The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health


Biologie des Oiseaux et Aviculture, INRA, Université de Tours, 37380 Nouzilly, France
*
Author to whom correspondence should be addressed.
Nutrients 2019, 11(3), 684; https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030684
Submission received: 15 February 2019
/
Revised: 15 March 2019
/
Accepted: 19 March 2019
/
Published: 22 March 2019
(This article belongs to the Special Issue Egg Intake and Human Health)
Abstract
:Egg is an encapsulated source of macro and micronutrients that meet all requirements to support embryonic development until hatching. The perfect balance and diversity in its nutrients along with its high digestibility and its affordable price has put the egg in the spotlight as a basic food for humans. However, egg still has to face many years of nutritionist recommendations aiming at restricting egg consumption to limit cardiovascular diseases incidence. Most experimental, clinical, and epidemiologic studies concluded that there was no evidence of a correlation between dietary cholesterol brought by eggs and an increase in plasma total-cholesterol. Egg remains a food product of high nutritional quality for adults including elderly people and children and is extensively consumed worldwide. In parallel, there is compelling evidence that egg also contains many and still-unexplored bioactive compounds, which may be of high interest in preventing/curing diseases. This review will give an overview of (1) the main nutritional characteristics of chicken egg, (2) emerging data related to egg bioactive compounds, and (3) some factors affecting egg composition including a comparison of nutritional value between eggs from various domestic species.
1. Introduction
In 1968, the egg industry had to face American Heart Association recommendations that encouraged people to consume less than three whole eggs per week claiming that high dietary cholesterol equals high blood cholesterol and consequently higher cardiovascular disease risks. These recommendations impacted, not only the egg industry, but also partly influenced people’s dietary habits depriving them from an affordable food of high nutritional interest. By 1995, there was a concerted effort to unify all the US national dietary recommendations and to support in vitro and in vivo research to rehabilitate eggs [1]. Half a century of research has now demonstrated that egg intake is not associated with increased health risk [2] and that it is worth incorporating such product in our diet with regard to its high nutrients content and its numerous bioactivities [1]. Some recent researches have highlighted the beneficial role of eggs for humans, including physically active people, and several authors have demonstrated that egg cholesterol was not well absorbed [3,4]. Consequently, consuming eggs does not significantly impact blood cholesterol concentration [3,4]. In parallel, egg consumers especially 6–24 month-old infants eat lower added and total sugars relative to non-consumers [5], which is likely correlated with its satiety effect [2,6,7]. It is now well established that egg can contribute to overall health across the life span, although people suffering from metabolic disorders such as diabetes, hypercholesterolemia, and hypertension still need to take caution with their dietary cholesterol intake [8]. Another concern relates to egg allergy, which is a common infant food allergy with a prevalence estimated to be between 1.8% and 2% in children younger than five years of age. Molecules that are associated with hypersensitivity to eggs are mainly concentrated in egg white, with ovalbumin, lysozyme, ovomucoid, and ovotransferrin being the major egg allergens [9]. Some yolk-derived proteins have also been reported [9]. Egg allergy usually develops within the first five years of life, with 50% of children outgrowing egg hypersensitivity by three years [10,11]. Fortunately, in most cases, the prevalence of egg allergy decreases with age [12] and usually, it resolves by school age.
Eggs are of particular interest from a nutritional point of view, gathering essential lipids, proteins, vitamins, minerals, and trace elements [13], while offering a moderate calorie source (about 140 kcal/100 g), great culinary potential, and low economic cost. Indeed, eggs have been identified to represent the lowest-cost animal source for proteins, vitamin A, iron, vitamin B12, riboflavin, choline, and the second lowest-cost source for zinc and calcium [14]. In addition to providing well-balanced nutrients for infants and adults, egg contains a myriad of biologically active components [15,16,17]. These components are allocated in the various internal egg components (Figure 1). It has to be mentioned that eggshell and its tightly associated eggshell membranes are usually not consumed, although eggshell membranes are edible (Figure 1). The average consumption of eggs/year/capita in the world ranges from 62 (India) to more than 358 (Mexico) [18] and is even less in African Countries (36 eggs/year/capita) ([19]. Table eggs that are commercialized are not fertilized and are produced by about 3 billion hens, specifically bred throughout the world for human consumption.
Egg components are also reported to be highly digestible although a small amount of egg proteins is not assimilated [20], especially when egg is consumed as a raw ingredient [20,21,22]. The higher digestibility of cooked egg proteins results from structural protein denaturation induced by heating, thereby facilitating hydrolytic action of digestive enzymes. However, although the assimilation of egg protein is facilitated by heat-pretreatment and at a high level (91–94% for cooked egg-white proteins), it remains partly incomplete. It is noteworthy that major proteins, essentially egg-white proteins such as the proteinase inhibitor ovomucoid, and major egg-white ovalbumin resist thermal heating [23,24]. This observation is particularly interesting, knowing that egg-derived proteins and many hydrolytic peptides generated in vitro from limited digestion of egg-white proteins possess biological activities of interest for human health and may thus be used as nutraceuticals [16]. Indeed, several of those have been shown to exhibit antimicrobial, antioxidant, and anti-cancerous properties [25,26,27]. Thus, many authors have highlighted the importance of protein-derived peptides in the gut and their substantial role in the body’s first line of immunological defense, immune-regulation, and normal body functioning [28].
2. Egg Nutrients
Egg proteins are distributed equally between egg white and egg yolk, while lipids, vitamins, and minerals are essentially concentrated in egg yolk (Figure 2). Water constitutes the major part of egg (Figure 2) and it is noteworthy that the egg is devoid of fibers. The relative content of egg minerals, vitamins, or specific fatty acids may vary from one national reference to another [29] but remains globally comparable when considering major constituents such as water, proteins, lipids, and carbohydrates. The major egg nutrients are, indeed, very stable and depend on the ratio of egg white to yolk in contrast to minor components, which are affected by several factors including hen nutrition (See Section 4.2). In a whole, raw, and freshly laid egg, water, protein, fat, carbohydrates, and ash represent about 76.1%, 12.6%, 9.5%, 0.7%, and 1.1%, respectively [30].
2.1. Macronutrients
2.1.1. Proteins
Egg white and egg yolk are highly concentrated in proteins. Hundreds of different proteins have been identified and are associated with specific physiological functions to fulfill time-specific requirements during embryo development. The compartment-specificity of some of these proteins can be explained by the fact that egg yolk and egg white are formed by distinct tissues. Egg yolk has essentially a hepatic origin, while egg white is synthesized and secreted after ovulation of the mature yolk in the hen’s oviduct [31].
The concentration of proteins is, on average, 12.5 g per 100 g of whole raw fresh egg, while egg yolk with its vitelline membrane and egg white contain 15.9 g protein and 10.90 g protein per 100 g, respectively. These values are slightly modified by hen genetics and age (See Section 4). Thanks to complementary proteomic approaches, nearly 1000 different proteins have been identified in the chicken egg, including the eggshell [32,33,34,35,36,37,38,39,40].
Yolk is a complex milieu containing 68% low-density lipoproteins (LDL), 16% high-density lipoproteins (HDLs), 10% livetins and other soluble proteins, and 4% phosvitins. These components are distributed between non-soluble protein aggregates called granules (19–23% of dry matter), which account for about 50% of yolk proteins, and a clear yellow fluid or plasma, that corresponds to 77–81% of dry matter [41,42]. Apolipoprotein B, apovitellenin-1, vitellogenins, serum albumin, immunoglobulins, ovalbumin, and ovotransferrin are the most abundant proteins of egg yolk, representing more than 80% of total egg-yolk proteins [43]. Yolk is tightly associated with the vitelline membranes, which consist of two distinct layers [44] that form an extracellular protein matrix embracing the yolk. These membranes provide to the yolk a physical separation from other egg compartments and prevents subsequent leakage of egg yolk towards egg white.
The egg white is a gel-like structure that lacks lipids and is composed mainly of water (about 88%) [44] (Figure 2), fibrous structural proteins (ovomucins), glycoproteins (ovalbumin, protease inhibitors), antibacterial proteins (lysozyme), and peptides (see Section 3.1) [33,45]. The average volume of egg white is estimated to be 30 mL (for an egg weighting 60 g, eggshell included) and protein concentration is about 110 mg/mL of egg white. In total, 150 distinct proteins have been identified in egg white [35], knowing that the very abundant ovalbumin accounts for 50% of the total egg-white proteins. The physiological function of this protein in egg remains unknown but ovalbumin is assumed to provide essential amino-acids for the chicken embryo growth. Egg-white ovalbumin thus represents a valuable source of amino-acids for human nutrition. Besides ovalbumin, egg white is concentrated in antibacterial lysozyme that is currently used as an anti-infectious agent in many pharmaceuticals and as food preservative (see Section 3.1). The viscous aspect of egg white is essentially due to ovomucin [46]. Remarkably, egg white is also characterized by the presence of four highly abundant protease inhibitors [47] that may delay digestion of egg components, especially when egg white is used as a raw ingredient in some food preparations.
2.1.2. Lipids
The total lipid content is relatively stable in the egg ranging from 8.7 to 11.2 per 100 g of whole egg, when considering various EU countries and USA egg composition tables [29]. These lipids are only concentrated in the egg yolk (Figure 2 and Table 1) and a small part may remain tightly associated with vitelline membranes [48,49].
Lipids are part of yolk lipoproteins whose structure consists of a core of triglycerides and cholesteryl esters, surrounded by a monolayer of phospholipids and cholesterol in which apoproteins are embedded [42]. It is very difficult to change total lipid content in egg. An increase of fat in egg essentially depends on the increase of yolk-to-egg-white ratio, which is, however, poorly affected by hen’s nutrition. In contrast, fatty acid profile strongly depends on hen’s diet (See Section 4.2). This variability is illustrated in Table 1 by minimal and maximal values of fatty acids (saturated, monounsaturated, and polyunsaturated). Noticeably, the relative amount of unsaturated (monounsaturated + polyunsaturated) to saturated fatty acids in yolk (5.31 g versus 2.64 g per 100g of whole egg, Table 1) is particularly high compared to other animal-derived food sources. Yolk is also a rich source of essential fatty acids such as linoleic acid (FA 18:2 9c,12c (n-6)). The high content of cholesterol in eggs (400 mg per 100 g of whole egg) has contributed to the decline of egg intake 30 to 40 years ago, although many studies conducted in the 1990s have reported an absence of correlation between egg intake and high level of plasmatic cholesterol [3,4]. It is now assumed that variation in plasmatic cholesterol and associated cardiovascular disease risk results from food factors but also saturated fatty acids intake (such as dietary myristic (14:0) and palmitic (16:0) acids). Old studies performed in vivo on monkeys and gerbils have shown that dietary 14:0 (myristic acid) and 18:2 (linoleic acid) are the main fatty acids modulating plasma cholesterol—14:0 was the main saturated fatty acid raising plasma cholesterol and 18:2 was the only fatty acid that consistently lowered it [50,51]. In egg, 14:0 (myristic acid, 0.024 g per 100 g whole egg) is much less concentrated compared to unsaturated fatty acids 18:2 (linoleic acid, 1.38 g per 100g whole egg). These data all concur to corroborate that egg is not associated with higher cardiovascular disease incidence in healthy patients. However, egg intake has to be controlled in hyper-responders to dietary cholesterol (about 15% to 25% of the population), as an increase in egg consumption in these people affects plasma lipids to a greater extent than in hypo-responders.
2.1.3. Carbohydrates
Egg does not contain any fibers and its content in carbohydrates is low (0.7%). Egg carbohydrates are distributed between egg yolk and egg white (Figure 2). Glucose is the dominant free sugar in the egg (about 0.37 g per 100 g of whole egg) and is essentially present in egg white (0.34 g per 100 g of egg white versus 0.18 g per 100 g of egg yolk) [30]. Trace amounts of fructose, lactose, maltose, and galactose have been detected in raw egg white and raw egg yolk [30]. Carbohydrates are also highly represented in egg proteins, knowing that many of them are glycoproteins undergoing post-translational glycosylations prior to secretion by reproductive tissues of the hen to form yolk, membranes, and egg white.
2.2. Micronutrients
2.2.1. Vitamins and Choline
The egg and, more precisely, the egg yolk, is a vitamin-rich food that contains all vitamins except vitamin C (ascorbic acid). The absence of vitamin C in the egg may result from the fact that birds are capable of satisfying their own vitamin C requirements, by de novo synthesis from glucose [52]. The ability to produce vitamin C has been lost during the process of evolution in several animal species including guinea pigs, monkeys, flying mammals, humans, and some evolved passerine birds [52]. Consequently, these latter species, but not domestic birds, are dependent on dietary sources of vitamin C (fruits and vegetables). The egg yolk contains high amount of vitamin A, D, E, K, B1, B2, B5, B6, B9, and B12, while egg white possesses high amounts of vitamins B2, B3, and B5 but also significant amounts of vitamins B1, B6, B8, B9, and B12 (Table 2). Eating two eggs per day covers 10% to 30% of the vitamin requirements for humans. It is noteworthy that the content of liposoluble vitamins (vitamins A, D, E, K) in egg yolk is highly dependent on the hen’s diet (See Section 4.2). In addition to these vitamins, eggs represent a major source of choline, which is essentially concentrated in the yolk (680 mg/100 g in the egg yolk versus 1 mg/100 g in the egg white) [30,53]. It has been reported that hard-boiled egg represents the second major source of choline after beef liver [54] and the first source of choline in the US diet [55]. In foods, choline is found as both water-soluble (free choline, phosphocholine, and glycerophosphocholine) and lipid-soluble forms (phosphatidylcholine and sphingomyelin) and has important and diverse functions in both cellular maintenance and growth across all life stages. It plays some roles in neurotransmission, brain development, and bone integrity [54,56,57].
2.2.2. Minerals and Trace Elements
Egg is rich in phosphorus, calcium, potassium, and contains moderate amounts of sodium (142 mg per 100 g of whole egg) (Table 3). It also contains all essential trace elements including copper, iron, magnesium, manganese, selenium, and zinc (Table 3), with egg yolk being the major contributor to iron and zinc supply. The presence of such minerals and micronutrients in egg is quite interesting as a deficiency in some of these (Zn, Mg, and Se) has been associated with depression and fatigue [59] and development of pathological diseases. The concentration of some of those trace elements (selenium, iodine) may be significantly increased depending on hen’s diet (See Section 4.2).
2.3. Antinutritional Factors
As mentioned above, major proteins of egg include protease inhibitors that may delay the proper degradation of egg proteins by inhibiting digestive enzymes including pepsin, trypsin, and chymotrypsin. Indeed, egg white is a major source of ovostatin, ovomucoid, ovoinhibitor, and cystatin [47]. Moreover, some of these molecules (ovoinhibitor, ovomucoid, cystatin) possess many disulfide bonds that are likely to confer moderate resistance to denaturation by proteases and gastric juices. Some of these antinutritional factors may be partly denatured by heat [20,22,23,24] during the process of cooking, thus facilitating protein access to digestive proteases. Moreover, some vitamin-binding proteins highly concentrated in egg may also limit some vitamin access—avidin that binds vitamin B12 (biotin) exhibits the highest known affinity in nature between a ligand and a protein [60]. The bioavailability of biotin for consumers may be compromised by the tight complex formed between avidin and its bound vitamin B8.
