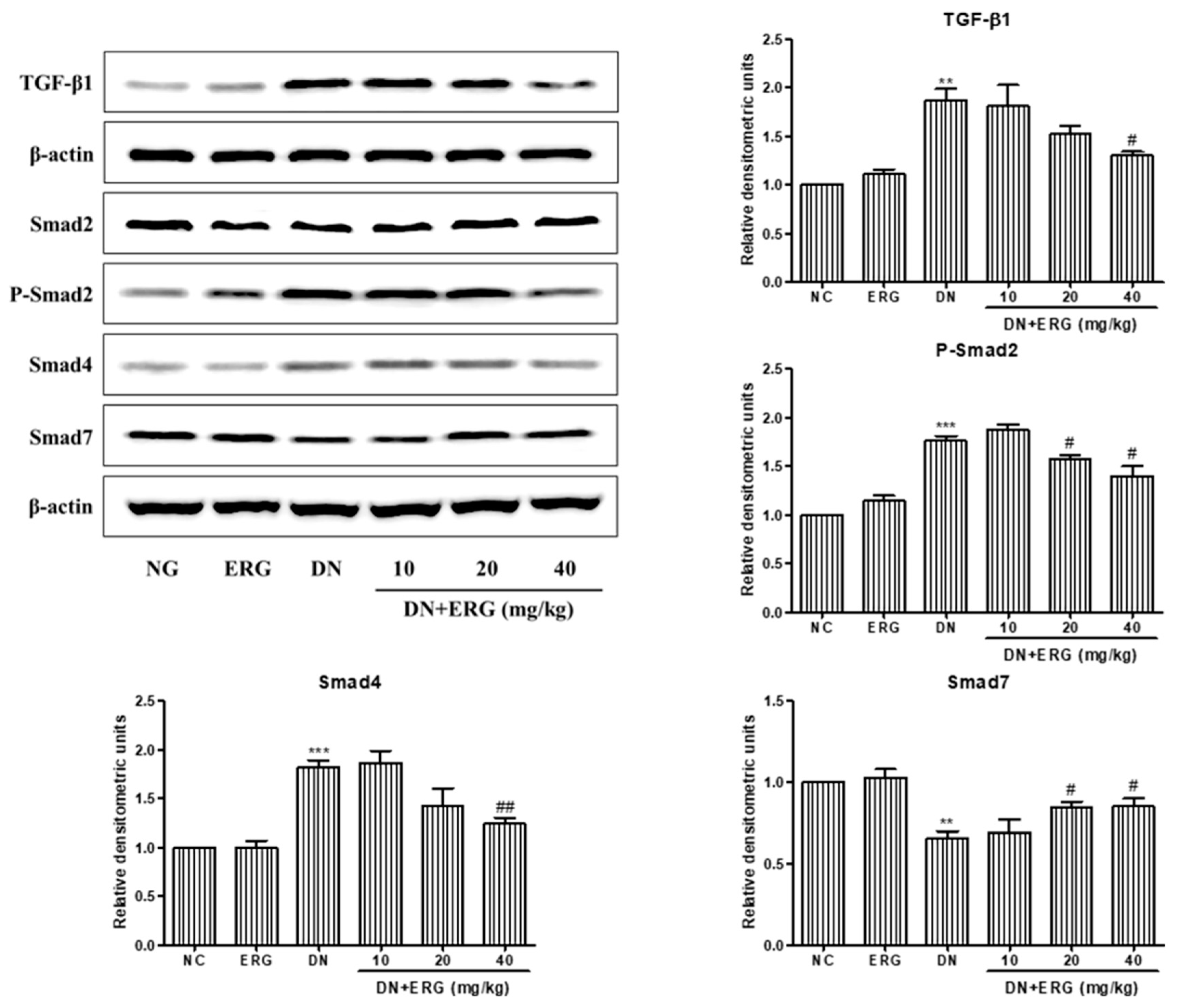
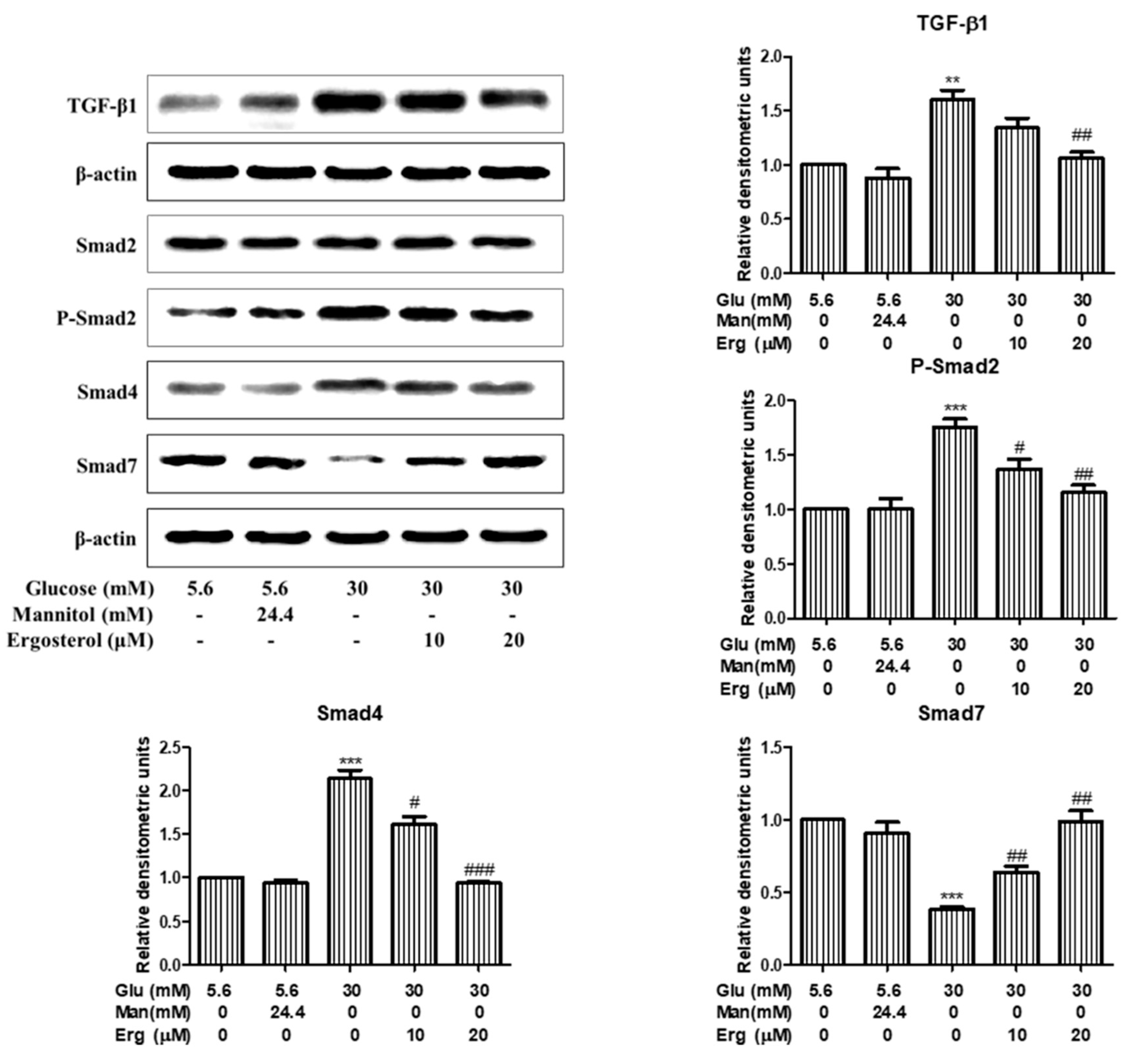
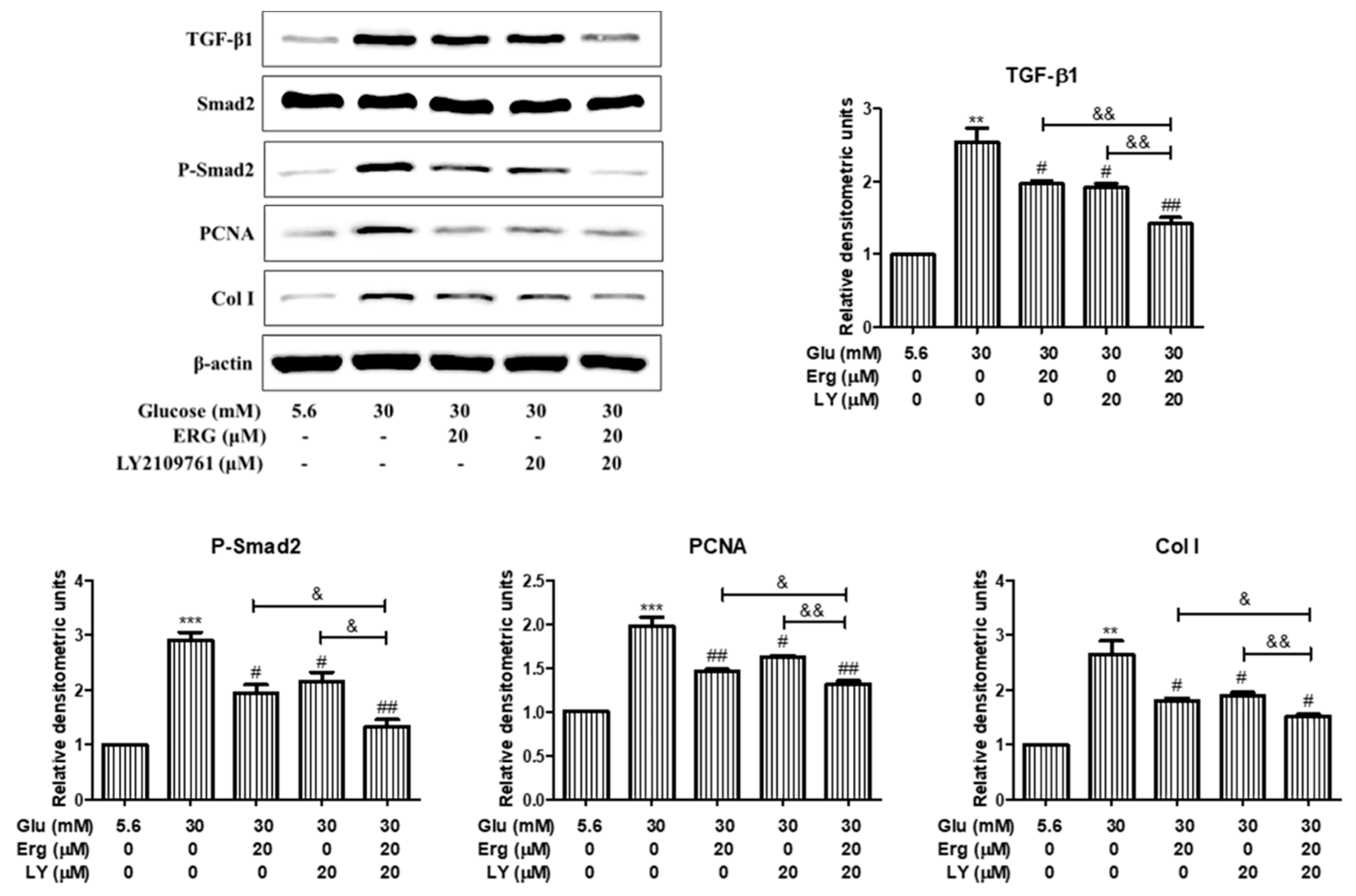
3.5. Ergosterol Inhibited Renal Mesangial Cells Proliferation and ECM Accumulation through TGF-B1/Smad2 Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hu, J.; Ye, M.; Zhou, Z. Aptamers: Novel diagnostic and therapeutic tools for diabetes mellitus and metabolic diseases. J. Mol. Med. 2017, 95, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Qiang, G.; Zhao, Y.; Yang, X.; Chen, X.; Yan, Y.; Wang, X.; Liu, C.; Zhang, L.; Du, G. Salvianolic acid a protects against diabetic nephropathy through ameliorating glomerular endothelial dysfunction via inhibiting age-rage signaling. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 2378–2394. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.J.; Ding, H.H.; Zhou, H.; Qiu, Y.Y.; Tang, L.Q. Renoprotective effects of berberine through regulation of the mmps/timps system in streptozocin-induced diabetic nephropathy in rats. Eur. J. Pharmacol. 2015, 764, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Kong, L.; Tan, Y.; Epstein, P.N.; Zeng, J.; Gu, J.; Liang, G.; Kong, M.; Chen, X.; Miao, L.; et al. C66 ameliorates diabetic nephropathy in mice by both upregulating nrf2 function via increase in mir-200a and inhibiting mir-21. Diabetologia 2016, 59, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Abboud, H.E. Mesangial cell biology. Exp. Cell Res. 2012, 318, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, M.; Chen, B.; Shi, J. Scoparone attenuates high glucose-induced extracellular matrix accumulation in rat mesangial cells. Eur. J. Pharmacol. 2017, 815, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Narula, S.; Tandon, C.; Tandon, S. Role of matrix metalloproteinases in degenerative kidney disorders. Curr. Med. Chem. 2018, 25, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Krochmal, M.; Kontostathi, G.; Magalhaes, P.; Makridakis, M.; Klein, J.; Husi, H.; Leierer, J.; Mayer, G.; Bascands, J.L.; Denis, C.; et al. Urinary peptidomics analysis reveals