Methionine Restriction Partly Recapitulates the Sympathetically Mediated Enhanced Energy Expenditure Induced by Total Amino Acid Restriction in Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
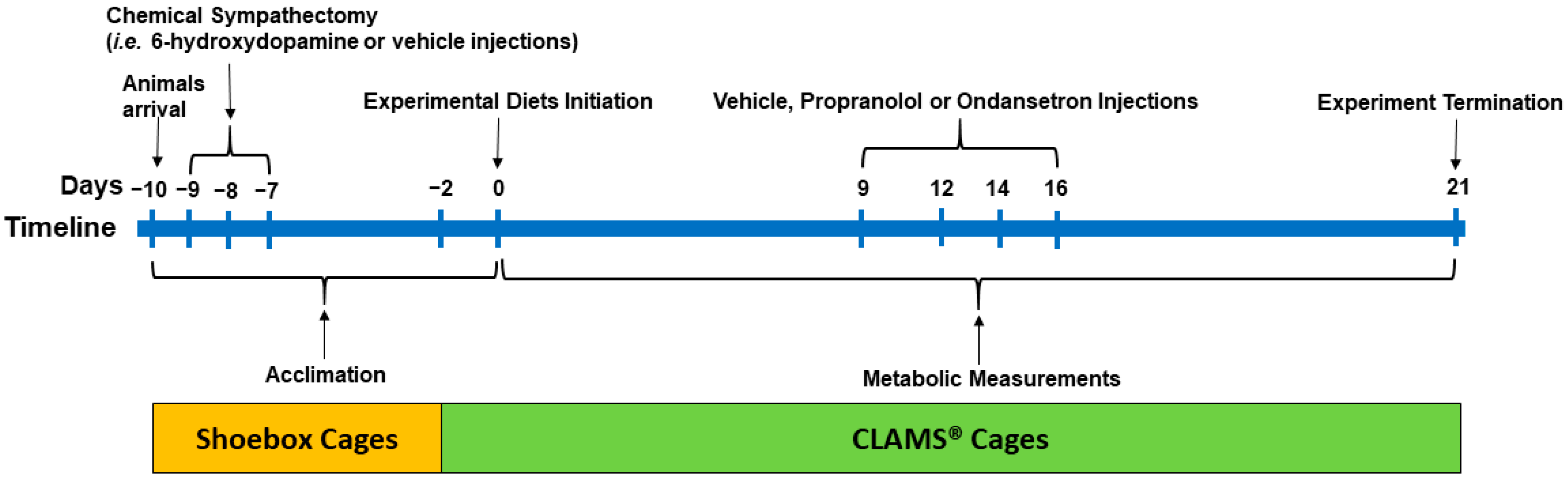
2.1. Animals and Housing
2.2. Chemical Sympathectomy
2.3. Experimental Diets
2.4. Metabolic Measurements
2.5. Blockade of β-AR and 5HT3 Receptors
2.6. Statistical Analysis
3. Results
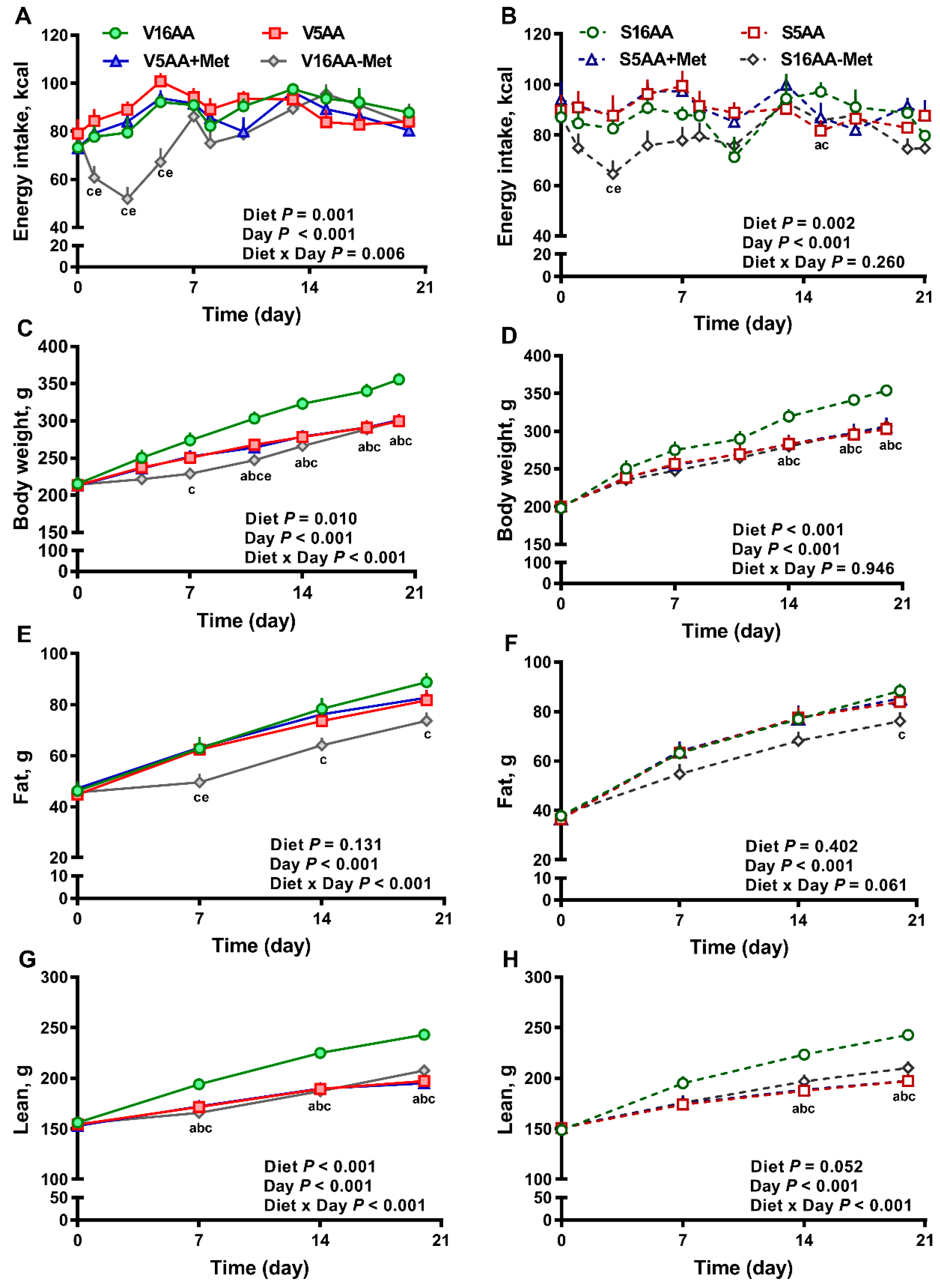
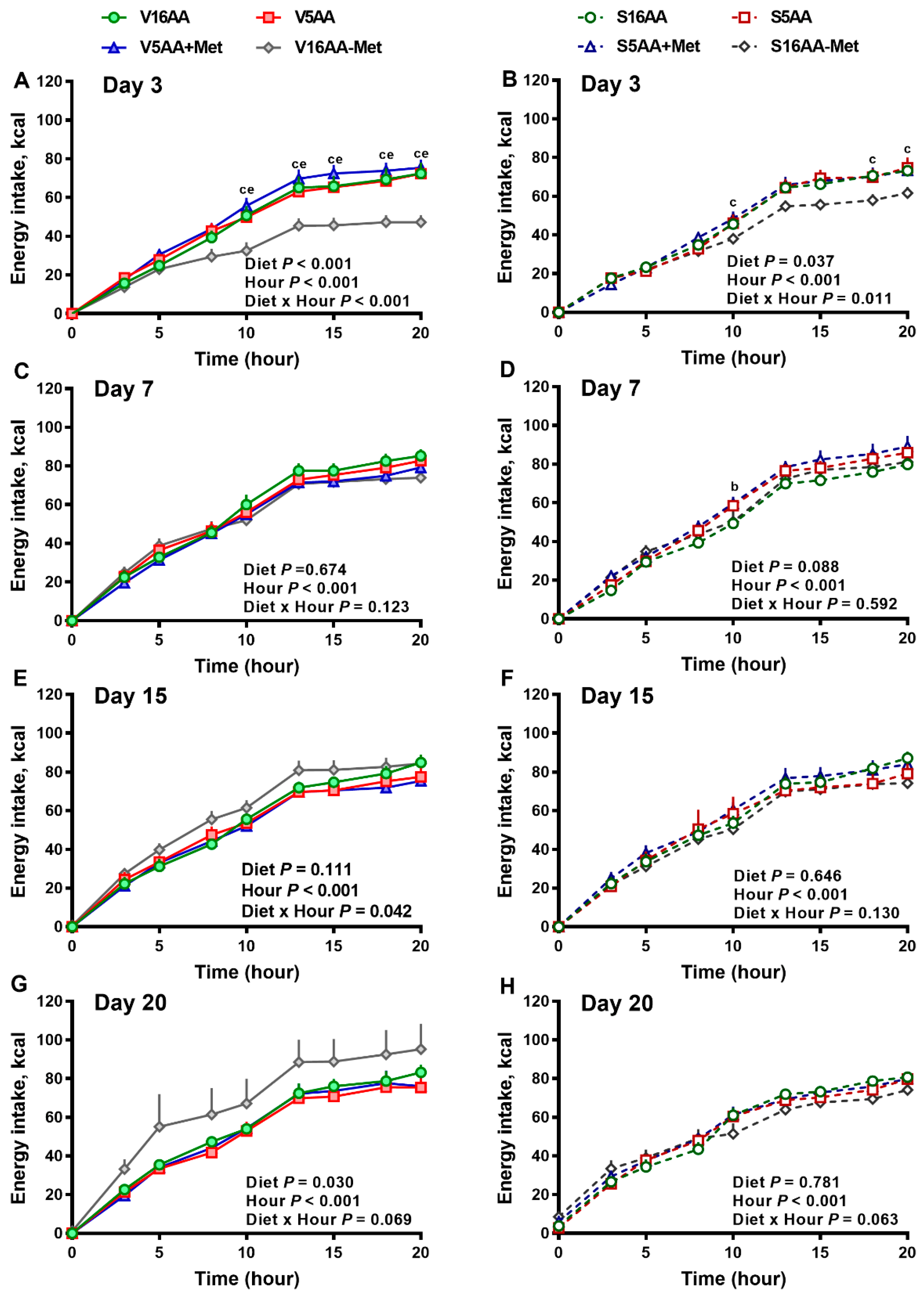
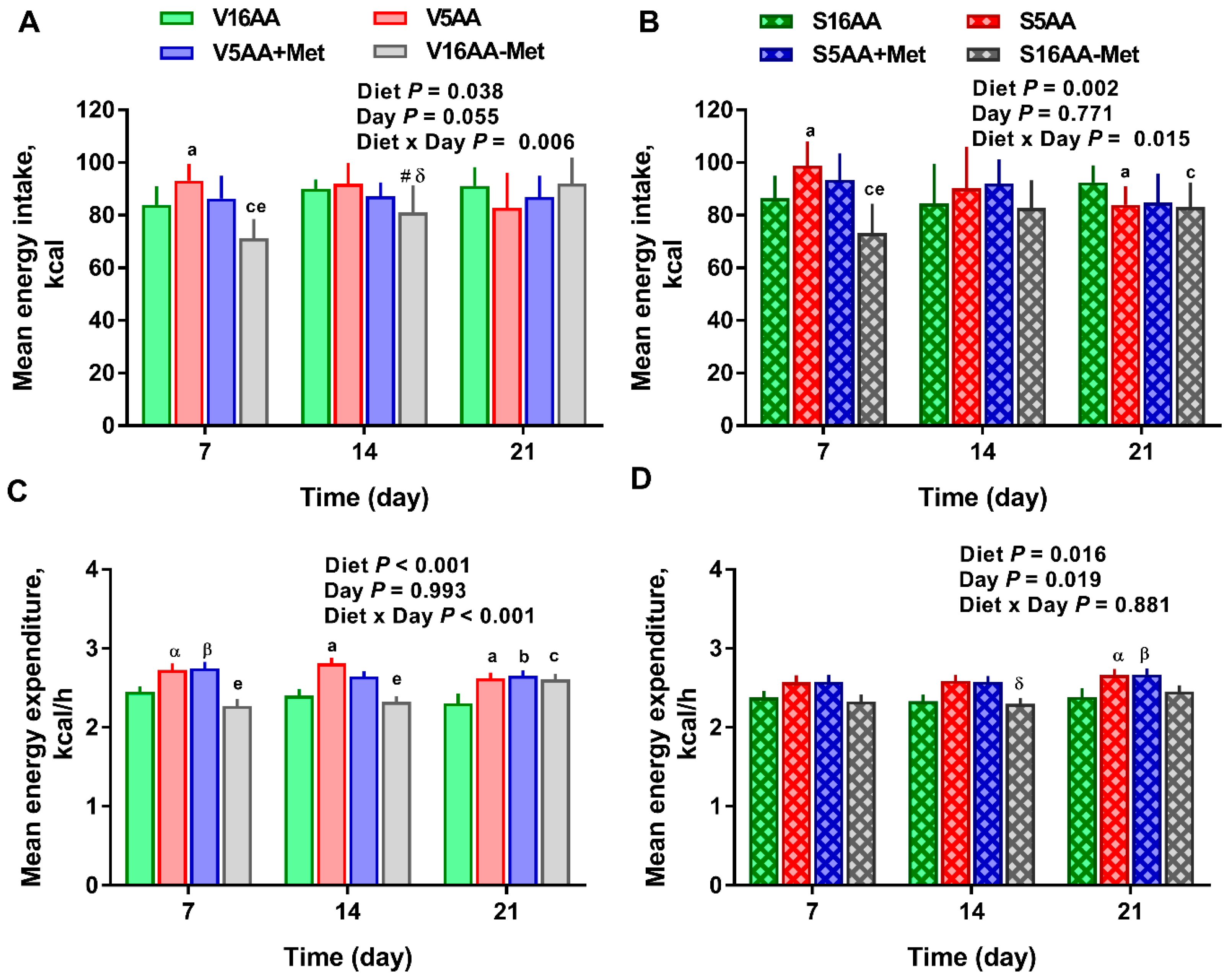
3.1. Energy Intake and Body Composition
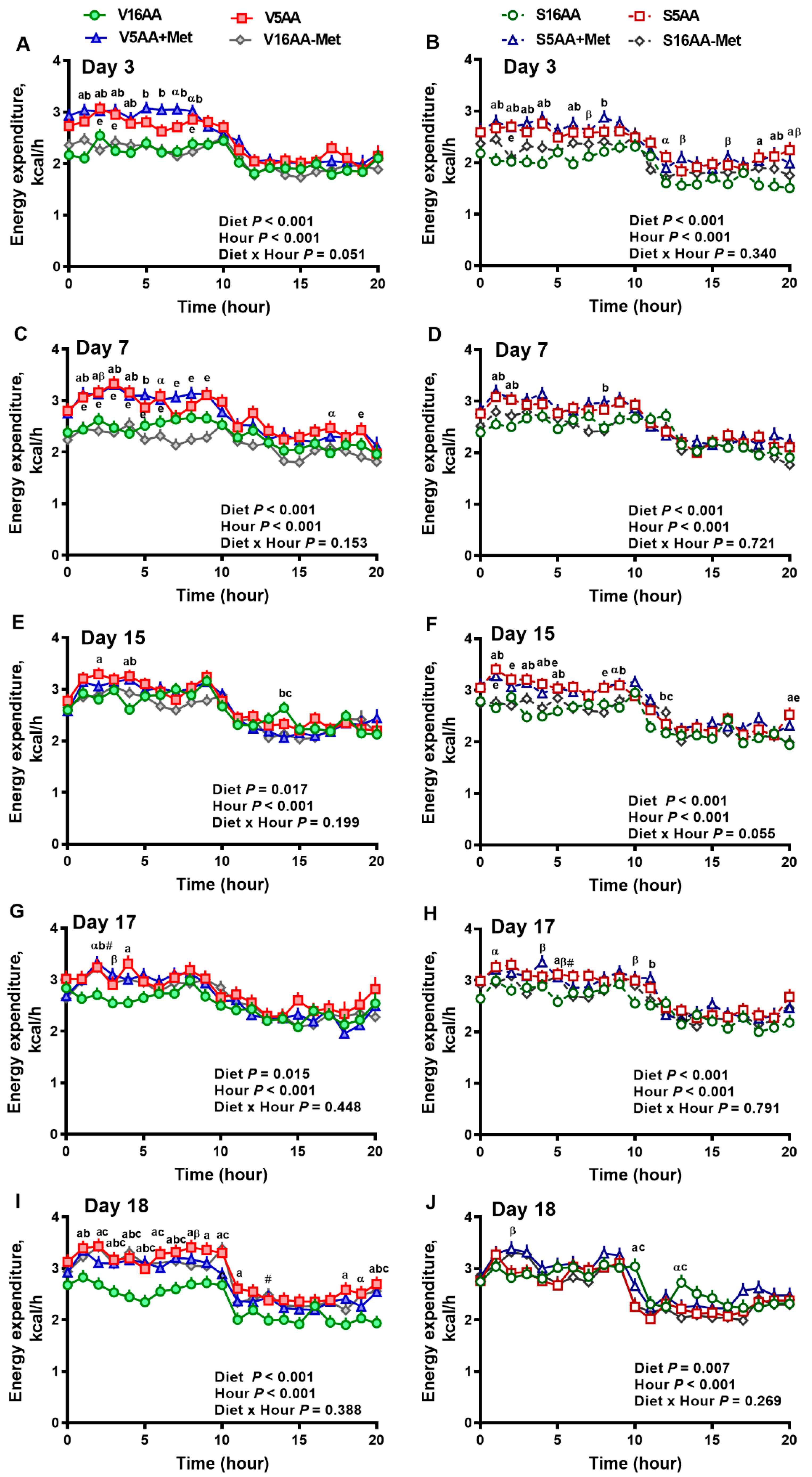
3.2. Energy Expenditure
3.3. β-AR Blockade with Propranolol
3.4. HT3 Receptor Blockade with Ondansetron
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Levine, M.E.; Suarez, J.A.; Brandhorst, S.; Balasubramanian, P.; Cheng, C.W.; Madia, F.; Fontana, L.; Mirisola, M.G.; Guevara-Aguirre, J.; Wan, J.; et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality. Cell Metab. 2014, 19, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.; Hodson, E.M.; Fouque, D. Low protein diets for non-diabetic adults with chronic kidney disease. Cochrane Database Syst. Rev. 2018, 10, CD001892. [Google Scholar] [CrossRef] [PubMed]
- Le Couteur, D.G.; Tay, S.S.; Solon-Biet, S.; Bertolino, P.; McMahon, A.C.; Cogger, V.C.; Colakoglu, F.; Warren, A.; Holmes, A.J.; Pichaud, N.; et al. The influence of macronutrients on splanchnic and hepatic lymphocytes in aging mice. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 70, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; McMahon, A.C.; Ballard, J.W.; Ruohonen, K.; Wu, L.E.; Cogger, V.C.; Warren, A.; Huang, X.; Pichaud, N.; Melvin, R.G.; et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metab. 2014, 19, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; Mitchell, S.J.; Coogan, S.C.; Cogger, V.C.; Gokarn, R.; McMahon, A.C.; Raubenheimer, D.; de Cabo, R.; Simpson, S.J.; Le Couteur, D.G. Dietary protein to carbohydrate ratio and caloric restriction: Comparing metabolic outcomes in mice. Cell Rep. 2015, 11, 1529–1534. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Hancock, D.P.; Gosby, A.K.; McMahon, A.C.; Solon, S.M.; Le Couteur, D.G.; Conigrave, A.D.; Raubenheimer, D.; Simpson, S.J. Effects of dietary protein to carbohydrate balance on energy intake, fat storage, and heat production in mice. Obesity 2013, 21, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. Obesity: The protein leverage hypothesis. Obes. Rev. 2005, 6, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, A.; Zapata, R.C.; Singh, A.; Yee, N.J.; Chelikani, P.K. Low protein diets produced divergent effects on energy balance. Sci. Rep. 2016, 6, 25145. [Google Scholar] [CrossRef] [PubMed]