3. Egg Nutraceuticals
There is increasing evidence that egg is not solely a basic food of high nutritional value but that it also contains many bioactive compounds (lipids, vitamins, proteins, and derived hydrolytic peptides) [16,61,62,63,64] of major interest for human health. In vitro analyses performed on purified proteins have revealed a great potential in egg proteins as they exhibit a diversity of biological activities. Various tools combining physicochemical, analytical, and in silico approaches [65,66] can be used to identify hydrolytic peptides with potential bioactivities. It is remarkable that a lot of egg proteins have no identified physiological function described yet, besides providing essential amino-acids for the embryo but also for egg-eating species including humans. In addition to egg proteins displaying a wide spectrum of antimicrobial activities that could contribute to intestine health, many efforts have been made in the last decades to further characterize biological activities of egg-derived hydrolytic peptides that may naturally occur during the digestive process [20,22]. Interestingly, some of these bioactive peptides are specifically generated after limited proteolysis of denatured egg proteins [67], after boiling. Most of these studies were performed in vitro, but this finding opens many fields of research. To date, little is known on how egg proteins resist acidic pH of the stomach, digestive proteases, and intestinal microbiote, and how the presence of egg protease inhibitors in the diet can interfere with the degradation of egg proteins by digestive proteases. The kinetics of protein digestion is sequential, starting with the hydrolysis of proteins into peptides until complete degradation into dipeptides and, finally, free amino acids. But it is known that some egg proteins (ovalbumin, ovomucoid) are only partly digested [20,22] suggesting that some bioactive peptides may be generated naturally without undergoing complete degradation into amino acids.
3.1. Antimicrobials
Egg antimicrobials in edible parts are essentially concentrated in egg white and the vitelline membrane. Depending on the protein considered, these antimicrobials may exhibit antibacterial, antiviral, antifungal, or antiparasitic activities (Table 4).
Their antibacterial effect relies on several bactericidal or bacteriostatic mechanisms. Some of them have a powerful activity via interaction with bacterial walls that further triggers permeabilization and bacterial death (lysozyme, avian beta-defensins, etc.). The effects of the other molecules are rather indirect by decreasing the bioavailability of iron (ovotransferrin) and vitamins (avidin) that are required for some microbial growth, and by inhibiting microbial proteases that are virulent factors of infection (ovoinhibitor, cystatin) [68]. The various egg antimicrobial molecules that have been described so far in literature are listed in Table 4. Interestingly, some of them (AvBD11, OVAX, avidin, beta-microseminoprotein) are not expressed in the human genome [69], suggesting that they may constitute powerful anti-infectious agents against human enteric pathogens, to reinforce intestinal host immunity.
In addition to these egg proteins and peptides, there are increasing data reporting the antimicrobial activity of egg-derived peptides that may be released after partial hydrolysis by exogenous proteases. Such hydrolytic peptides obtained from lysozyme [70,71,72,73], from ovotransferrin [25], from ovomucin [74], and from cystatin [75] have shown a broad range of antibacterial activities.
3.2. Antioxidant Activities
Long-term oxidative stress in the gastrointestinal tract can lead to chronic intestinal disorders and there is increasing interest in investigating the potential of food-derived antioxidants, including egg antioxidants, in intestinal health. Chicken egg contains many antioxidant compounds that encompass vitamins, carotenoids, minerals, and trace elements but also major egg-white proteins [103,104,105,106] such as ovotransferrin, in its native form or as hydrolytic peptides [98,99,104,105,107,108,109,110], ovomucoid and ovomucoid hydrolysates [111,112], ovomucin hydrolysates and derived peptides [112], and egg yolk-proteins including phosvitin [113]. Most of these molecules have been generated in vitro but some assays performed in a porcine model have revealed the beneficial effect of proteins derived from egg yolk in reducing the production of pro-inflammatory cytokines [114]. The authors concluded that supplementation of the diet with egg yolk-proteins may be a novel strategy to reduce intestinal oxidative stress [114].
3.3. Anti-Cancerous Molecules
There are only few data showing that food-derived proteins and peptides can also be beneficial to prevent and to cure cancer diseases [26]. Several studies have confirmed the tumor-inhibitory activity of egg white lysozyme using experimental tumors. Its effect essentially relies on immunopotentiation [115]. Ovomucin (beta subunit) and ovomucin-derived peptides also showed anti-tumor activities via cytotoxic effects and activation of the immune system [74]. The anticancerous effect of egg tripeptides [27] and hydrolytic peptides from ovotransferrin [116] have also been published. Information in this field is quite scarce, but it may be worth continuing to investigate such activities. Some interesting data may arise from studies on egg protease inhibitors [47] since similar molecules existing in other food product, including legumes like pea, have been described as potential colorectal chemopreventive agents [117].
3.4. Immunomodulatory Activities
Several egg proteins have potential immunomodulatory activities. Among these, egg-white lysozyme is a promising agent for the treatment of inflammatory bowel disease. In a colitis porcine model, lysozyme was demonstrated to significantly protect animals from colitis and reduce the local expression of pro-inflammatory cytokines while increasing the expression of the anti-inflammatory mediators [118]. Sulfated glycopeptides generated by proteolysis from ovomucin, chalazae, and yolk membrane can exhibit macrophage-stimulating activities in vitro [74]. Cytokines, such as egg-white pleiotrophin, play a pivotal role in the generation and resolution of inflammatory responses. In human, pleiotrophin have been shown to promote lymphocyte survival, and to drive immune cell chemotaxis [119,120]. But, the biological significance of the potential immunomodulatory activity of egg white pleiotrophin in human intestine remains very speculative. In contrast, some valuable immunomodulatory activities might emerge from ovotransferrin and egg yolk vitellogenin hydrolysates [121,122] after partial degradation by digestive proteases.
3.5. Antihypertensive Activities
Considering the prevalence and importance of hypertension worldwide (over 1.2 billion individuals) [123], there is increasing ongoing research to find ways to regulate this multifactorial disease. At the population level, the most important factors of long-term control of blood pressure are sodium and potassium intakes and the importance of the renin-angiotensin-aldosterone system. Most egg-derived peptides with anti-hypertensive activities exhibit inhibitory activities against the angiotensin-converting enzyme (ACE). This enzyme triggers the processing and activation of angiotensin I into the active vasoconstrictor angiotensin II. Several yolk-derived peptides bearing antihypertensive activities have been described in the literature [113,124] along with ovotransferrin and egg white hydrolysates [125,126]. Some of these peptides contain only three amino-acids [27,127]. Some of these tripeptides were demonstrated to be active in vivo—the oral administration of these peptides that have been administrated orally in hypertensive rats contributed to significantly reduce blood pressure [128] and thus, may help in diminishing the occurrence of cardiovascular diseases [127,129].
4. Factors Affecting Egg Quality
4.1. Genetics
Selection for egg quality is an important component of the breeding strategies of companies that market egg-laying type hens. Indeed, consumers demand high quality products with strong eggshell, while reducing cost, guaranteeing eggs devoided of microbial contaminants, and improving the acceptability of rearing systems [130,131]. Most selection strategies to improve egg quality has focused on the shell’s physical properties (and its ability to resist shocks), stability of egg weight, egg-white quality, and yolk percentage. Egg-white quality essentially refers to albumen height that reflects freshness, and its ability to prevent microbial growth and survival that may be associated with toxi-infection risks for consumers (Salmonellosis). Recently, some authors reported differences in albumen height/pH in various selected lines [132] and corroborated that selection on specific traits has changed the proportion of the yolk, albumen, and shell while increasing albumen height. The moderately high variability in weights of egg yolk and albumen was also observed when comparing selected and traditional lines [133]. Egg white is a very unfavorable medium for bacteria, due to its high viscosity, its pH becoming progressively alkaline upon egg storage (7.8 to 9.5), and to the presence of a myriad of antimicrobial molecules (See Section 3.1). In fact, the antibacterial potential of egg white was demonstrated to be moderately inheritable [134]. Concerning egg proteins and peptides, some difference in relative abundance of certain molecules have been reported in brown versus white eggs or in different lines, but the major egg proteins remain basically unchanged [135,136].
4.2. Nutrition and Rearing Systems
Nutrition of laying hens, the characteristics of the feed (composition in nutrients, energy content, but also feedstuff texture and presentation), and the mode of feed delivery throughout the day affect, not only egg weight, but also to a lesser extent, egg-yolk and egg-white proportion [137]. The quality of the diet for pullets will influence the egg weight essentially at the onset of lay but it is much less significant when considering the whole period of lay [137]. The dietary characteristics include the level of calcium supply as well as its particular size. Dietary calcium in a particular form allow hens to express a specific appetite for calcium at the end of the day, that is stored and further assimilated during the night when the shell formation take place [138]. The egg weight is influenced by the daily energy consumption of hens. High energetic diet and the dietary supply of linoleic acid increase the egg weight. This effect is particularly relevant at the onset of laying (22–32 weeks) and much less pronounced in older hens [139]. Egg weight is also increased by the level of dietary proteins, and some studies have revealed that methionine was the main limiting amino acid as its presence in the hen’s diet is positively correlated with egg weight [140]. Additionally, energy intake depends on the protein source considering that laying hens are traditionally fed corn, wheat, and soybean meal. The presence of antinutritional factors in the diet (protease inhibitors and proteins that are highly resistant to digestive proteases such as convicilin, glycinin, cruciferin [141]) may affect the overall digestibility of the feed by the hens and subsequent egg weight. However, the content in major egg components is relatively stable and the variability depends essentially on the proportion of the albumen to the yolk, which exhibit very contrasted composition (See Section 2). In contrast, the fatty-acid profile of the yolk and the content of micronutrients such as vitamins and trace minerals (See Section 2.2) or of carotenoids are very variable and directly rely on diet composition [137].
The fatty-acid profile of an egg, incorporated in triglycerides and phospholipids, directly reflects the composition of fatty acids in the hen’s diet. In contrast, the enrichment of the hen’s diet with saturated fatty acids has less influence of the yolk lipid profile. The saturated and unsaturated fatty-acid content of the hen’s diet can be modified by the inclusion of oil or feedstuffs showing a high degree of unsaturated fatty acid such as fish, chia, flaxseeds [142,143], olive, or soy oils (see Reference [144] for a review). For instance, including olive oil in a hen’s diet favors the incorporation of mono-unsaturated fatty acids (in particular, oleic acid content) in the yolk, while soil oil increases that of unsaturated n-6 fatty acids (linoleic acid) [144]. More recently, results obtained with an enrichment of the diet with microalgae or linseeds have revealed the potential of these compounds to increase the content of yolk in n-3 fatty acids (more than 3- and 4-fold increase, respectively) [145]. A similar trend in increasing the polyunsaturated content of yolk was observed with marigold extract powder [146], Schizochytrium microalgae [147], prebiotic and probiotic combination [148], etc. To conclude, it is relatively easy to enrich the egg in some unsaturated fatty acids, of interest for human health. The challenge remains to identify animal and vegetable sources of polyunsaturated fatty acids that increase the content of these fatty acids in yolk without affecting its technological and/or sensorial quality, to suit both the food industry requirements (egg products) and consumer demand.
The content of some trace minerals in eggs such as selenium, iodine, and, at lower magnitudes, iron, zinc, fluoride, or magnesium can also be increased by larger dietary supply for hens [149]. The average selenium egg content is around 5 µg per egg and can be increased 3- to 6-fold (12-fold in the albumen and 4-fold in the yolk) and reach 30–40 µg/egg when hens are supplied with 0.3 to 0.5 mg selenium (from selenomethionine or selenium enriched yeast/kg diet). Such egg enrichment provides 50–70% of the daily human requirement [150].
Similarly, hen nutrition is a way to enrich egg in lipophilic vitamins (A, D, E, K) or water-soluble vitamins (folate, B12, pantothenic acid, and, at lower magnitudes, riboflavin, thiamin, and biotin). Egg content in vitamin A can be enhanced 10-fold from its initial value when hens are supplied with 30,000 IU retinol and that of vitamin D3 by 15-fold (2–5 to 34 µg/100g in hens fed 2500 and 15,000 IU D3). Vitamin E content in the yolk can increase 3- to 20-fold depending on basal diet content and dietary supply. For water-soluble vitamins, the magnitude of increase by larger dietary supply is lower—more than 2-fold for folate, riboflavin, or cobalamin and, at a lower level, thiamin, biotin and panthotenic acid, pyridoxine, or niacin [137]. Yolk color (yellow/orange shading) is also determined by the content in carotenoids in the diet [137]. The main sources of carotenoids (lutein, xanthophylls, and zeaxanthin) for birds are corn, lucerne, flower (marigold), and paprika extract (red carotenoids) that are incorporated in a hen’s diet to meet consumer demand for a more orange-yellow yolk. Beside its interest in increasing the visual aspect of the yolk, the high content of carotenoids in the yolk may also have a positive incidence for human health in increasing visual performance and reducing the risk of age-related macular degeneration [151].
Because birds in free-rearing systems have access to grass, insects, and worms in addition to their basic diet, the content in some egg micronutrients may also slightly vary. As an example, free rearing results in significantly higher total tocopherol, alpha-tocopherol, and lutein contents, as compared to the battery cage and the organic system, respectively, when hens are fed a similar conventional diet [152]. Conversely, no significant differences were observed in the lipid and total sterol contents [152]. A decrease in egg-white height and in yolk color was also observed when comparing egg from conventional cages to free-range systems [153]. However, eggs from conventional systems contain, in general, more carotenoids and vitamins due to the possibility of including chemical additives in the diet, knowing that such practice is not conducted in organic systems. In parallel, since the immune system of the hens is likely to be more challenged by the presence of environmental microbes in free-range systems, an increase of immunoglobulin Y content in egg yolk (initially to provide some passive immunity for the chick, similarly to maternal colostrum for infants) is also likely to increase. Furthermore, some have shown that the antimicrobial capacity of egg white may also be slightly modulated when hens are exposed to environmental microbes [154]. To conclude, rearing laying hens in free-range systems may globally improve the antimicrobial potential of eggs (See Section 3.1).
4.3. Physiological Status
Egg production and quality are greatly influenced by the physiological status of the hen—age, stress, and immune status [155]. Egg weight varies from 50 g to 70 g depending mainly on hen’s age and genetics (See Section 4.1). In modern flock, the egg weight has been restricted to 62–66 g throughout the laying cycle (20 to 80 weeks of age). Egg weight is the primary criterion used in egg grading (small, medium, large, extra-large). The increase of egg weight observed in older hens is associated with an increase in albumen and yolk average weights and in relative yolk proportion [156]. Hen age is also associated with a decrease in eggshell strength, in albumen height (the higher albumen height, the higher the freshness grade) [153], and a decrease in the strength of vitelline membrane that is frequently associated with a higher incidence of yolk rupture [157]. This latter observation is likely to result from an increase in yolk proportion in older hens. However, when considering the chemical composition of eggs, the results are quite controversial, although some have shown some difference in fatty-acid composition (docosahexaenoic acid and arachidonic acid) of egg depending on hen’s age [158]. Overall, these alterations in egg quality with age are, however, consistent with physiological changes and maybe some primary metabolic dysfunctions occurring in commercial hens at the end of the laying cycle [159].