proteases involved in diabetic nephropathy. Sci. Rep. 2017, 7, 15160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.Y.; Huang, P.H.; Yang, A.H.; Tarng, D.C.; Yang, W.C.; Lin, C.C.; Chen, J.W.; Schmid-Schonbein, G.; Lin, S.J. Matrix metalloproteinase-9 deficiency attenuates diabetic nephropathy by modulation of podocyte functions and dedifferentiation. Kidney Int. 2014, 86, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Sawires, H.; Botrous, O.; Aboulmagd, A.; Madani, N.; Abdelhaleem, O. Transforming growth factor-beta1 in children with diabetic nephropathy. Pediatr. Nephrol. 2019, 34, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, N.; Tahira, Y.; Matsuda, H.; Matsumoto, K. Transforming growth factor-beta as a treatment target in renal diseases. J. Nephrol. 2009, 22, 708–715. [Google Scholar] [PubMed]
- Lee, E.J.; Kang, M.K.; Kim, D.Y.; Kim, Y.H.; Oh, H.; Kang, Y.H. Chrysin inhibits advanced glycation end products-induced kidney fibrosis in renal mesangial cells and diabetic kidneys. Nutrients 2018, 10, 882. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Su, P.; Zhang, W. Advances in microalgae-derived phytosterols for functional food and pharmaceutical applications. Mar. Drugs 2015, 13, 4231–4254. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, E.A.; Koppenol, W.P.; de Jong, A.; Hiemstra, H.; Vermeer, M.A.; Noakes, M.; Luscombe-Marsh, N.D. Plant sterols lower ldl-cholesterol and triglycerides in dyslipidemic individuals with or at risk of developing type 2 diabetes; a randomized, double-blind, placebo-controlled study. Nutr. Diabetes 2018, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegard, L.; Jessup, W.; Jones, P.J.; Lutjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassbender, K.; Lutjohann, D.; Dik, M.G.; Bremmer, M.; Konig, J.; Walter, S.; Liu, Y.; Letiembre, M.; von Bergmann, K.; Jonker, C. Moderately elevated plant sterol levels are associated with reduced cardiovascular risk--the lasa study. Atherosclerosis 2008, 196, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Hashim, S.N.; Schwarz, L.J.; Danylec, B.; Mitri, K.; Yang, Y.; Boysen, R.I.; Hearn, M.T. Recovery