- White, B.D.; He, B.; Dean, R.G.; Martin, R.J. Low protein diets increase neuropeptide Y gene expression in the basomedial hypothalamus of rats. J. Nutr. 1994, 124, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Gosby, A.K.; Conigrave, A.D.; Lau, N.S.; Iglesias, M.A.; Hall, R.M.; Jebb, S.A.; Brand-Miller, J.; Caterson, I.D.; Raubenheimer, D.; Simpson, S.J. Testing protein leverage in lean humans: A randomised controlled experimental study. PLoS ONE 2011, 6, e25929. [Google Scholar] [CrossRef]
- Griffioen-Roose, S.; Mars, M.; Siebelink, E.; Finlayson, G.; Tomé, D.; de Graaf, C. Protein status elicits compensatory changes in food intake and food preferences. Am. J. Clin. Nutr. 2012, 95, 32–38. [Google Scholar] [CrossRef]
- Griffioen-Roose, S.; Smeets, P.A.; van den Heuvel, E.; Boesveldt, S.; Finlayson, G.; de Graaf, C. Human protein status modulates brain reward responses to food cues. Am. J. Clin. Nutr. 2014, 100, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanders, D.; Burk, D.H.; Cortez, C.C.; Van, N.T.; Stone, K.P.; Baker, M.; Mendoza, T.; Mynatt, R.L.; Gettys, T.W. Ucp1 is an essential mediator of the effects of methionine restriction on energy balance but not insulin sensitivity. FASEB J. 2015, 29, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Invest. 2014, 124, 3913–3922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothwell, N.J.; Stock, M.J. Influence of carbohydrate and fat intake on diet-induced thermogenesis and brown fat activity in rats fed low protein diets. J. Nutr. 1987, 117, 1721–1726. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, N.J.; Stock, M.J.; Tyzbir, R.S. Energy balance and mitochondrial function in liver and brown fat of rats fed “cafeteria” diets of varying protein content. J. Nutr. 1982, 112, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, N.J.; Stock, M.J.; Tyzbir, R.S. Mechanisms of thermogenesis induced by low protein diets. Metabolism 1983, 32, 257–261. [Google Scholar] [CrossRef]
- Aparecida de França, S.; Dos Santos, M.P.; Garófalo, M.A.; Navegantes, L.C.; Kettelhut Ido, C.; Lopes, C.F.; Kawashita, N.H. Low protein diet changes the energetic balance and sympathetic activity in brown adipose tissue of growing rats. Nutrition 2009, 25, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Higginbotham, A.; White, D. Food intake, energy balance and serum leptin concentrations in rats fed low-protein diets. J. Nutr. 2000, 130, 514–521. [Google Scholar] [CrossRef] [PubMed]