Egg-laying performance also depends on general hen’s health as diseases and infections can induce loss of appetite and physiological failure thereby affecting animal growth, egg laying, and egg quality (eggshell deformation, eggshell defects, egg-white thinning, etc.). The most commonly found microbes in laying hens that affect hygienic egg quality are Salmonella enterica Enteritidis, mycoplasma, infectious bronchitis virus, and avian influenza virus [160]. Another major issue for hen welfare and egg quality is the poultry red mite that is found in most rearing systems for laying hens, regardless of the system type [161]. As a blood-feeder, this mite has dramatic effects on fowl host welfare including distress, anemia, reduced egg production, and reduced egg quality [162]. The prevalence of red mites is expected to increase, as a result of recent hen husbandry legislation changes (in favor of non-cage systems), increased acaricide resistance, climate warming, and the absence of efficient and sustainable solutions to control infestations [163]. It is also a major concern for public health as it may be a vector for food-borne pathogens including Salmonella species [164]. Amongst all these microbes, Salmonella enterica Enteritidis is the most critical for egg consumers as this pathogen can survive in egg white [165], even after several weeks of storage at 4 °C and 25 °C [166] and be responsible for food-borne diseases. It remains the dominant pathogen associated with egg consumption [167]; but, considerable efforts have been made by authorities to control Salmonella Enteritidis dissemination throughout the egg production chain. Reported salmonellosis case numbers continued to decrease thanks to the implementation of successful Salmonella control programs in poultry production [168], including Salmonella detection and monitoring [169], the establishment of pre-harvest measures [170], management and sanitation measures [171], and egg decontamination by washing under certain conditions [172]. It is noteworthy that European legislation does not allow egg washing in Europe (Commission Regulation (EC) No 589/2008), “because of the potential damage to the physical barriers, such as the cuticle, may favor trans-shell contamination with bacteria and moisture loss and thereby increase the risk to consumers, particularly if subsequent drying and storage conditions are not optimal”.
4.4. Egg Storage and Heat Treatment
Shell eggs are stored at room temperature or preferably in the fridge prior to be used by consumers (eggs are considered as “fresh” up to 28 days after laying). The conditions of egg storage can induce deep internal changes including physicochemical modifications that may increase some technological properties that are useful for the food industry, and alteration of antibacterial properties of the egg white [173,174,175,176] (Figure 3). These alterations result from water exchange between yolk and the egg white and from water and carbon dioxide loss through the eggshell pores, which elicited an increase in the air cell that develops between the two eggshell membranes (Figure 3a). Albumen height decreases with time of storage while albumen pH and whipping volume increase [136]. In parallel, the strength of vitelline membrane decreases upon egg storage due to loosening thereby impacting yolk shape/index (the yolk becomes flat and its diameter is higher) [173]. These latter modifications favor egg white/egg yolk exchanges of components such as carbohydrates and glucose [177], proteins [178,179,180], vitamins, and trace elements [181]. In addition, storage duration and conditions are associated with protein degradation [175,179,180] and a decrease in its antibacterial potential [175]. However, except for proteins, there is only little information available that describes the changes/denaturation in lipids, vitamins, and minerals composing both egg white and egg yolk during storage. It will be interesting to further investigate how these modifications impact the respective functional, nutritional, and technological properties of egg yolk and egg white (foaming, emulsifying properties, etc.). Recent data has demonstrated that the antioxidant activity of egg yolk was globally unchanged during six weeks of retail storage [182]. All these alterations of freshness criteria are accelerated at room temperature when compared to refrigerated conditions.
In addition to storage, one would expect that egg nutrients may also be modified during cooking. No clear evidence of minerals or vitamins denaturation could be observed when comparing fresh, soft-boiled, and hard-boiled egg (Table 5).
Actually, some data appear contradictory from one reference source to another (CIQUAL versus USDA, Table 5). Anyhow, it seems that the amount of polyunsaturated fatty acids, selenium, and vitamin A [21] tends to decrease upon cooking, especially in hard-boiled eggs (Table 5).
Noticeably, proteins undergo major conformational modifications upon cooking, even though their relative amount is not impacted by cooking (Table 5). This protein denaturation may be beneficial to inactivate antinutritional factors such as egg-white antiproteases but also to denature highly resistant proteins, thereby facilitating protease activity in the digestive tract. A higher digestibility of egg proteins may also contribute to limiting hypersensitivity to eggs in children [183,184]. Meanwhile, it has been shown that cooking significantly reduces the oxygen radical scavenging capacity (antioxidant potential) of egg yolk associated with free aromatic amino acids, lutein, and zeaxanthin [182], and also impacts lipids of yolk [185]. These observations corroborate that taking into account the food matrix and the way eggs are prepared is of major importance to appreciate egg digestibility and its associated nutritional and nutraceutical quality [186]. To conclude, it is fairly difficult to assess the beneficial/risk balance of cooking egg for human health as many molecules may be affected by cooking, while, in parallel, the heating process may increase the digestibility of egg proteins and potentially reveal potential new bioactive peptides [187,188]; but, it is worth mentioning that cooking eggs also allows for the elimination potential pathogens responsible for toxi-infections in consumers. In conclusion, taking into account all these data, the advice to retain most nutritional and nutraceutical benefits associated with egg would be to foster the consumption of poached or soft-boiled eggs, where the egg white is cooked (to inactivate antinutritional factors and potential pathogenic bacteria) while the egg yolk remains essentially raw (to preserve most vitamins, lipids, micronutrients, and some bioactive (antioxydant) molecules).
4.5. Variability between Avian Domestic Species
The table-egg market is dominated by chicken eggs in all countries. However, duck eggs are also widely consumed in some Asian countries. The major reason for this ascendancy of chicken eggs relies on several reasons—chickens are easy to handle and to rear and they have been selected for decades to lay close to 320 eggs a year. Conversely, geese, turkeys, and ducks are seasonal layers and require most specific sanitary and rearing conditions. Chicken eggs are also of reasonable size, not too big and larger than quail eggs, the latter being eaten occasionally as a gourmet ingredient.
Although egg compositions from traditional domestic species share common characteristics [189,190], they possess some significant differences in terms of energy that are mostly explained by the change in relative proportion of yolk to egg white (Figure 4). The energy (kcal/100g) for chicken, quail, duck, goose, and turkey eggs is 143, 158, 185, 185, 171, respectively. While the relative amount of proteins remains stable between species (about 13%), the lipid proportion varies from 9.5% (chicken) to more than 13% (duck, goose) (Figure 4, which explains most of the variation in their respective energy value. Yolks from duck and goose species have relatively higher fat content and higher percentage of yolk compared to chicken egg [189]. In parallel, the lipid profile of egg yolk exhibits some specificities depending on species [191,192,193]. Overall, the composition of duck egg resembles that of goose egg, which is consistent with their phylogenetic proximity.
It is also noteworthy that the content of minerals and trace elements of chicken eggs are usually lower that those observed for the other species, particularly in duck and goose species (Table 6). A similar trend is observed for vitamins (Figure 5). However, it has to be mentioned that the change in vitamins and trace elements in eggs depends mainly on diet composition. Therefore, these differences might rather reflect conditions of bird rearing than genetic capacity of hens to retain these compounds in eggs.
Besides these chemical compounds, some variability in bioactive molecules were also reported. Indeed, the comparative analysis of egg-white and egg-yolk proteomes revealed some proteins that are specifically associated with one or the other species [194,195,196,197] and others (ovotransferrin, lysozyme, gallin), displaying difference in abundance [196]. Overall, these characteristics may affect the overall bioactive potential of egg-white extract (antibacterial and/or antioxidant activity) depending on its bird origin, and it was shown that egg white from chicken egg retains the highest antibacterial potential compared to egg white from turkey, duck, and goose, at least, against some bacterial strains (Bacillus subtilis, Staphylococcus aureus, Pseudomonas aeruginosa, and Escherichia coli) [91]. Furthermore, the lower abundance of ovomucoid (a protease inhibitor that is highly resistant to chemical and thermal denaturation) in egg white from duck, goose, or turkey [196] may be associated with higher protein digestibility.
To conclude, such differences in the chemical composition of egg, and also in some of its biologically active molecules, are likely to be correlated with an increased or lower nutraceutical value of some egg proteins from other species, compared with the chicken egg.
5. Conclusions
For ages, eggs have been considered as foods of high nutritional value for humans and are widely consumed worldwide. Its consumption is predicted to continuously increase in the future, considering the growing number of occidental consumers who start to adopt a meat-free diet (vegetarians) or who significantly reduce their meat intake. This change in our consumption mode and food habits is motivated by much data on the association risk of meat intake with digestive cancers and cardiovascular diseases, and growing numbers of studies that praise vegetarian diet [198,199,200,201]. In parallel, it is also driven by ethical concerns and environmental issues with regards to modes of meat production [202]. It has also to be highlighted the existence of substantial disparities in egg consumption between countries [18], which is particularly low in Central Africa, with only 36 egg/year/capita [19]. The development of the egg industry in developing countries may represent a great opportunity for human nutrition/health and economy.
Besides basic nutrients, eggs are also a great source of potential nutraceuticals. A total of 550 distinct proteins were identified so far in egg-white and yolk/vitelline membranes and the physiological function of only 20 of them is characterized to date. This remark suggests that the egg probably still encloses many unknown activities that merit further investigations considering the current lack of research assessing the fate of egg proteins along the digestive tract. Such studies could help to better appreciate the in vivo potential of egg proteins and of the resulting hydrolytic peptides, and could be easily apprehended using dynamic gastric models that have been used with other foodstuffs, in food- and pharmaceutically-based research [203,204]. These in vitro models mimic both the biochemical and mechanical aspects of gastric digestion. They incorporate artificial saliva, compressive forces to disintegrate food, simulate continuous gastric emptying, and gastric secretion that generate pH profiles similar to human stomach. They also include bile salts and intestinal enzymes that act sequentially in a realistic time-dependent manner [205] and may be improved by adding intestine-like microbiota. This model was already used in a wide range of studies in order to assess the bioaccessibility of nutrients and to study the structural changes of food matrices. It is expected that such an experimental strategy would constitute a promising way to study the impact of egg preparation in the diet (raw versus cooked) on the physiological generation of bioactive peptides, and to better appreciate their biological significance for human health. It is well assumed now that gut health depends on the interplay between host genome, nutrition, and lifestyle that it contributes to normal brain function and mental health [206].
Author Contributions
S.R.-G. supervised the review, edited the chapter plan, and wrote the original draft of the article. N.G. was involved in the writing of the paragraph related to antimicrobials and Y.N. was in charge of the paragraph related to the nutritional value of eggs. All authors contributed to the critical review and the editing of the article.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McNamara, D.J. The Fifty Year Rehabilitation of the Egg. Nutrients 2015, 7, 8716–8722. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, X.; Thouvenot, P.; Belbraouet, S.; Chayvialle, J.A.; Hanesse, B.; Mayeux, D.; Debry, G. Effect of egg consumption in healthy volunteers: Influence of yolk, white or whole-egg on gastric emptying and on glycemic and hormonal responses. Ann. Nutr. Metab. 1996, 40, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Lopez Sobaler, A.M.; Aparicio Vizuete, A.; Ortega, R.M. Role of the egg in the diet of athletes and physically active people. Nutr. Hosp. 2017, 34, 31–35. [Google Scholar] [CrossRef]
- Kim, J.E.; Campbell, W.W. Dietary Cholesterol Contained in Whole Eggs Is Not Well Absorbed and Does Not Acutely Affect Plasma Total Cholesterol Concentration in Men and Women: Results from 2 Randomized Controlled Crossover Studies. Nutrients 2018, 10, 1272. [Google Scholar] [CrossRef]
- Papanikolaou, Y.; Fulgoni, V.L., 3rd. Egg Consumption in Infants is Associated with Longer Recumbent Length and Greater Intake of Several Nutrients Essential in Growth and Development. Nutrients 2018, 10, 719. [Google Scholar] [CrossRef]
- Missimer, A.; DiMarco, D.M.; Andersen, C.J.; Murillo, A.G.; Vergara-Jimenez, M.; Fernandez, M.L. Consuming Two Eggs per Day, as Compared to an Oatmeal Breakfast, Decreases Plasma Ghrelin while Maintaining the LDL/HDL Ratio. Nutrients 2017, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Stanciuc, N.; Banu, I.; Turturica, M.; Aprodu, I. pH and heat induced structural changes of chicken ovalbumin in relation with antigenic properties. Int. J. Biol. Macromol. 2016, 93, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Yang, F.; Zhang, Y.; Wang, T.; Chen, G. The Impact of Egg Nutrient Composition and Its Consumption on Cholesterol Homeostasis. Cholesterol 2018, 2018, 6303810. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Yang, M. Recent advances in the understanding of egg allergens: Basic, industrial, and clinical perspectives. J. Agric. Food Chem. 2008, 56, 4874–4900. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Pereira, B.; Voigt, K.; Grundy, J.; Clayton, C.B.; Higgins, B.; Arshad, S.H.; Dean, T. Prevalence and cumulative incidence of food hypersensitivity in the first 3 years of life. Allergy 2008, 63, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Osterballe, M.; Hansen, T.K.; Mortz, C.G.; Host, A.; Bindslev-Jensen, C. The prevalence of food hypersensitivity in an unselected population of children and adults. Pediatric Allergy Immunol. Off. Publ. Eur. Soc. Pediatric Allergy Immunol. 2005, 16, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.L.; Koplin, J.J.; Gurrin, L.C.; Dharmage, S.C.; Wake, M.; Ponsonby, A.L.; Tang, M.L.K.; Lowe, A.J.; Matheson, M.; Dwyer, T.; et al. The prevalence of food allergy and other allergic diseases in early childhood in a population-based study: HealthNuts age 4-year follow-up. J. Allergy Clin. Immunol. 2017, 140, 145–153.e148. [Google Scholar] [CrossRef] [PubMed]
- Nys, Y.; Sauveur, B. Valeur nutritionnelle des oeufs. INRA Prod. Anim. 2004, 17, 385–393. [Google Scholar]
- Drewnowski, A. The Nutrient Rich Foods Index helps to identify healthy, affordable foods. Am. J. Clin. Nutr. 2010, 91, 1095s–1101s. [Google Scholar] [CrossRef]
- Abeyrathne, E.; Ahn, D.U. Isolation of value-added components from egg white and their potential uses in food, nutraceutical and pharmaceutical industries. In Handbook of Eggs in Human Function; Watson, R.R., DeMeester, F., Eds.; Wageningen Acad Publ: Wageningen, The Netherlands, 2015; pp. 35–52. [Google Scholar]
- Kovacs-Nolan, J.; Phillips, M.; Mine, Y. Advances in the value of eggs and egg components for human health. J. Agric. Food Chem. 2005, 53, 8421–8431. [Google Scholar] [CrossRef] [PubMed]
- Anton, M.; Nau, F.; Guerin-Dubiard, C. Bioactive fractions of eggs for human and animal health. In Improving the Safety and Quality of Eggs and Egg Products, Vol 2: Egg Safety and Nutritional Quality; VanImmerseel, F., Nys, Y., Bain, M., Eds.; Woodhead Publ Ltd.: Cambridge, UK, 2011; pp. 321–345. [Google Scholar]
- Windhorst, H.-W.; Grabkowsky, B.; Wilke, A. Atlas of the Global Egg Industry; International Egg Commission: London, UK, 2015. [Google Scholar]
- Tukur, H.M. Egg production in Africa. In Improving the Safety of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 1, pp. 27–38. [Google Scholar]
- Evenepoel, P.; Claus, D.; Geypens, B.; Hiele, M.; Geboes, K.; Rutgeerts, P.; Ghoos, Y. Amount and fate of egg protein escaping assimilation in the small intestine of humans. Am. J. Physiol. 1999, 277, G935–G943. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, H.M.; Santos, V.V.; Medeiros, V.P.; Silva, K.H.; Dimenstein, R. Effect of thermal processing on retinol levels of free-range and caged hen eggs. Int. J. Food Sci. Nutr. 2006, 57, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Evenepoel, P.; Geypens, B.; Luypaerts, A.; Hiele, M.; Ghoos, Y.; Rutgeerts, P. Digestibility of cooked and raw egg protein in humans as assessed by stable isotope techniques. J. Nutr. 1998, 128, 1716–1722. [Google Scholar] [CrossRef]
- Stanciuc, N.; Cretu, A.A.; Banu, I.; Aprodu, I. Advances on the impact of thermal processing on structure and antigenicity of chicken ovomucoid. J. Sci. Food Agric. 2018, 98, 3119–3128. [Google Scholar] [CrossRef]
- Van der Plancken, I.; Van Remoortere, M.; Van Loey, A.; Hendrickx, M.E. Trypsin inhibition activity of heat-denatured ovomucoid: A kinetic study. Biotechnol. Prog. 2004, 20, 82–86. [Google Scholar] [CrossRef]
- Giansanti, F.; Massucci, M.T.; Giardi, M.F.; Nozza, F.; Pulsinelli, E.; Nicolini, C.; Botti, D.; Antonini, G. Antiviral activity of ovotransferrin derived peptides. Biochem. Biophys. Res. Commun. 2005, 331, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ledesma, B.; Hsieh, C.C. Chemopreventive role of food-derived proteins and peptides: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2358–2376. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Fan, H.B.; Wu, J.P. Egg White-Derived Antihypertensive Peptide IRW (Ile-Arg-Trp) Inhibits Angiotensin II-Stimulated Migration of Vascular Smooth Muscle Cells via Angiotensin Type I Receptor. J. Agric. Food Chem. 2018, 66, 5133–5138. [Google Scholar] [CrossRef] [PubMed]
- Dave, L.A.; Hayes, M.; Montoya, C.A.; Rutherfurd, S.M.; Moughan, P.J. Human gut endogenous proteins as a potential source of angiotensin-I-converting enzyme (ACE-I)-, renin inhibitory and antioxidant peptides. Peptides 2016, 76, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Seuss-Baum, I.; Nau, F.; Guérin-Dubiard, C. The nutritional quality of eggs. In Improving the Safety and Quality of Egg and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2 Egg Safety and Nutrional Quality; pp. 201–236. [Google Scholar]
- USDA National Nutrient Database for Standard Reference, Release 1; U.S. Department of Agriculture. Food Group: Dairy and Egg Products: Beltsville, MD, USA, 2018.