of ergosterol from the medicinal mushroom, ganoderma tsugae var. Janniae, with a molecularly imprinted polymer derived from a cleavable monomer-template composite. J. Chromatogr. A 2016, 1468, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, C.; Wang, T.; Zhang, P.; Liu, Z.; Lu, C. Ergosterol attenuates lps-induced myocardial injury by modulating oxidative stress and apoptosis in rats. Cell. Physiol. Biochem. 2018, 48, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Huang, Y.; Liu, Y.; Huang, M.; Song, G.; Ming, Q.; Ma, X.; Yang, J.; Deng, S.; Wen, Y.; et al. Antidiabetic activity of ergosterol from pleurotus ostreatus in kk-a(y) mice with spontaneous type 2 diabetes mellitus. Mol. Nutr. Food Res. 2018, 62, 1700444. [Google Scholar] [CrossRef] [PubMed]
- Huan, W.; Tianzhu, Z.; Yu, L.; Shumin, W. Effects of ergosterol on copd in mice via jak3/stat3/nf-kappab pathway. Inflammation 2017, 40, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Vaneckova, I.; Kujal, P.; Huskova, Z.; Vanourkova, Z.; Vernerova, Z.; Certikova Chabova, V.; Skaroupkova, P.; Kramer, H.J.; Tesar, V.; Cervenka, L. Effects of combined endothelin a receptor and renin-angiotensin system blockade on the course of end-organ damage in 5/6 nephrectomized ren-2 hypertensive rats. Kidney Blood Press. Res. 2012, 35, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, J.M.; Wang, Y.; Wang, X.; Niu, A.; Gewin, L.S.; Yao, B.; Harris, R.C.; Zhang, M.-Z. Selective activation of epidermal growth factor receptor in renal proximal tubule induces tubulointerstitial fibrosis. FASEB J. 2017, 31, 4407–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Lu, C.; Dai, Q.; Sheng, J.; Xu, M. Sirt3 facilitates amniotic fluid stem cells to repair diabetic nephropathy through protecting mitochondrial homeostasis by modulation of mitophagy. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1508–1524. [Google Scholar] [CrossRef] [PubMed]