- White, B.D.; Porter, M.H.; Martin, R.J. Protein selection, food intake, and body composition in response to the amount of dietary protein. Physiol. Behav. 2000, 69, 383–389. [Google Scholar] [CrossRef]
- White, B.D.; Porter, M.H.; Martin, R.J. Effects of age on the feeding response to moderately low dietary protein in rats. Physiol. Behav. 2000, 68, 673–681. [Google Scholar] [CrossRef]
- Peng, Y.S.; Meliza, L.L.; Vavich, M.G.; Kemmerer, A.R. Changes in food intake and nitrogen metabolism of rats while adapting to a low or high protein diet. J. Nutr. 1974, 104, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Uppal, S.O.; Moorman, J.L.; Bennett, C.; Gruca, L.L.; Parimi, P.S.; Dasarathy, S.; Serre, D.; Hanson, R.W. Metabolic and genomic response to dietary isocaloric protein restriction in the rat. J. Biol. Chem. 2011, 286, 5266–5277. [Google Scholar] [CrossRef]
- Anthony, T.G.; Gietzen, D.W. Detection of amino acid deprivation in the central nervous system. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 96–101. [Google Scholar] [CrossRef]
- Glick, Z.; Teague, R.J.; Bray, G.A. Brown adipose tissue: Thermic response increased by a single low protein, high carbohydrate meal. Science 1981, 213, 1125–1127. [Google Scholar] [CrossRef] [PubMed]
- Gietzen, D.W.; Hao, S.; Anthony, T.G. Mechanisms of food intake repression in indispensable amino acid deficiency. Annu. Rev. Nutr. 2007, 27, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Mercer, L.P.; Kelley, D.S.; Haq, A.; Humphries, L.L. Dietary induced anorexia: A review of involvement of the histaminergic system. J. Am. Coll. Nutr. 1996, 15, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Maurin, A.C.; Benani, A.; Lorsignol, A.; Brenachot, X.; Parry, L.; Carraro, V.; Guissard, C.; Averous, J.; Jousse, C.; Bruhat, A.; et al. Hypothalamic eIF2α signaling regulates food intake. Cell. Rep. 2014, 6, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Laeger, T.; Reed, S.D.; Henagan, T.M.; Fernandez, D.H.; Taghavi, M.; Addington, A.; Münzberg, H.; Martin, R.J.; Hutson, S.M.; Morrison, C.D. Leucine acts in the brain to suppress food intake but does not function as a physiological signal of low dietary protein. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R310–R320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malloy, V.L.; Krajcik, R.A.; Bailey, S.J.; Hristopoulos, G.; Plummer, J.D.; Orentreich, N. Methionine restriction decreases visceral fat mass and preserves insulin action in aging male Fischer 344 rats independent of energy restriction. Aging Cell 2006, 5, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasek, B.E.; Stewart, L.K.; Henagan, T.M.; Boudreau, A.; Lenard, N.R.; Black, C.; Shin, J.; Huypens, P.; Malloy, V.L.; Plaisance, E.P.; et al. Dietary methionine restriction enhances metabolic flexibility and increases uncoupled respiration in both fed and fasted states. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, E.P.; Henagan, T.M.; Echlin, H.; Boudreau, A.; Hill, K.L.; Lenard, N.R.; Hasek, B.E.; Orentreich, N.; Gettys, T.W. Role of beta-adrenergic receptors in the hyperphagic and hypermetabolic responses to dietary methionine restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R740–R750. [Google Scholar] [CrossRef] [PubMed]
- Orentreich, N.; Matias, J.R.; DeFelice, A.; Zimmerman, J.A. Low methionine ingestion by rats extends life span. J. Nutr. 1993, 123, 269–274. [Google Scholar] [PubMed]
- Zimmerman, J.A.; Malloy, V.; Krajcik, R.; Orentreich, N. Nutritional control of aging. Exp. Gerontol. 2003, 38, 47–52. [Google Scholar] [CrossRef]
- Wanders, D.; Forney, L.A.; Stone, K.P.; Burk, D.H.; Pierse, A.; Gettys, T.W. FGF21 mediates the thermogenic and insulin-sensitizing effects of dietary methionine restriction but not its effects on hepatic lipid metabolism. Diabetes 2017, 66, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Ables, G.P.; Johnson, J.E. Pleiotropic responses to methionine restriction. Exp. Gerontol. 2017, 94, 83–88. [Google Scholar] [CrossRef]
- Lee, B.C.; Kaya, A.; Ma, S.; Kim, G.; Gerashchenko, M.V.; Yim, S.H.; Hu, Z.; Harshman, L.G.; Gladyshev, V.N. Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nat. Commun. 2014, 5, 3592. [Google Scholar] [CrossRef] [PubMed]
- Grandison, R.C.; Piper, M.D.; Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 2009, 462, 1061–1064. [Google Scholar] [CrossRef]
- Forney, L.; Wanders, D.; Stone, K.; Pierse, A.; Gettys, T. Concentration-dependent linkage of dietary methionine restriction to the components of its metabolic phenotype. Obesity 2017, 25, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Lees, E.K.; Król, E.; Grant, L.; Shearer, K.; Wyse, C.; Moncur, E.; Bykowska, A.S.; Mody, N.; Gettys, T.W.; Delibegovic, M. Methionine restriction restores a younger metabolic phenotype in adult mice with alterations in fibroblast growth factor 21. Aging Cell 2014, 13, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Cummings, N.E.; Williams, E.M.; Kasza, I.; Konon, E.N.; Schaid, M.D.; Schmidt, B.A.; Poudel, C.; Sherman, D.S.; Yu, D.; et al. Restoration of metabolic health by decreased consumption of branched-chain amino acids. J. Physiol. 2018, 596, 623–645. [Google Scholar] [CrossRef]