- Nys, Y.; Guyot, N. Egg formation and chemistry. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 1, pp. 83–132. [Google Scholar]
- Mann, K. The chicken egg white proteome. Proteomics 2007, 7, 3558–3568. [Google Scholar] [CrossRef] [PubMed]
- Mann, K. Proteomic analysis of the chicken egg vitelline membrane. Proteomics 2008, 8, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Mann, M. The chicken egg yolk plasma and granule proteomes. Proteomics 2008, 8, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.; Rehault-Godbert, S.; Nys, Y.; Mann, K.; Righetti, P.G. Use of high-throughput technology to identify new egg components. In Improving the Safety and Quality of Eggs and Egg Products, Vol 1: Egg Chemistry, Production and Consumption; Nys, Y., Bain, M., VanImmerseel, F., Eds.; Woodhead Publ Ltd.: Cambridge, UK, 2011; Volume 1, pp. 133–150. [Google Scholar]
- Gautron, J.; Nau, F.; Mann, K.; Guerin-Dubiard, C.; Rehault, S.; Hincke, M.T.; Nys, Y. Molecular approaches for the identification of novel egg components. Worlds Poult. Sci. J. 2007, 63, 82–90. [Google Scholar] [CrossRef]
- Farinazzo, A.; Restuccia, U.; Bachi, A.; Guerrier, L.; Fortis, F.; Boschetti, E.; Fasoli, E.; Citterio, A.; Righetti, P.G. Chicken egg yolk cytoplasmic proteome, mined via combinatorial peptide ligand libraries. J. Chromatogr. A 2009, 1216, 1241–1252. [Google Scholar] [CrossRef]
- Guerin-Dubiard, C.; Pasco, M.; Molle, D.; Desert, C.; Croguennec, T.; Nau, F. Proteomic analysis of hen egg white. J. Agric. Food Chem. 2006, 54, 3901–3910. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Macek, B.; Olsen, J.V. Proteomic analysis of the acid-soluble organic matrix of the chicken calcified eggshell layer. Proteomics 2006, 6, 3801–3810. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Olsen, J.V.; Macek, B.; Gnad, F.; Mann, M. Phosphoproteins of the chicken eggshell calcified layer. Proteomics 2007, 7, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Anton, M. Egg yolk: Structures, functionalities and processes. J. Sci. Food Agric. 2013, 93, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Anton, M.; Gandemer, G. Composition, Solubility and Emulsifying Properties of Granules and Plasma of Egg Yolk. J. Food Sci. 1997, 62, 484–487. [Google Scholar] [CrossRef]
- Rehault-Godbert, S.; Guyot, N. Vitellogenesis and Yolk Proteins, Birds. In Encyclopedia of Reproduction; Skinner, M., Ed.; Elsevier: Amsterdam, The Netherlands; London, UK, 2018; Volume 6. [Google Scholar]
- Bellairs, R.; Harkness, M.; Harkness, R.D. The vitelline membrane of the hen’s egg: A chemical and electron microcopical study. J. Ultrastruct. Res. 1963, 8, 339–359. [Google Scholar] [CrossRef]
- Back, J.F.; Bain, J.M.; Vadehra, D.V.; Burley, R.W. Proteins of the outer layer of the vitelline membrane of hen’s eggs. Biochim. Et Biophys. Acta 1982, 705, 12–19. [Google Scholar] [CrossRef]
- Hawthorne, J.R. The action of egg white lysozyme on ovomucoid and ovomucin. Biochim. Et Biophys. Acta 1950, 6, 28–35. [Google Scholar] [CrossRef]
- Saxena, I.; Tayyab, S. Protein proteinase inhibitors from avian egg whites. Cell. Mol. Life Sci. CMLS 1997, 53, 13–23. [Google Scholar] [CrossRef]
- Trziszka, T.; Smolinska, T. Chemical characterization of the vitelline membrane of Hens eggs. Food Chem. 1982, 8, 61–70. [Google Scholar] [CrossRef]
- Shinn, S.E.; Liyanage, R.; Lay, J.O., Jr.; Proctor, A. Isolation and Characterization of Chicken Yolk Vitelline Membrane Lipids Using Eggs Enriched with Conjugated Linoleic Acid. Lipids 2016, 51, 769–779. [Google Scholar] [CrossRef]
- Hayes, K.C.; Khosla, P. Dietary fatty acid thresholds and cholesterolemia. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1992, 6, 2600–2607. [Google Scholar] [CrossRef]
- Pronczuk, A.; Khosla, P.; Hayes, K.C. Dietary myristic, palmitic, and linoleic acids modulate cholesterolemia in gerbils. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1994, 8, 1191–1200. [Google Scholar] [CrossRef]
- Chatterjee, I.B. Evolution and the biosynthesis of ascorbic acid. Science 1973, 182, 1271–1272. [Google Scholar] [CrossRef]
- Patterson, K.Y.; Bhagwat, S.A.; Williams, J.R.; Howe, J.C.; Holden, J.M. USDA Database for the Choline Content of Common Foods, Release 2; Nutrient Data Laboratory: Beltsville, MD, USA, 2008. [Google Scholar]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef]
- Wallace, T.C.; Fulgoni, V.L. Usual Choline Intakes Are Associated with Egg and Protein Food Consumption in the United States. Nutrients 2017, 9, 839. [Google Scholar] [CrossRef]
- Leermakers, E.T.; Moreira, E.M.; Kiefte-de Jong, J.C.; Darweesh, S.K.; Visser, T.; Voortman, T.; Bautista, P.K.; Chowdhury, R.; Gorman, D.; Bramer, W.M.; et al. Effects of choline on health across the life course: A systematic review. Nutr. Rev. 2015, 73, 500–522. [Google Scholar] [CrossRef]
- Oyen, J.; Gjesdal, C.G.; Karlsson, T.; Svingen, G.F.; Tell, G.S.; Strand, E.; Drevon, C.A.; Vinknes, K.J.; Meyer, K.; Ueland, P.M.; et al. Dietary Choline Intake Is Directly Associated with Bone Mineral Density in the Hordaland Health Study. J. Nutr. 2017, 147, 572–578. [Google Scholar] [CrossRef]
- Staggs, C.G.; Sealey, W.M.; McCabe, B.J.; Teague, A.M.; Mock, D.M. Determination of the biotin content of select foods using accurate and sensitive HPLC/avidin binding. J. Food Compos. Anal. Off. Publ. U. N. Univ. Int. Netw. Food Data Syst. 2004, 17, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Um, P.; Dickerman, B.A.; Liu, J. Zinc, Magnesium, Selenium and Depression: A Review of the Evidence, Potential Mechanisms and Implications. Nutrients 2018, 10, 584. [Google Scholar] [CrossRef]
- Livnah, O.; Bayer, E.A.; Wilchek, M.; Sussman, J.L. Three-dimensional structures of avidin and the avidin-biotin complex. Proc. Natl. Acad. Sci. USA 1993, 90, 5076–5080. [Google Scholar] [CrossRef]
- Chang, C.; Lahti, T.; Tanaka, T.; Nickerson, M.T. Egg proteins: Fractionation, bioactive peptides and allergenicity. J. Sci. Food Agric. 2018, 98, 5547–5558. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.; Lee, H.Y.; Ahn, D.U. Egg white proteins and their potential use in food processing or as nutraceutical and pharmaceutical agents—A review. Poult. Sci. 2013, 92, 3292–3299. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.J. Bioactive Egg Components and Inflammation. Nutrients 2015, 7, 7889–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giansanti, F.; Leboffe, L.; Angelucci, F.; Antonini, G. The Nutraceutical Properties of Ovotransferrin and Its Potential Utilization as a Functional Food. Nutrients 2015, 7, 9105–9115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef]
- Arena, S.; Scaloni, A. An Extensive Description of the Peptidomic Repertoire of the Hen Egg Yolk Plasma. J. Agric. Food Chem. 2018, 66, 3239–3255. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Mendez, P.; Reyes, W.; Romero, H.; Pinto, A.; Carrillo, W. Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments. Sci. Pharm. 2018, 86, 48. [Google Scholar] [CrossRef]
- Rehault-Godbert, S.; Herve-Grepinet, V.; Gautron, J.; Cabau, C.; Nys, Y.; Hincke, M. Molecules involved in chemical defence of the chicken egg. In Improving the Safety and Quality of Eggs and Egg Products, Vol 1: Egg Chemistry, Production and Consumption; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publ Ltd.: Cambridge, UK, 2011; Volume 1, pp. 183–208. [Google Scholar]
- Da Silva, M.; Dombre, C.; Brionne, A.; Monget, P.; Chesse, M.; De Pauw, M.; Mills, M.; Combes-Soia, L.; Labas, V.; Guyot, N.; et al. The unique features of proteins depicting the chicken amniotic fluid. Mol. Cell. Proteom. MCP 2018. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Ma, F.P.; Lauriau, S. Antimicrobial peptides released by enzymatic hydrolysis of hen egg white lysozyme. J. Agric. Food Chem. 2004, 52, 1088–1094. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Thomas, U.; Pellegrini, A. A helix-loop-helix peptide at the upper lip of the active site cleft of lysozyme confers potent antimicrobial activity with membrane permeabilization action. J. Biol. Chem. 2001, 276, 43767–43774. [Google Scholar] [CrossRef]
- Pellegrini, A.; Thomas, U.; Wild, P.; Schraner, E.; von Fellenberg, R. Effect of lysozyme or modified lysozyme fragments on DNA and RNA synthesis and membrane permeability of Escherichia coli. Microbiol. Res. 2000, 155, 69–77. [Google Scholar] [CrossRef]
- Pellegrini, A.; Thomas, U.; Bramaz, N.; Klauser, S.; von Fellenberg, R. Identification and isolation of a bactericidal domain in chicken egg white lysozyme. J. Appl. Microbiol. 1997, 82, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Omana, D.A.; Wang, J.P.; Wu, J.P. Ovomucin—A glycoprotein with promising potential. Trends Food Sci. Technol. 2010, 21, 455–463. [Google Scholar] [CrossRef]
- Blankenvoorde, M.F.J.; van’t Hof, W.; Walgreen-Weterings, E.; van Steenbergen, T.J.M.; Brand, H.S.; Veerman, E.C.I.; Amerongen, A.V.N. Cystatin and cystatin-derived peptides have antibacterial activity against the pathogen Porphyromonas gingivalis. Biol. Chem. 1998, 379, 1371–1375. [Google Scholar]
- Guyot, N.; Labas, V.; Harichaux, G.; Chesse, M.; Poirier, J.C.; Nys, Y.; Rehault-Godbert, S. Proteomic analysis of egg white heparin-binding proteins: Towards the identification of natural antibacterial molecules. Sci. Rep. 2016, 6, 27974. [Google Scholar] [CrossRef]
- Herve-Grepinet, V.; Rehault-Godbert, S.; Labas, V.; Magallon, T.; Derache, C.; Lavergne, M.; Gautron, J.; Lalmanach, A.C.; Nys, Y. Purification and Characterization of Avian beta-Defensin 11, an Antimicrobial Peptide of the Hen Egg. Antimicrob. Agents Chemother. 2010, 54, 4401–4408. [Google Scholar] [CrossRef]
- Korpela, J.; Salonen, E.M.; Kuusela, P.; Sarvas, M.; Vaheri, A. Binding of avidin to bacteria and to the outer-membrane porin of Escherichia coli. FEMS Microbiol. Lett. 1984, 22, 3–10. [Google Scholar] [CrossRef]
- Krkavcova, E.; Kreisinger, J.; Hyankova, L.; Hyrsl, P.; Javurkova, V. The hidden function of egg white antimicrobials: Egg weight-dependent effects of avidin on avian embryo survival and hatchling phenotype. Biol. Open 2018, 7, 9. [Google Scholar] [CrossRef]
- Kolaczkowska, A.; Kolaczkowski, M.; Sokolowska, A.; Miecznikowska, H.; Kubiak, A.; Rolka, K.; Polanowski, A. The antifungal properties of chicken egg cystatin against Candida yeast isolates showing different levels of azole resistance. Mycoses 2010, 53, 314–320. [Google Scholar] [CrossRef]
- Wesierska, E.; Saleh, Y.; Trziszka, T.; Kopec, W.; Siewinski, M.; Korzekwa, K. Antimicrobial activity of chicken egg white cystatin. World J. Microbiol. Biotechnol. 2005, 21, 59–64. [Google Scholar] [CrossRef]
- Bjorck, L. Proteinase inhibition, immunoglobulin-binding proteins and a novel antimicrobial principle. Mol. Microbiol. 1990, 4, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Korant, B.D.; Brzin, J.; Turk, V. Cystatin, a protein inhibitor of cysteine proteases alters viral protein cleavages in infected human cells. Biochem. Biophys. Res. Commun. 1985, 127, 1072–1076. [Google Scholar] [CrossRef]
- Ebina, T.; Tsukada, K. Protease inhibitors prevent the development of human rotavirus-induced diarrhea in suckling mice. Microbiol. Immunol. 1991, 35, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.P.; dos Reis, F.C.; Serveau, C.; Lalmanach, G.; Juliano, L.; Menard, R.; Vernet, T.; Thomas, D.Y.; Storer, A.C.; Scharfstein, J. Cysteine protease isoforms from Trypanosoma cruzi, cruzipain 2 and cruzain, present different substrate preference and susceptibility to inhibitors. Mol. Biochem. Parasitol. 2001, 114, 41–52. [Google Scholar] [CrossRef]
- Yu, L.T.; Xiao, Y.P.; Li, J.J.; Ran, J.S.; Yin, L.Q.; Liu, Y.P.; Zhang, L. Molecular characterization of a novel ovodefensin gene in chickens. Gene 2018, 678, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Whenham, N.; Lu, T.C.; Maidin, M.B.M.; Wilson, P.W.; Bain, M.M.; Stevenson, M.L.; Stevens, M.P.; Bedford, M.R.; Dunn, I.C. Ovodefensins, an Oviduct-Specific Antimicrobial Gene Family, Have Evolved in Birds and Reptiles to Protect the Egg by Both Sequence and Intra-Six-Cysteine Sequence Motif Spacing. Biol. Reprod. 2015, 92, 13. [Google Scholar] [CrossRef]
- Herve, V.; Meudal, H.; Labas, V.; Rehault-Godbert, S.; Gautron, J.; Berges, M.; Guyot, N.; Delmas, A.F.; Nys, Y.; Landon, C. Three-dimensional NMR Structure of Hen Egg Gallin (Chicken Ovodefensin) Reveals a New Variation of the beta-Defensin Fold. J. Biol. Chem. 2014, 289, 7211–7220. [Google Scholar] [CrossRef]
- Gong, D.Q.; Wilson, P.W.; Bain, M.M.; McDade, K.; Kalina, J.; Herve-Grepinet, V.; Nys, Y.; Dunn, I.C. Gallin; an antimicrobial peptide member of a new avian defensin family, the ovodefensins, has been subject to recent gene duplication. BMC Immunol. 2010, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Kovacs-Nolan, J.; Mine, Y. Egg Yolk Antibodies for Passive Immunity. In Annual Review of Food Science and Technology, Volume 3; Doyle, M.P., Klaenhammer, T.R., Eds.; Annual Reviews: Palo Alto, CA, USA, 2012; Volume 3, pp. 163–182. [Google Scholar]
- Wellman-Labadie, O.; Picman, J.; Hincke, M.T. Comparative antibacterial activity of avian egg white protein extracts. Br. Poult. Sci. 2008, 49, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Samaranayake, Y.H.; Samaranayake, L.P.; Pow, E.H.N.; Beena, V.T.; Yeung, K.W.S. Antifungal effects of lysozyme and lactoferrin against genetically similar, sequential Candida albicans isolates from a human immunodeficiency virus-infected southern Chinese cohort. J. Clin. Microbiol. 2001, 39, 3296–3302. [Google Scholar] [CrossRef]
- Rehault-Godbert, S.; Labas, V.; Helloin, E.; Herve-Grepinet, V.; Slugocki, C.; Berges, M.; Bourin, M.C.; Brionne, A.; Poirier, J.C.; Gautron, J.; et al. Ovalbumin-related protein X is a heparin-binding ov-serpin exhibiting antimicrobial activities. J. Biol. Chem. 2013, 288, 17285–17295. [Google Scholar] [CrossRef]
- Maehashi, K.; Ueda, M.; Matano, M.; Takeuchi, J.; Uchino, M.; Kashiwagi, Y.; Watanabe, T. Biochemical and functional characterization of transiently expressed in neural precursor (TENP) protein in emu egg white. J. Agric. Food Chem. 2014, 62, 5156–5162. [Google Scholar] [CrossRef]
- Whenham, N.; Wilson, P.W.; Bain, M.M.; Stevenson, L.; Dunn, I.C. Comparative biology and expression of TENP, an egg protein related to the bacterial permeability-increasing family of proteins. Gene 2014, 538, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Malicka-Blaszkiewicz, M.; Filipczak, N.; Golab, K.; Juszczynska, K.; Sebzda, T.; Gburek, J. Ovocystatin affects actin cytoskeleton organization and induces proapoptotic activity. Acta Biochim. Pol. 2014, 61, 753–758. [Google Scholar] [PubMed]
- Bourin, M.; Gautron, J.; Berges, M.; Attucci, S.; Le Blay, G.; Labas, V.; Nys, Y.; Rehault-Godbert, S. Antimicrobial Potential of Egg Yolk Ovoinhibitor, a Multidomain Kazal-like Inhibitor of Chicken Egg. J. Agric. Food Chem. 2011, 59, 12368–12374. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Lee, J.H.; Lee, Y.J.; Paik, J.Y.; Ahn, D.U.; Paik, H.D. Antioxidant, Antimicrobial, and Cytotoxic Activities of Ovotransferrin from Egg White. Korean J. Food Sci. 2012, 32, 612–617. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.P.; Acero-Lopez, A. Ovotransferrin: Structure, bioactivities, and preparation. Food Res. Int. 2012, 46, 480–487. [Google Scholar] [CrossRef]
- Giansanti, F.; Giardi, M.F.; Massucci, M.T.; Botti, D.; Antonini, G. Ovotransferrin expression and release by chicken cell lines infected with Marek’s disease virus. Biochem. Cell Biol. 2007, 85, 150–155. [Google Scholar] [CrossRef]
- Khan, M.A.S.; Nakamura, S.; Ogawa, M.; Akita, E.; Azakami, H.; Kato, A. Bactericidal action of egg yolk phosvitin against Escherichia coli under thermal stress. J. Agric. Food Chem. 2000, 48, 1503–1506. [Google Scholar] [CrossRef]