- Azushima, K.; Gurley, S.B.; Coffman, T.M. Modelling diabetic nephropathy in mice. Nat. Rev. Nephrol. 2018, 14, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.; Yuguang, L.; Liying, W.; Shuying, Z.; Liting, X.; Shumin, W. Ergosterol alleviates kidney injury in streptozotocin-induced diabetic mice. Evid.-Based Complement. Altern. Med. eCAM 2015, 2015, 691594. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.J.; Tang, L.Q.; Zhou, H.; Ding, H.H.; Qiu, Y.Y. Renoprotective effect of berberine via regulating the pge2-ep1-galphaq-ca(2+) signalling pathway in glomerular mesangial cells of diabetic rats. J. Cell. Mol. Med. 2016, 20, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Chen, W.; Li, J.; Ding, J.; Mei, X.; Hu, H. High glucose induces mouse mesangial cell overproliferation via inhibition of hydrogen sulfide synthesis in a tlr-4-dependent manner. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wen, X.; Han, X.R.; Wang, Y.J.; Shen, M.; Fan, S.H.; Zhuang, J.; Zhang, Z.F.; Shan, Q.; Li, M.Q.; et al. Repression of microrna-382 inhibits glomerular mesangial cell proliferation and extracellular matrix accumulation via foxo1 in mice with diabetic nephropathy. Cell Prolif. 2018, 51, e12462. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Xue, H.; Zhou, L.; Qu, L.; Li, C.; Wang, Z.; Ni, J.; Yu, C.; Yao, T.; Huang, Y.; et al. Rescue of mesangial cells from high glucose-induced over-proliferation and extracellular matrix secretion by hydrogen sulfide. Nephrol. Dial. Transplant. 2011, 26, 2119–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornigold, N.; Johnson, T.S.; Huang, L.; Haylor, J.L.; Griffin, M.; Mooney, A. Inhibition of collagen i accumulation reduces glomerulosclerosis by a hic-5-dependent mechanism in experimental diabetic nephropathy. Lab. Investig. 2013, 93, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ma, Y.; Sun, Z.; Zhu, X. Tangeretin inhibits high glucose-induced extracellular matrix accumulation in human glomerular mesangial cells. Biomed. Pharmacother. 2018, 102, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Catania, J.M.; Chen, G.; Parrish, A.R. Role of matrix metalloproteinases in renal pathophysiologies. Am. J. Physiol. Ren. Physiol. 