- Fontana, L.; Cummings, N.E.; Arriola Apelo, S.I.; Neuman, J.C.; Kasza, I.; Schmidt, B.A.; Cava, E.; Spelta, F.; Tosti, V.; Syed, F.A.; et al. Decreased consumption of branched-chain amino acids improves metabolic health. Cell. Rep. 2016, 16, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Forney, L.A.; Stone, K.P.; Wanders, D.; Gettys, T.W. Sensing and signaling mechanisms linking dietary methionine restriction to the behavioral and physiological components of the response. Front. Neuroendocrinol. 2018, 51, 36–45. [Google Scholar] [CrossRef]
- Maida, A.; Chan, J.S.K.; Sjøberg, K.A.; Zota, A.; Schmoll, D.; Kiens, B.; Herzig, S.; Rose, A.J. Repletion of branched chain amino acids reverses mTORC1 signaling but not improved metabolism during dietary protein dilution. Mol. Metab. 2017, 6, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Finch, L.; Haeusler, G.; Kuhn, H.; Thoenen, H. Rapid recovery of vascular adrenergic nerves in the rat after chemical sympathectomy with 6-hydroxydopamine. Br. J. Pharmacol. 1973, 48, 59–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezeshki, A.; Fahim, A.; Chelikani, P.K. Dietary whey and casein differentially affect energy balance, gut hormones, glucose metabolism, and taste preference in diet-induced obese rats. J. Nutr. 2015, 145, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, D.L.; Stevens, S.Y.; Thyaga Rajan, S.; Lorton, D.; Madden, K.S. Aging and sympathetic modulation of immune function in Fischer 344 rats: Effects of chemical sympathectomy on primary antibody response. J. Neuroimmunol. 2005, 165, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Even, P.C.; Nadkarni, N.A. Indirect calorimetry in laboratory mice and rats: Principles, practical considerations, interpretation and perspectives. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R459–R476. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Stat. Soc. Series B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Donato, K.; Hegsted, D.M. Efficiency of utilization of various sources of energy for growth. Proc. Natl. Acad. Sci. USA 1985, 82, 4866–4870. [Google Scholar] [CrossRef]
- Blais, A.; Chaumontet, C.; Azzout-Marniche, D.; Piedcoq, J.; Fromentin, G.; Gaudichon, C.; Tomé, D.; Even, P.C. Low-protein diet-induced hyperphagia and adiposity are modulated through interactions involving thermoregulation, motor activity, and protein quality in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E139–E151. [Google Scholar] [CrossRef]