- Svensson, S.L.; Pasupuleti, M.; Walse, B.; Malmsten, M.; Morgelin, M.; Sjogren, C.; Olin, A.I.; Collin, M.; Schmidtchen, A.; Palmer, R.; et al. Midkine and pleiotrophin have bactericidal properties: Preserved antibacterial activity in a family of heparin-binding growth factors during evolution. J. Biol. Chem. 2010, 285, 16105–16115. [Google Scholar] [CrossRef]
- Sun, S.; Niu, H.; Yang, T.; Lin, Q.; Luo, F.; Ma, M. Antioxidant and anti-fatigue activities of egg white peptides prepared by pepsin digestion. J. Sci. Food Agric. 2014, 94, 3195–3200. [Google Scholar] [CrossRef] [PubMed]
- Abeyrathne, E.; Huang, X.; Ahn, D.U. Antioxidant, angiotensin-converting enzyme inhibitory activity and other functional properties of egg white proteins and their derived peptides—A review. Poult. Sci. 2018, 97, 1462–1468. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Bandara, N.; Wu, J.P. Purification and characterization of antioxidant peptides from enzymatically hydrolyzed chicken egg white. Food Chem. 2015, 188, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Nimalaratne, C.; Wu, J. Hen Egg as an Antioxidant Food Commodity: A Review. Nutrients 2015, 7, 8274–8293. [Google Scholar] [CrossRef]
- Yi, J.E.; Zhao, J.; Wu, J.P. Egg ovotransferrin derived IRW exerts protective effect against H2O2-induced oxidative stress in Caco-2 cells. J. Funct. Foods 2017, 39, 160–167. [Google Scholar] [CrossRef]
- Shen, S.W.; Chahal, B.; Majumder, K.; You, S.J.; Wu, J.P. Identification of Novel Antioxidative Peptides Derived from a Thermolytic Hydrolysate of Ovotransferrin by LC-MS/MS. J. Agric. Food Chem. 2010, 58, 7664–7672. [Google Scholar] [CrossRef]
- Kim, J.; Moon, S.H.; Ahn, D.U.; Paik, H.D.; Park, E. Antioxidant effects of ovotransferrin and its hydrolysates. Poult. Sci. 2012, 91, 2747–2754. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chi, Y.J.; Zhao, M.Y.; Lv, L. Purification and identification of antioxidant peptides from egg white protein hydrolysate. Amino Acids 2012, 43, 457–466. [Google Scholar] [CrossRef]
- Abeyrathne, E.; Lee, H.Y.; Jo, C.; Suh, J.W.; Ahn, D.U. Enzymatic hydrolysis of ovomucoid and the functional properties of its hydrolysates. Poult. Sci. 2015, 94, 2280–2287. [Google Scholar] [CrossRef] [Green Version]
- Chang, O.K.; Ha, G.E.; Han, G.S.; Seol, K.H.; Kim, H.W.; Jeong, S.G.; Oh, M.H.; Park, B.Y.; Ham, J.S. Novel Antioxidant Peptide Derived from the Ultrafiltrate of Ovomucin Hydrolysate. J. Agric. Food Chem. 2013, 61, 7294–7300. [Google Scholar] [CrossRef]
- Yousr, M.; Howell, N. Antioxidant and ACE Inhibitory Bioactive Peptides Purified from Egg Yolk Proteins. Int. J. Mol. Sci. 2015, 16, 29161–29178. [Google Scholar] [CrossRef] [Green Version]
- Young, D.; Fan, M.Z.; Mine, Y. Egg Yolk Peptides Up-regulate Glutathione Synthesis and Antioxidant Enzyme Activities in a Porcine Model of Intestinal Oxidative Stress. J. Agric. Food Chem. 2010, 58, 7624–7633. [Google Scholar] [CrossRef]
- Sava, G. Reduction of B16 melanoma metastases by oral administration of egg-white lysozyme. Cancer Chemother. Pharmacol. 1989, 25, 221–222. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Kiyono, T. Novel Anticancer Activity of the Autocleaved Ovotransferrin against Human Colon and Breast Cancer Cells. J. Agric. Food Chem. 2009, 57, 11383–11390. [Google Scholar] [CrossRef]
- Clemente, A.; Arques Mdel, C. Bowman-Birk inhibitors from legumes as colorectal chemopreventive agents. World J. Gastroenterol. 2014, 20, 10305–10315. [Google Scholar] [CrossRef]
- Lee, M.; Kovacs-Nolan, J.; Yang, C.; Archbold, T.; Fan, M.Z.; Mine, Y. Hen egg lysozyme attenuates inflammation and modulates local gene expression in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Agric. Food Chem. 2009, 57, 2233–2240. [Google Scholar] [CrossRef]
- Sorrelle, N.; Dominguez, A.T.A.; Brekken, R.A. From top to bottom: Midkine and pleiotrophin as emerging players in immune regulation. J. Leukoc. Biol. 2017, 102, 277–286. [Google Scholar] [CrossRef]
- Achour, A.; M’Bika, J.P.; Baudouin, F.; Caruelle, D.; Courty, J. Pleiotrophin induces expression of inflammatory cytokines in peripheral blood mononuclear cells. Biochimie 2008, 90, 1791–1795. [Google Scholar] [CrossRef]
- Liu, L.; Xu, M.S.; Tu, Y.G.; Du, H.Y.; Zhou, Y.L.; Zhu, G.X. Immunomodulatory effect of protease hydrolysates from ovotransferrin. Food Funct. 2017, 8, 1452–1459. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Yao, Y.; Xu, M.S.; Du, H.Y.; Zhang, M.Y.; Tu, Y.G. Anti-inflammatory activity of di-peptides derived from ovotransferrin by CrossMark simulated peptide-cut in TNF-alpha-induced Caco-2 cells. J. Funct. Foods 2017, 37, 424–432. [Google Scholar] [CrossRef]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Whelton, P.K.; He, J. Worldwide prevalence of hypertension: A systematic review. J. Hypertens. 2004, 22, 11–19. [Google Scholar] [CrossRef]
- Yoshii, H.; Tachi, N.; Ohba, R.; Sakamura, O.; Takeyama, H.; Itani, T. Antihypertensive effect of ACE inhibitory oligopeptides from chicken egg yolks. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2001, 128, 27–33. [Google Scholar] [CrossRef]
- Moon, S.H.; Lee, J.H.; Kim, J.H.; Paik, H.D.; Ahn, D.U. In vitro cytotoxic and ACE-inhibitory activities of promod 278P hydrolysate of ovotransferrin from chicken egg white. Poult. Sci. 2017, 96, 1982–1987. [Google Scholar] [CrossRef]
- Majumder, K.; Wu, J.P. Purification and characterisation of angiotensin I converting enzyme (ACE) inhibitory peptides derived from enzymatic hydrolysate of ovotransferrin. Food Chem. 2011, 126, 1614–1619. [Google Scholar] [CrossRef]
- Liao, W.; Chakrabarti, S.; Davidge, S.T.; Wu, J.P. Modulatory Effects of Egg White Ovotransferrin-Derived Tripeptide IRW (Ile-Arg-Trp) on Vascular Smooth Muscle Cells against Angiotensin II Stimulation. J. Agric. Food Chem. 2016, 64, 7342–7347. [Google Scholar] [CrossRef]
- Majumder, K.; Chakrabarti, S.; Morton, J.S.; Panahi, S.; Kaufman, S.; Davidge, S.T.; Wu, J.P. Egg-derived ACE-inhibitory peptides IQW and LKP reduce blood pressure in spontaneously hypertensive rats. J. Funct. Foods 2015, 13, 50–60. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, H.M.; Peng, H.H.; Wu, X.S.; Fang, J. The Utility of Ovotransferrin and Ovotransferrin-Derived Peptides as Possible Candidates in the Clinical Treatment of Cardiovascular Diseases. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef]
- Dunn, I.C. Breeding strategies to improve the egg’s natural defence. Worlds Poult. Sci. J. 2004, 60, 458–468. [Google Scholar] [CrossRef]
- Dunn, I.C. Poultry breeding for egg quality. In Improving the Safety and Quality of Eggs and Egg Products, Volume 1: Egg Chemistry, Production and Consumption; Nys, Y., Bain, M., VanImmerseel, F., Eds.; Woodhead Publ Ltd.: Cambridge, UK, 2011; Volume 1, pp. 245–260. [Google Scholar]
- Bilkova, B.; Swiderska, Z.; Zita, L.; Laloe, D.; Charles, M.; Benes, V.; Stopka, P.; Vinkler, M. Domestic Fowl Breed Variation in Egg White Protein Expression: Application of Proteomics and Transcriptomics. J. Agric. Food Chem. 2018, 66, 11854–11863. [Google Scholar] [CrossRef]
- Hocking, P.M.; Bain, M.; Channing, C.E.; Fleming, R.; Wilson, S. Genetic variation for egg production, egg quality and bone strength in selected and traditional breeds of laying fowl. Br. Poult. Sci. 2003, 44, 365–373. [Google Scholar] [CrossRef]
- Sellier, N.; Vidal, M.L.; Baron, F.; Michel, J.; Gautron, J.; Protais, M.; Beaumont, C.; Gautier, M.; Nys, Y. Estimations of repeatability and heritability of egg albumen antimicrobial activity and of lysozyme and ovotransferrin concentrations. Br. Poult. Sci. 2007, 48, 559–566. [Google Scholar] [CrossRef]
- Anderson, K.E. Comparison of fatty acid, cholesterol, vitamin A and E composition, and trans fats in eggs from brown and white egg strains that were molted or nonmolted. Poult. Sci. 2013, 92, 3259–3265. [Google Scholar] [CrossRef] [Green Version]
- Silversides, F.G.; Budgell, K. The relationships among measures of egg albumen height, pH, and whipping volume. Poult. Sci. 2004, 83, 1619–1623. [Google Scholar] [CrossRef]
- Bouvarel, I.; Nys, Y.; Lescoat, P. Hen nutrition for sustained egg quality. In Improving the Safety and Quality of Eggs and Egg Products, Vol 1: Egg Chemistry, Production and Consumption; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publ Ltd.: Cambridge, UK, 2011; pp. 261–299. [Google Scholar]
- Nys, Y.; Le Roy, N. Calcium Homeostasis and Eggshell Biomineralization in Female Chicken. In Vitamin D, 4th ed.; Feldman, D., Ed.; Academic Press: London, UK, 2018; Volume 1, pp. 361–382. [Google Scholar]
- Grobas, S.; Mendez, J.; De Blas, C.; Mateos, G.G. Influence of dietary energy, supplemental fat and linoleic acid concentration on performance of laying hens at two ages. Br. Poult. Sci. 1999, 40, 681–687. [Google Scholar] [CrossRef]
- Bregendahl, K.; Roberts, S.A.; Kerr, B.; Hoehler, D. Ideal ratios of isoleucine, methionine, methionine plus cystine, threonine, tryptophan, and valine relative to lysine for white leghorn-type laying hens of twenty-eight to thirty-four weeks of age. Poult. Sci. 2008, 87, 744–758. [Google Scholar] [CrossRef]
- Recoules, E.; Sabboh-Jourdan, H.; Narcy, A.; Lessire, M.; Harichaux, G.; Labas, V.; Duclos, M.J.; Rehault-Godbert, S. Exploring the in vivo digestion of plant proteins in broiler chickens. Poult. Sci. 2017, 96, 1735–1747. [Google Scholar] [CrossRef] [Green Version]
- Nain, S.; Renema, R.A.; Korver, D.R.; Zuidhof, M.J. Characterization of the n-3 polyunsaturated fatty acid enrichment in laying hens fed an extruded flax enrichment source. Poult. Sci. 2012, 91, 1720–1732. [Google Scholar] [CrossRef] [Green Version]
- Coorey, R.; Novinda, A.; Williams, H.; Jayasena, V. Omega-3 fatty acid profile of eggs from laying hens fed diets supplemented with chia, fish oil, and flaxseed. J. Food Sci. 2015, 80, S180–S187. [Google Scholar] [CrossRef]
- Sirri, F.; Meluzzi, A. Modifying egg lipids for human health. In Improving the Safety and Quality of Eggs and Egg Products; Van Immerseel, F., Nys, Y., Bain, M., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 272–288. [Google Scholar]
- Baeza, E.; Chartrin, P.; Lessire, M.; Meteau, K.; Chesneau, G.; Guillevic, M.; Mourot, J. Is it possible to increase the n-3 fatty acid content of eggs without affecting their technological and/or sensorial quality and the laying performance of hens? Br. Poult. Sci. 2015, 56, 748–754. [Google Scholar] [CrossRef]
- Grcevic, M.; Kralik, Z.; Kralik, G.; Galovic, O. Effects of dietary marigold extract on lutein content, yolk color and fatty acid profile of omega-3 eggs. J. Sci. Food Agric. 2019, 99, 2292–2299. [Google Scholar] [CrossRef]
- Park, J.H.; Upadhaya, S.D.; Kim, I.H. Effect of dietary marine microalgae (schizochytrium) powder on egg production, blood lipid profiles, egg quality, and Fatty Acid composition of egg yolk in layers. Asian-Australas. J. Anim. Sci. 2015, 28, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.G.; Sieo, C.C.; Kalavathy, R.; Saad, W.Z.; Yong, S.T.; Wong, H.K.; Ho, Y.W. Chemical Compositions of Egg Yolks and Egg Quality of Laying Hens Fed Prebiotic, Probiotic, and Synbiotic Diets. J. Food Sci. 2015, 80, C1686–C1695. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; Barroeta, A.C. Egg enrichment with vitamins and trace minerals. In Improving the Safety and Quality of Eggs and Egg Products; Van Immerseel, F., Nys, Y., Bain, M., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 289–320. [Google Scholar]
- Nys, Y.; Schlegel, P.; Durosoy, S.; Jondreville, C.; Narcy, A. Adapting trace mineral nutrition of birds for optimising the environment and poultry product quality. Worlds Poult. Sci. J. 2018, 74, 225–238. [Google Scholar] [CrossRef]
- Ranard, K.M.; Jeon, S.; Mohn, E.S.; Griffiths, J.C.; Johnson, E.J.; Erdman, J.W., Jr. Dietary guidance for lutein: Consideration for intake recommendations is scientifically supported. Eur. J. Nutr. 2017, 56, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Pignoli, G.; Rodriguez-Estrada, M.T.; Mandrioli, M.; Barbanti, L.; Rizzi, L.; Lercker, G. Effects of different rearing and feeding systems on lipid oxidation and antioxidant capacity of freeze-dried egg yolks. J. Agric. Food Chem. 2009, 57, 11517–11527. [Google Scholar] [CrossRef] [PubMed]
- Samiullah; Roberts, J.R.; Chousalkar, K.K. Effect of production system and flock age on egg quality and total bacterial load in commercial laying hens. J. Appl. Poult. Res. 2014, 23, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Bedrani, L.; Helloin, E.; Guyot, N.; Rehault-Godbert, S.; Nys, Y. Passive maternal exposure to environmental microbes selectively modulates the innate defences of chicken egg white by increasing some of its antibacterial activities. BMC Microbiol. 2013, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Travel, A.; Nys, Y.; Bain, M. Effect of hens age, moult, laying environment and egg storage on egg quality. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 1, pp. 300–329. [Google Scholar]
- Rossi, M.; Pompei, C. Changes in some egg components and analytical values due to hen age. Poult. Sci. 1995, 74, 152–160. [Google Scholar] [CrossRef]
- Ngoka, D.A.; Froning, G.W.; Babji, A.S. Effect of temperature on egg-yolk characteristics of eggs from young and old laying hens. Poult. Sci. 1983, 62, 718–720. [Google Scholar] [CrossRef]
- Cherian, G. Egg quality and yolk polyunsaturated fatty acid status in relation to broiler breeder hen age and dietary n-3 oils. Poult. Sci. 2008, 87, 1131–1137. [Google Scholar] [CrossRef]
- Liu, X.T.; Lin, X.; Mi, Y.L.; Zeng, W.D.; Zhang, C.Q. Age-related changes of yolk precursor formation in the liver of laying hens. J. Zhejiang Univ. Sci. B 2018, 19, 390–399. [Google Scholar] [CrossRef]
- Roberts, J.R.; Souillard, R.; Bertin, J. Avian diseases which affect egg production and quality. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 1, pp. 376–393. [Google Scholar]
- Heerkens, J.L.; Delezie, E.; Kempen, I.; Zoons, J.; Ampe, B.; Rodenburg, T.B.; Tuyttens, F.A. Specific characteristics of the aviary housing system affect plumage condition, mortality and production in laying hens. Poult. Sci. 2015, 94, 2008–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaab, H.; Bain, M.M.; Bartley, K.; Turnbull, F.; Wright, H.W.; Nisbet, A.J.; Birchmore, R.; Eckersall, P.D. Serum and acute phase protein changes in laying hens, infested with poultry red mite. Poult. Sci. 2018. [Google Scholar] [CrossRef]
- Sigognault Flochlay, A.; Thomas, E.; Sparagano, O. Poultry red mite (Dermanyssus gallinae) infestation: A broad impact parasitological disease that still remains a significant challenge for the egg-laying industry in Europe. Parasites Vectors 2017, 10, 357. [Google Scholar] [CrossRef]
- Tomley, F.M.; Sparagano, O. Spotlight on avian pathology: Red mite, a serious emergent problem in layer hens. Avian Pathol. J. 2018, 47, 533–535. [Google Scholar] [CrossRef]
- De Vylder, J.; Raspoet, R.; Dewulf, J.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Salmonella Enteritidis is superior in egg white survival compared with other Salmonella serotypes. Poult. Sci. 2013, 92, 842–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lublin, A.; Sela, S. The Impact of Temperature During the Storage of Table Eggs on the Viability of Salmonella enterica Serovars Enteritidis and Virchow in the Eggs. Poult. Sci. 2008, 87, 2208–2214. [Google Scholar] [CrossRef] [Green Version]
- Chemaly, M.; Salvat, G. Foodborne disease associated with eggs: Microbial hazards and Salmonella Enteritidis risk assessment. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Immersel, F.V., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2. [Google Scholar]
- European Centre for Disease Prevention and Control. Salmonellosis—Annual Epidemiological Report 2016 [2014 Data]; ECDC: Solna, Sweden, 2016.