2007, 292, F905–F911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Xiao, L.; Xiao, P.; Yang, S.; Chen, G.; Liu, F.; Kanwar, Y.S.; Sun, L. A glimpse of matrix metalloproteinases in diabetic nephropathy. Curr. Med. Chem. 2014, 21, 3244–3260. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Pan, Q.; Chen, Y.; Yang, X.; Zhao, B.; Jia, L.; Zhu, Y.; Zhang, B.; Gao, X.; Li, X.; et al. Administration of danhong injection to diabetic db/db mice inhibits the development of diabetic retinopathy and nephropathy. Sci. Rep. 2015, 5, 11219. [Google Scholar] [CrossRef] [PubMed]
- McLennan, S.V.; Kelly, D.J.; Cox, A.J.; Cao, Z.; Lyons, J.G.; Yue, D.K.; Gilbert, R.E. Decreased matrix degradation in diabetic nephropathy: Effects of ace inhibition on the expression and activities of matrix metalloproteinases. Diabetologia 2002, 45, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.G.; Thompson, J.C.; Yoder, M.H.; Charnigo, R.; Tannock, L.R. Prevention of renal apob retention is protective against diabetic nephropathy: Role of tgf-beta inhibition. J. Lipid Res. 2017, 58, 2264–2274. [Google Scholar] [CrossRef] [PubMed]
- Jie, L.; Pengcheng, Q.; Qiaoyan, H.; Linlin, B.; Meng, Z.; Fang, W.; Min, J.; Li, Y.; Ya, Z.; Qian, Y.; et al. Dencichine ameliorates kidney injury in induced type ii diabetic nephropathy via the tgf-beta/smad signalling pathway. Eur. J. Pharmacol. 2017, 812, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Koga, K.; Yokoi, H.; Mori, K.; Kasahara, M.; Kuwabara, T.; Imamaki, H.; Ishii, A.; Mori, K.P.; Kato, Y.; Ohno, S.; et al. Microrna-26a inhibits tgf-beta-induced extracellular matrix protein expression in podocytes by targeting ctgf and is downregulated in diabetic nephropathy. Diabetologia 2015, 58, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Chen, X.M.; Poronnik, P.; Pollock, C.A. Transforming growth factor-beta/connective tissue growth factor axis in the kidney. Int. J. Biochem. Cell Biol. 2008, 40, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Huang, X.R.; Wang, W.S.; Li, J.H.; Heuchel, R.L.; Chung, A.C.K.; Lan, H.Y. The protective role of smad7 in diabetic kidney disease: Mechanism and therapeutic potential. Diabetes 2011, 60, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Luangmonkong, T.; Suriguga, S.; Adhyatmika, A.; Adlia, A.; Oosterhuis, D.; Suthisisang, C.; de Jong, K.P.; Mutsaers, H.A.M.; Olinga, P. In vitro and ex vivo anti-fibrotic effects of ly2109761, a small molecule inhibitor against tgf-beta. Toxicol. Appl. Pharmacol. 2018, 355, 127–137. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Primer Sequence |
| Fibronectin | XM_017315848.1 | Forward: 5′-GCCGAATGTAGATGAGGAG-3′ Reverse 5′-AGGAACTTGGAACTGTAAGG-3′ |
| Collagen I | NM_007742.4 | Forward: 5′-TGTGTGCGATGACGTGCAAT-3′ Reverse 5′-GGGTCCCTCGACTCCTACA-3′ |
| MMP-2 | NM_008610.3 | Forward: 5′-TGGCAAGGTGTGGTGTGCGAC-3′ Reverse 5′-TCGGGGCCATCAGAGCTCCAG-3′ |
| MMP-9 | NM_013599.4 | Forward: 5′-TCTTCGACTCCAGTAGACAATCCTT-3′ Reverse 5′-AATTGGCTTCCTCCGTGATTCG-3′ |
| β-actin | NM_007393.5 | Forward: 5′-TCCATCATGAAGTGTGACGT-3′ Reverse 5′-GAGCAATGATCTTGATCTTCAT-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.; Sun, Y.; Wei, G.; Li, S.; Zhao, Z. Ergosterol Ameliorates Diabetic Nephropathy by Attenuating Mesangial Cell Proliferation and Extracellular Matrix Deposition via the TGF-β1/Smad2 Signaling Pathway. Nutrients 2019, 11, 483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020483
Dong Z, Sun Y, Wei G, Li S, Zhao Z. Ergosterol Ameliorates Diabetic Nephropathy by Attenuating Mesangial Cell Proliferation and Extracellular Matrix Deposition via the TGF-β1/Smad2 Signaling Pathway. Nutrients. 2019; 11(2):483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020483
Chicago/Turabian StyleDong, Zhonghua, Yueyue Sun, Guangwei Wei, Siying Li, and Zhongxi Zhao. 2019. "Ergosterol Ameliorates Diabetic Nephropathy by Attenuating Mesangial Cell Proliferation and Extracellular Matrix Deposition via the TGF-β1/Smad2 Signaling Pathway" Nutrients 11, no. 2: 483. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020483