- Anthony, T.G. Homeostatic responses to amino acid insufficiency. Anim. Nutr. 2015, 1, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Gallinetti, J.; Harputlugil, E.; Mitchell, J.R. Amino acid sensing in dietary-restriction-mediated longevity: Roles of signal-transducing kinases GCN2 and TOR. Biochem. J. 2013, 449, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Anthony, T.G.; Reiter, A.K.; Anthony, J.C.; Kimball, S.R.; Jefferson, L.S. Deficiency of dietary EAA preferentially inhibits mRNA translation of ribosomal proteins in liver of meal-fed rats. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E430–E439. [Google Scholar] [CrossRef] [PubMed]
- Anthony, T.G.; McDaniel, B.J.; Byerley, R.L.; McGrath, B.C.; Cavener, D.R.; McNurlan, M.A.; Wek, R.C. Preservation of liver protein synthesis during dietary leucine deprivation occurs at the expense of skeletal muscle mass in mice deleted for eIF2 kinase GCN2. J. Biol. Chem. 2004, 279, 36553–36561. [Google Scholar] [CrossRef]
- Guo, F.; Cavener, D.R. The GCN2 eIF2alpha kinase regulates fatty-acid homeostasis in the liver during deprivation of an essential amino acid. Cell Metab. 2007, 5, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Chaumontet, C.; Azzout-Marniche, D.; Blais, A.; Piedcoq, J.; Tomé, D.; Gaudichon, C.; Even, P.C. Low protein and methionine, high starch diets increase energy intake and expenditure, increase FGF21, decrease IGF-1, and has little effect on adiposity in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Ji, T.; Guo, H.; Liu, S.M.; Li, Y.; Zheng, L.F.; Zhang, Y.; Zhang, X.F.; Duan, D.P. Expression and activation of β-adrenoceptors in the colorectal mucosa of rat and human. Neurogastroenterol. Motil. 2010, 22, e325–e334. [Google Scholar] [CrossRef] [PubMed]
- Denroche, H.C.; Kwon, M.M.; Glavas, M.M.; Tudurí, E.; Philippe, M.; Quong, W.L.; Kieffer, T.J. The role of autonomic efferents and uncoupling protein 1 in the glucose-lowering effect of leptin therapy. Mol. Metab. 2016, 5, 716–724. [Google Scholar] [CrossRef] [PubMed]
- De Marte, M.L.; Enesco, H.E. Influence of diet on plasma tryptophan and brain serotonin levels in mice. Experientia 1985, 41, 48–50. [Google Scholar] [CrossRef]
- Segall, P.E.; Timiras, P.S. Patho-physiologic findings after chronic tryptophan deficiency in rats: A model for delayed growth and aging. Mech. Ageing. Dev. 1976, 5, 109–124. [Google Scholar] [CrossRef]