- Davies, R. Detection and monotoring of Salmonella in laying hens flocks. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Immersel, F.V., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 83–106. [Google Scholar]
- Gast, R.K. Pre-harvest measures to control Salmonella in laying hens. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 120–145. [Google Scholar]
- Ducatelle, R.; Van Immerseel, F. Management and sanitation procedures to control Salmonella in laying hens flocks. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Immersel, F.V., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 146–162. [Google Scholar]
- Messens, W.; Gittins, J.; Leleu, S.; Sparks, N. Egg decontamination by washing. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 163–180. [Google Scholar]
- Berardinelli, A.; Ragni, L.; Giunchi, A.; Gradari, P.; Guarnieri, A. Physical-mechanical modifications of eggs for food-processing during storage. Poult. Sci. 2008, 87, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Guyot, N.; Rehault-Godbert, S.; Nys, Y.; Baron, F. Understanding the natural antibacterial defences of egg white and their regulation. In Achieving Sustainable Production of Eggs; Roberts, J., Ed.; Burleigh Dodds Science Publishin Limited: Cambridge, UK, 2016; Volume 1, pp. 161–193. [Google Scholar]
- Rehault-Godbert, S.; Baron, F.; Mignon-Grasteau, S.; Labas, V.; Gautier, M.; Hincke, M.T.; Nys, Y. Effect of Temperature and Time of Storage on Protein Stability and Anti-Salmonella Activity of Egg White. J. Food Prot. 2010, 73, 1604–1612. [Google Scholar] [CrossRef]
- Samli, H.E.; Agma, A.; Senkoylu, N. Effects of storage time and temperature on egg quality in old laying hens. J. Appl. Poult. Res. 2005, 14, 548–553. [Google Scholar] [CrossRef]
- Garcia, F.J.; Pons, A.; Alemany, M.; Palou, A. Permeability of chicken egg vitelline membrane to glucose, carbohydrate gradients between albumen and yolk. Comp. Biochem. Physiology. Bcomp. Biochem. 1983, 75, 137–140. [Google Scholar] [CrossRef]
- Rehault-Godbert, S.; Mann, K.; Bourin, M.; Brionne, A.; Nys, Y. Effect of embryonic development on the chicken egg yolk plasma proteome after 12 days of incubation. J. Agric. Food Chem. 2014, 62, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Qiu, N.; Liu, Y.P.; Ma, M.H. Comparative proteome analysis of egg yolk plasma proteins during storage. J. Sci. Food Agric. 2017, 97, 2392–2400. [Google Scholar] [CrossRef]
- Qiu, N.; Ma, M.; Zhao, L.; Liu, W.; Li, Y.; Mine, Y. Comparative proteomic analysis of egg white proteins under various storage temperatures. J. Agric. Food Chem. 2012, 60, 7746–7753. [Google Scholar] [CrossRef] [PubMed]
- Bush, L.; White, H.B., 3rd. Avidin traps biotin diffusing out of chicken egg yolk. Comp. Biochem. Physiol. Bcomp. Biochem. 1989, 93, 543–547. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Schieber, A.; Wu, J. Effects of storage and cooking on the antioxidant capacity of laying hen eggs. Food Chem. 2016, 194, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Martos, G.; Lopez-Fandino, R.; Molina, E. Immunoreactivity of hen egg allergens: Influence on in vitro gastrointestinal digestion of the presence of other egg white proteins and of egg yolk. Food Chem. 2013, 136, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Vanga, S.K.; Wang, J.; Raghavan, V. Impact of food processing on the structural and allergenic properties of egg white. Trends Food Sci. Technol. 2018, 78, 188–196. [Google Scholar] [CrossRef]
- Juhaimi, F.A.; Uslu, N.; Ozcan, M.M. Oil content and fatty acid composition of eggs cooked in drying oven, microwave and pan. J. Food Sci. Technol. 2017, 54, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Nyemb, K.; Guerin-Dubiard, C.; Pezennec, S.; Jardin, J.; Briard-Bion, V.; Cauty, C.; Rutherfurd, S.M.; Dupont, D.; Nau, F. The structural properties of egg white gels impact the extent of in vitro protein digestion and the nature of peptides generated. Food Hydrocoll. 2016, 54, 315–327. [Google Scholar] [CrossRef]
- Wang, X.F.; Qiu, N.; Liu, Y.P. Effect of Different Heat Treatments on In Vitro Digestion of Egg White Proteins and Identification of Bioactive Peptides in Digested Products. J. Food Sci. 2018, 83, 1140–1148. [Google Scholar] [CrossRef]
- Remanan, M.K.; Wu, J. Antioxidant activity in cooked and simulated digested eggs. Food Funct. 2014, 5, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.F.; Lin, C.C. Production, composition and quality of duck eggs. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Immersel, F.V., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 487–508. [Google Scholar]
- Tserveni-Goussi, A.; Fortomaris, P. Production and quality of quail, pheasant, goose and turkey eggs for uses other than human consumption. In Improving the Safety and Quality of Eggs and Egg Products; Nys, Y., Bain, M., Van Immerseel, F., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2011; Volume 2, pp. 509–537. [Google Scholar]
- Ali, A.H.; Zou, X.; Lu, J.; Abed, S.M.; Yao, Y.; Tao, G.; Jin, Q.; Wang, X. Identification of phospholipids classes and molecular species in different types of egg yolk by using UPLC-Q-TOF-MS. Food Chem. 2017, 221, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Bair, C.W.; Marion, W.W. Yolk cholesterol in eggs from various avian species. Poult. Sci. 1978, 57, 1260–1265. [Google Scholar] [CrossRef] [PubMed]
- Golzar Adabi, S.H.; Ahbab, M.; Fani, A.R.; Hajbabaei, A.; Ceylan, N.; Cooper, R.G. Egg yolk fatty acid profile of avian species--influence on human nutrition. J. Anim. Physiol. Anim. Nutr. 2013, 97, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Qiu, N.; Liu, Y.; Zhao, H.; Gao, D.; Song, R.; Ma, M. Identification and comparative proteomic study of quail and duck egg white protein using 2-dimensional gel electrophoresis and matrix-assisted laser desorption/ionization time-of-flight tandem mass spectrometry analysis. Poult. Sci. 2016, 95, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Qiu, N.; Gao, D.; Ma, M.H. Comparative proteomic analysis of chicken, duck, and quail egg yolks. Int. J. Food Prop. 2018, 21, 1311–1321. [Google Scholar] [CrossRef]
- Sun, C.J.; Liu, J.N.; Li, W.B.; Xu, G.Y.; Yang, N. Divergent Proteome Patterns of Egg Albumen from Domestic Chicken, Duck, Goose, Turkey, Quail and Pigeon. Proteomics 2017, 17, 12. [Google Scholar] [CrossRef]
- Bacon, W.L.; Brown, K.I.; Musser, M.A. Low density lipoproteins of chicken, turkey and quail egg yolk. Poult. Sci. 1973, 52, 1741–1744. [Google Scholar] [CrossRef]
- Appleby, P.N.; Key, T.J. The long-term health of vegetarians and vegans. Proc. Nutr. Soc. 2016, 75, 287–293. [Google Scholar] [CrossRef]
- Key, T.J.; Davey, G.K.; Appleby, P.N. Health benefits of a vegetarian diet. Proc. Nutr. Soc. 1999, 58, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Dinu, M.; Abbate, R.; Gensini, G.F.; Casini, A.; Sofi, F. Vegetarian, vegan diets and multiple health outcomes: A systematic review with meta-analysis of observational studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3640–3649. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Lv, J.; Guo, Y.; Bian, Z.; Si, J.; Yang, L.; Chen, Y.; Zhou, Y.; Zhang, H.; Liu, J.; et al. Associations of egg consumption with cardiovascular disease in a cohort study of 0.5 million Chinese adults. Heart (Br. Card. Soc.) 2018, 104, 1756–1763. [Google Scholar] [CrossRef]
- Orlich, M.J.; Jaceldo-Siegl, K.; Sabate, J.; Fan, J.; Singh, P.N.; Fraser, G.E. Patterns of food consumption among vegetarians and non-vegetarians. Br. J. Nutr. 2014, 112, 1644–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuenemann, E.C.; Mandalari, G.; Rich, G.T.; Faulks, R.M. Dynamic Gastric Model (DGM). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., Lopez-Exposito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 47–59. [Google Scholar] [Green Version]
- Wang, X.; Ye, A.; Lin, Q.; Han, J.; Singh, H. Gastric digestion of milk protein ingredients: Study using an in vitro dynamic model. J. Dairy Sci. 2018, 101, 6842–6852. [Google Scholar] [CrossRef] [PubMed]
- Do, D.H.T.; Kong, F.B.; Penet, C.; Winetzky, D.; Gregory, K. Using a dynamic stomach model to study efficacy of supplemental enzymes during simulated digestion. Lwt-Food Sci. Technol. 2016, 65, 580–588. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
Figure 1.
Egg structure. In italics: eggshell membranes are edible but usually not consumed, as they remain tightly associated with the eggshell.
Figure 1.
Egg structure. In italics: eggshell membranes are edible but usually not consumed, as they remain tightly associated with the eggshell.

Figure 2.
Basic composition of edible parts of the egg. (a) Egg white; (b) Egg yolk. Note that for (b), results refer to egg yolk/vitelline membrane complex. Retrieved on 01/11/2019 from the Ciqual homepage https://ciqual.anses.fr/ (French Agency for Food, Environmental and Occupational Health & Safety. ANSES-CIQUAL).
Figure 2.
Basic composition of edible parts of the egg. (a) Egg white; (b) Egg yolk. Note that for (b), results refer to egg yolk/vitelline membrane complex. Retrieved on 01/11/2019 from the Ciqual homepage https://ciqual.anses.fr/ (French Agency for Food, Environmental and Occupational Health & Safety. ANSES-CIQUAL).

Figure 3.
Physicochemical changes associated with egg storage (freshly laid egg versus egg stored for 2 weeks at room temperature). (a) air cell; (b) major modifications occurring during storage.
Figure 3.
Physicochemical changes associated with egg storage (freshly laid egg versus egg stored for 2 weeks at room temperature). (a) air cell; (b) major modifications occurring during storage.

Figure 4.
Variability of egg composition in five poultry species. Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl/.
Figure 4.
Variability of egg composition in five poultry species. Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl/.

Figure 5.
Vitamin profile of eggs from various domestic species. (a) Vitamins of high abundance; (b) vitamins of low abundance. Note that the concentration of vitamin E, D and K in turkey egg were not available. Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl/.
Figure 5.
Vitamin profile of eggs from various domestic species. (a) Vitamins of high abundance; (b) vitamins of low abundance. Note that the concentration of vitamin E, D and K in turkey egg were not available. Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl/.


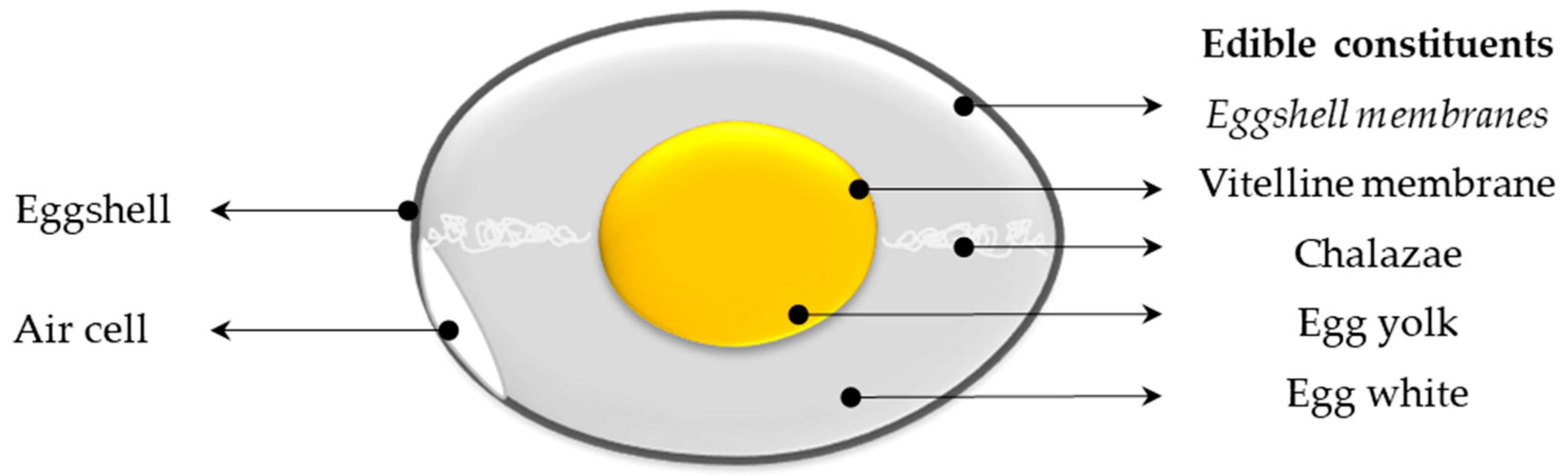{kind=link}
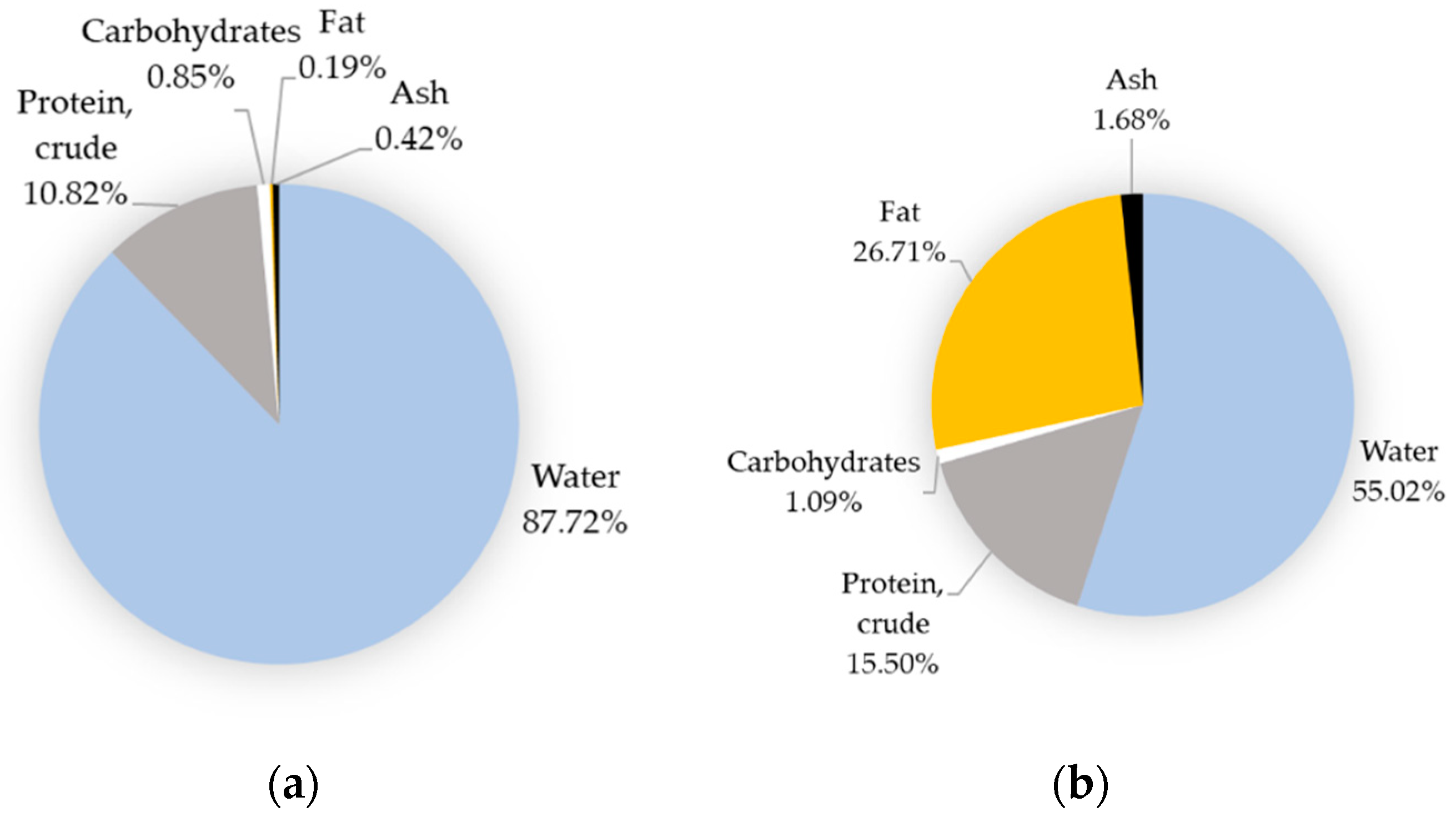{kind=link}
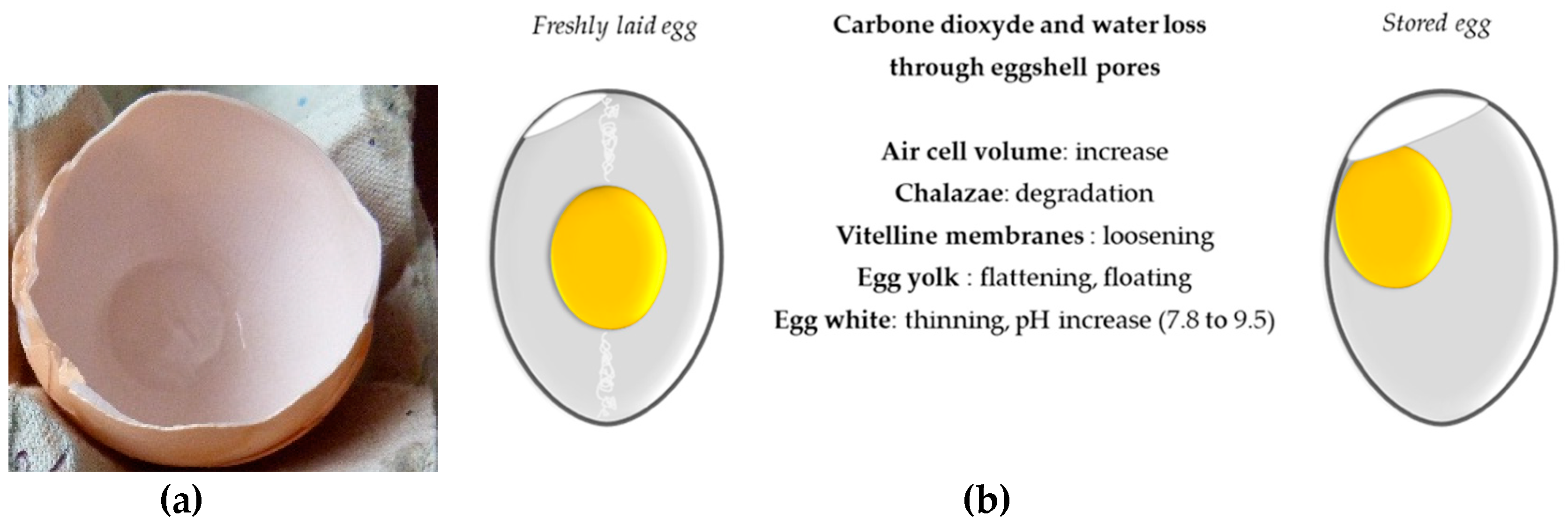{kind=link}
{kind=link}
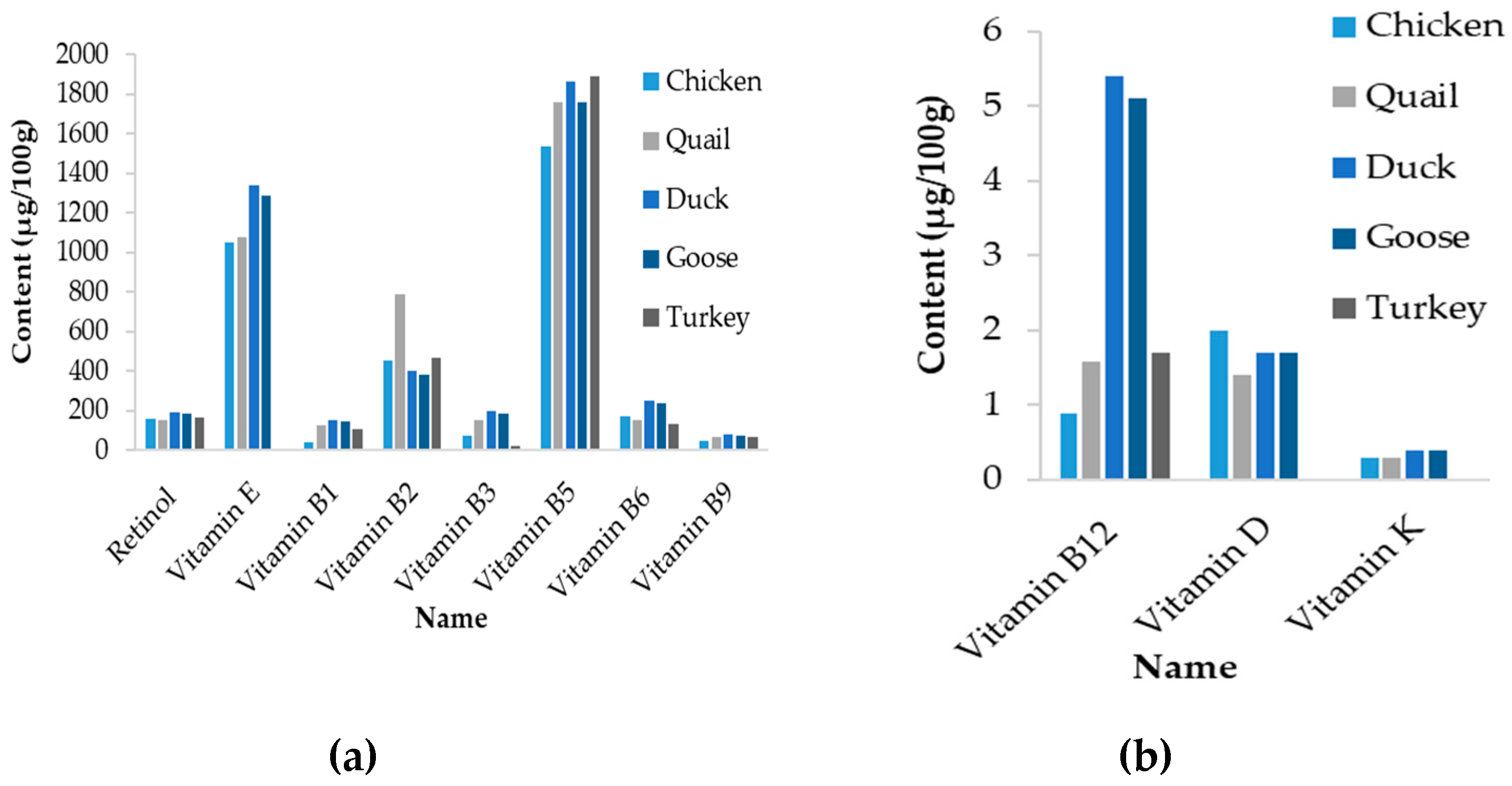{kind=link}
Table 1.