- Zapata, R.C.; Singh, A.; Ajdari, N.M.; Chelikani, P.K. Dietary tryptophan restriction dose-dependently modulates energy balance, gut hormones, and microbiota in obesity-prone rats. Obesity 2018, 26, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Venero, J.L.; Herrera, A.J.; Machado, A.; Cano, J. Changes in neurotransmitter levels associated with the deficiency of some essential amino acids in the diet. Br. J. Nutr. 1992, 68, 409–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, J.M.; Struyker-Boudier, H.A. Propranolol in conscious spontaneously hypertensive rats. II. Disposition after subcutaneous and intracerebroventricular administration. Naunyn. Schmiedebergs. Arch. Pharmacol. 1979, 309, 19–24. [Google Scholar] [CrossRef]
- Shand, D.G. Pharmacokinetics of propranolol: A review. Postgrad. Med. J. 1976, 4, 22–25. [Google Scholar]
- Choudhary, D.R.; Patel, V.A.; Chhalotiya, U.K.; Patel, H.V.; Kundawala, A.J. Comparative pharmacokinetic studies of fast dissolving film and oral solution of ondansetron in rats. Curr Drug Deliv. 2013, 10, 696–700. [Google Scholar] [CrossRef] [PubMed]
- Perrone, C.; Malloy, V.; Orentreich, D.; Orentreich, N. Metabolic adaptions to methionine restriction that benefit health and lifespan in rodents. Exp. Gerontol. 2013, 48, 654–660. [Google Scholar] [CrossRef]
- Maida, A.; Zota, A.; Sjøberg, K.A.; Schumacher, J.; Sijmonsma, T.P.; Pfenninger, A.; Christensen, M.M.; Gantert, T.; Fuhrmeister, J.; Rothermel, U.; et al. A liver stress-endocrine nexus promotes metabolic integrity during dietary protein dilution. J. Clin. Invest. 2016, 126, 3263–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaisance, E.P.; Greenway, F.L.; Boudreau, A.; Hill, K.L.; Johnson, W.D.; Krajcik, R.A.; Perrone, C.E.; Orentreich, N.; Cefalu, W.T.; Gettys, T.W. Dietary methionine restriction increases fat oxidation in obese adults with metabolic syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E836–E840. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 16AA | 5AA | 5AA+Met | 16AA-Met | |
|---|---|---|---|---|
| Ingredients (g kg−1) 1 | ||||
| Corn starch | 363.94 | 482.98 | 479.93 | 366.98 |
| L-Alanine | 4.57 | 1.52 | 1.52 | 4.57 |
| L-Arginine | 6.36 | 2.12 | 2.12 | 6.36 |
| L-Aspartic acid | 12.13 | 4.04 | 4.04 | 12.13 |
| L-Cystine | 3.68 | 1.23 | 1.23 | 3.68 |
| L-Glutamic acid | 36.10 | 12.03 | 12.03 | 36.10 |
| Glycine | 3.18 | 1.06 | 1.06 | 3.18 |
| L-Histidine | 4.57 | 1.52 | 1.52 | 4.57 |
| L-Isoleucine | 8.45 | 2.82 | 2.82 | 8.45 |
| L-Leucine | 15.31 | 5.10 | 5.10 | 15.31 |
| L-Lysine | 12.93 | 4.31 | 4.31 | 12.93 |
| D,L-Methionine | 4.57 | 1.52 | 4.57 | 1.52 |
| L-Phenylalanine | 8.75 | 2.92 | 2.92 | 8.75 |
| L-Proline | 20.38 | 6.79 | 6.79 | 20.38 |
| L-Serine | 9.64 | 3.21 | 3.21 | 9.64 |
| L-Threonine | 6.66 | 2.22 | 2.22 | 6.66 |
| L-Tryptophan | 2.09 | 0.70 | 0.70 | 2.09 |
| L-Tyrosine | 9.25 | 3.08 | 3.08 | 9.25 |