Egg lipids 1.
| Name | Egg, Whole, Raw | Egg Yolk, Raw | ||||
|---|---|---|---|---|---|---|
| Average Content (g/100g) | Min. Value (g/100g) | Max. Value (g/100g) | Average Content (g/100g) | Min. Value (g/100g) | Max. Value (g/100g) | |
| FA saturated | 2.64 | 0.05 | 3.13 | 8.47 | 7.13 | 9.55 |
| FA 4:0 | <0.05 | 0 | - 2 | 0 | - | - |
| FA 6:0 | <0.05 | 0 | - | 0 | - | - |
| FA 8:0 | <0.05 | 0 | - | 0.009 | - | - |
| FA 10:0 | <0.05 | 0 | - | 0.009 | - | - |
| FA 12:0 | <0.05 | 0 | - | 0.009 | - | - |
| FA 14:0 | 0.024 | 0 | 0.038 | 0.091 | 0.077 | 0.1 |
| FA 16:0 | 1.96 | 0.05 | 2.43 | 6.04 | 5.03 | 6.86 |
| FA 18:0 | 0.65 | 0.05 | 0.89 | 1.73 | - | 2.42 |
| FA monounsaturated | 3.66 | 0.05 | 6.73 | 11.9 | 10.2 | 13.8 |
| FA 18:1 n-9 cis | 3.51 | 3.03 | 3.65 | 10.4 | 9.69 | 11.2 |
| FA polyunsaturated | 1.65 | 0.05 | 3.39 | 4.07 | 3.33 | 4.66 |
| FA 18:2 9c,12c (n-6) | 1.38 | 1.18 | 2.7 | 3.28 | - | 3.62 |
| FA 18:3 9c,12c,15c (n-3) | 0.061 | 0.02 | 0.58 | 0.15 | - | 0.27 |
| FA 20:4 5c,8c,11c,14c (n-6) | 0.12 | - | 0.13 | 0.37 | - | 0.4 |
| FA 20:5 5c,8c,11c,14c,17c (n-3) EPA | 0 | - | 0.003 | 0.01 | - | 0.011 |
| FA 22:6 4c,7c,10c,13c,16c,19c (n-3) DHA | 0.09 | 0.045 | 0.18 | 0.25 | 0.11 | 0.46 |
| Cholesterol | 0.398 | 0.344 | 0.423 | 0.939 | 1.280 | |
1 Retrieved on 01/17/2019 from the Ciqual homepage https://ciqual.anses.fr/ (French Agency for Food, Environmental and Occupational Health & Safety. ANSES-CIQUAL). FA, fatty acids; EPA, eicosapentaenoic acid (omega-3 fatty acid); DHA, docosahexaenoic acid (omega-3 fatty acid). 2 not available.
Table 2.
Egg vitamins 1 (average content; µg/100g).
| Name | Egg, Whole, Raw | Egg Yolk, Raw | Egg White, Raw |
|---|---|---|---|
| Vitamin A or Retinol | 160 | 371 | 0 |
| Vitamin A precursor or Beta-carotene | 0 | 88 | 0 |
| Vitamin D or Cholecalciferol | 2.0 | 5.4 | 0 |
| Vitamin E or Alpha-tocopherol | 1050 | 2580 | 0 |
| Vitamin K or Phylloquinone | 0.3 | 0.7 | 0 |
| Vitamin C | 0 | 0 | 0 |
| Vitamin B1 or Thiamin | 40 | 176 | 4 |
| Vitamin B2 or Riboflavin | 457 | 528 | 439 |
| Vitamin B3 or Niacin | 75 | 24 | 105 |
| Vitamin B5 or Pantothenic acid | 1533 | 2990 | 190 |
| Vitamin B6 | 170 | 350 | 5 |
| Vitamin B8 or Biotin | 16.5–53.8 2 | 27.2–49.4 2 | 5.7–7.9 2 |
| Vitamin B9 or Folate | 47 | 146 | 4 |
| Vitamin B12 or Colabamin | 0.89 | 1.95 | 0.09 |
1 Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl 2 [58].
Table 3.
Egg minerals and trace elements (average content; mg/100g) 1.
| Name | Egg, Whole, Raw | Egg Yolk, Raw | Egg White, Raw |
|---|---|---|---|
| Calcium | 56 | 129 | 7 |
| Copper | 0.072 | 0.077 | 0.023 |
| Iodine | 0.021 | 0.18 | 0.002 |
| Iron | 1.75 | 2.73 | 0.08 |
| Magnesium | 12 | 5 | 11 |
| Manganese | 0.028 | 0.055 | 0.011 |
| Phosphorus | 198 | 390 | 15 |
| Potassium | 138 | 109 | 163 |
| Selenium | 0.030 | 0.056 | 0.020 |
| Sodium | 142 | 48 | 166 |
| Zinc | 1.29 | 2.30 | 0.03 |
1 Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27 http://www.ars.usda.gov/ba/bhnrc/ndl and from the Ciqual homepage https://ciqual.anses.fr/ (French Agency for Food, Environmental and Occupational Health & Safety. ANSES-CIQUAL) for iodine content.
Table 4.
Major egg antimicrobial proteins.
| Gene ID/Gene Symbol | Target Organisms | Localization 1 | References | |
|---|---|---|---|---|
| Avian beta-defensin 11 | 414876/AVBD11 | Bacteria | EW, VM | [76,77] |
| Avidin | 396260/AVD | Bacteria | EW, VM, EY | [78,79] |
| Beta-microseminoprotein-like | 101750704 | Bacteria | EW | [76] |
| Cystatin | 396497/CST3 | Bacteria, viruses, fungi, parasites | EW, VM, EY | [75,80,81,82,83,84,85] |
| Gallin | 422030/OvoDA1 | Bacteria | EW | [86,87,88,89] |
| Immunoglobulin Y | - | EY | [90] | |
| Lysozyme | 396218/LYZ | Bacteria, viruses, fungi | EW, VM, EY | [67,91,92] |
| Ovalbumin-related protein X | 420898/OVALX | Bacteria | EW, VM, EY | [93] |
| OvoglobulinG2/TENP | 395882/BPIFB2 | Bacteria | EW, VM, EY | [94,95,96] |
| Ovoinhibitor | 416235/SPIK5 | Bacteria | EW, EY | [97] |
| Ovomucin (alpha and beta subunits) | 395381/LOC395381 (alpha) 414878/MUC6 (beta) | Bacteria, viruses | EW, VM | [74] |
| Ovotransferrin | 396241/TF | Bacteria, viruses | EW, VM, EY | [98,99,100] |
| Phosvitin | 424547/VTG1 | Bacteria | EY | [101] |
| Pleiotrophin | 418125/PTN | Bacteria | EW | [76,102] |
| Vitelline membrane outer layer protein 1 | 418974/VMO1 | Bacteria | EW, VM, EY | [76] |
1 Edible parts: EW, egg white; EY, egg yolk; VM, vitelline membrane.
Table 5.
List of egg characteristics and major components that vary upon cooking 1.
| Name | Egg, Whole, Raw, Fresh | Egg, Whole, Soft-Boiled | Egg, Whole, Hard-Boiled |
|---|---|---|---|
| Energy (kcal/100g) | 140 1; 143 2 | 142; 143 | 134; 155 |
| Protein (g/100g) | 12.7; 12.56 | 12.2; 12.51 | 13.5; 12.58 |
| Carbohydrate (g/100g) | 0.27; 0.72 | 1.08; 0.71 | 0.52; 1.12 |
| Fat (g/100g) | 9.83; 9.51 | 9.82; 9.47 | 8.62; 10.61 |
| FA saturated (g/100g) | 2.64; 3.126 | 3.11; 3.11 | 2.55; 3.267 |
| FA monounsaturated (g/100g) | 3.66; 3.658 | 4.42; 3.643 | 3.57; 4.077 |
| FA polyunsaturated (g/100g) | 1.65; 1.911 | 1.28; 1.904 | 1.03; 1.414 |
| Cholesterol (mg/100g) | 398; 372 | 222; 370 | 355; 373 |
| Salt (g/100g) | 0.31 | 0.2 | 0.31 |
| Calcium (mg/100g) | 76.8; 56 | 150; 56 | 41; 50 |
| Potassium (mg/100g) | 134; 138 | 164; 138 | 120; 126 |
| Selenium (µg/100g) | 30 | 23.8 | 7.01 |
| Vitamin A, Retinol (µg/100g) | 182; 160 | 132; 160 | 61.5; 149 |
| Vitamin D (µg/100g) | 1.88; 2.0 | 1.28; 2.0 | 1.12; 2.2 |
| Vitamin E (mg/100g) | 1.43; 1.05 | 2.17; 1.04 | 1.03; 1.03 |
| Choline (mg/100g) | 250; 293.8 | - | 230; 293.8 |
1 Source: French Agency for Food, Environmental, and Occupational Health & Safety. ANSES-CIQUAL French food composition table version 2017. Retrieved on 01/11/2019 from the Ciqual homepage https://ciqual.anses.fr/; 2 In italics, retrieved on 01/18/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27 and USDA Database for the Choline Content of Common Foods, Release 2 [30]; FA, fatty acid.
Table 6.
Comparison of egg minerals and trace elements in chicken, quail, duck, goose, and turkey eggs (average content; mg/100g).
Table 6.
Comparison of egg minerals and trace elements in chicken, quail, duck, goose, and turkey eggs (average content; mg/100g).
| Name | Chicken | Quail | Duck | Goose | Turkey |
|---|---|---|---|---|---|
| Calcium | 56 | 64 | 64 | 60 | 99 |
| Copper | 0.072 | 0.062 | 0.062 | 0.062 | 0.062 |
| Iron | 1.75 | 3.65 | 3.85 | 3.64 | 4.1 |
| Magnesium | 12 | 13 | 17 | 16 | 13 |
| Manganese | 0.028 | 0.038 | 0.038 | 0.038 | 0.038 |
| Phosphorus | 198 | 226 | 220 | 208 | 170 |
| Potassium | 138 | 132 | 222 | 210 | 142 |
| Selenium | 0.0307 | 0.032 | 0.0364 | 0.0369 | 0.0343 |
| Sodium | 142 | 141 | 146 | 138 | 151 |
| Zinc | 1.29 | 1.47 | 1.41 | 1.33 | 1.58 |
Retrieved on 01/17/2019 from Department of Agriculture, Agricultural Research Service (2014). USDA National Nutrient Database for Standard Reference, Release 27. http://www.ars.usda.gov/ba/bhnrc/ndl.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Réhault-Godbert, S.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030684
AMA Style
Réhault-Godbert S, Guyot N, Nys Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients. 2019; 11(3):684. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030684
Chicago/Turabian StyleRéhault-Godbert, Sophie, Nicolas Guyot, and Yves Nys. 2019. "The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health" Nutrients 11, no. 3: 684. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030684
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.