| L-Valine | 9.95 | 3.32 | 3.32 | 9.95 |
| Sucrose | 200.00 | 200.00 | 200.00 | 200.00 |
| Corn oil | 60.00 | 60.00 | 60.00 | 60.00 |
| Lard | 100.00 | 100.00 | 100.00 | 100.00 |
| α-Cellulose | 50.00 | 50.00 | 50.00 | 50.00 |
| AIN-93-MX | 35.00 | 35.00 | 35.00 | 35.00 |
| AIN-93-VX | 10.00 | 10.00 | 10.00 | 10.00 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 | 2.50 |
| Composition | ||||
| Protein (% kcal) 2 | 16.20% | 5.40% | 5.68% | 15.92% |
| Carbohydrate (% kcal) 2 | 51.15% | 61.95% | 61.67% | 51.43% |
| Fat (% kcal) 2 | 32.65% | 32.65% | 32.65% | 32.65% |
| Energy density, kcal/g 2 | 4.41 | 4.41 | 4.41 | 4.41 |
| Methionine (DL)% diet 3 | 0.46 | 0.15 | 0.46 | 0.15 |
| Time | p-Values | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Variable | Diet | Week 1 | Week 2 | Week 3 | Diet | Week | Diet × Week |
| Vehicle (V) | Food Conversion Rate 2 (kcal intake/kcal energy deposited) | V16AA | 1.7 ± 0.3 | 2.2 ± 0.3 | 3.6 ± 0.3 | <0.01 | <0.01 | <0.01 |
| V5AA | 2.4 ± 0.3a | 3.5 ± 0.3a | 4.1 ± 0.4 | |||||
| V5AA+Met | 2.4 ± 0.3b | 3.2 ± 0.3b | 5.4 ± 0.4 | |||||
| V16AA-Met | 5.3 ± 0.4ce | 2.3 ± 0.3e | 3.5 ± 0.3 | |||||
| Relative Energy Intake 3 (kcal intake/100 g body weight) | V16AA | 33.4 ± 0.7 | 29.1 ± 1.5 | 24.5 ± 1.4 | 0.08 | <0.01 | 0.59 | |
| V5AA | 37.8 ± 2.0a | 29.9 ± 1.5 | 28.1 ± 1.5 | |||||
| V5AA+Met | 36.4 ± 1.3b | 32.3 ± 2.1 | 26.5 ± 1.9 | |||||
| V16AA-Met | 37.9 ± 4.8 | 35.9 ± 1.6ce | 27.0 ± 1.8 | |||||
| Energy Efficiency 4 (kcal energy deposited/kcal intake) | V16AA | 0.59 ± 0.02 | 0.47 ± 0.02 | 0.30 ± 0.02 | <0.01 | <0.01 | <0.01 | |
| V5AA | 0.41 ± 0.02a | 0.30 ± 0.02a | 0.25 ± 0.03 | |||||
| V5AA+Met | 0.42 ± 0.02b | 0.35 ± 0.02b | 0.20 ± 0.03 | |||||
| V16AA-Met | 0.21 ± 0.03ce | 0.44 ± 0.02e | 0.29 ± 0.02 | |||||
| 6-OHDA (S) | Food Conversion Rate 2 (kcal intake/kcal energy deposited) | S16AA | 1.3 ± 0.3 | 2.1 ± 0.3 | 3.3 ± 0.3 | <0.01 | <0.01 | 0.23 |
| S5AA | 1.8 ± 0.3a | 3.3 ± 0.3a | 5.2 ± 0.3a | |||||
| S5AA+Met | 1.7 ± 0.3b | 3.6 ± 0.3b | 5.1 ± 0.3b | |||||
| S16AA-Met | 1.9 ± 0.3c | 2.6 ± 0.3ce | 4.5 ± 0.3c | |||||
| Relative Energy Intake 3 (kcal intake/100 g body weight) | S16AA | 32.4 ± 1.7 | 29.6 ± 3.1 | 24.8 ± 0.9 | <0.01 | <0.01 | 0.29 | |
| S5AA | 38.8 ± 1.7a | 32.1 ± 2.6 | 25.8 ± 1.2 | |||||
| S5AA+Met | 38.3 ± 1.3b | 35.6 ± 1.3 | 29.1 ± 1.0b | |||||
| S16AA-Met | 31.5±1.8e | 33.7 ± 3.3 | 23.4 ± 1.0 | |||||
| Energy Efficiency 4 (kcal energy deposited/kcal intake) | S16AA | 0.77 ± 0.03 | 0.43 ± 0.03 | 0.31 ± 0.03 | <0.01 | <0.01 | 0.12 | |
| S5AA | 0.59 ± 0.03a | 0.32 ± 0.03 | 0.20 ± 0.03a | |||||
| S5AA+Met | 0.61 ± 0.03b | 0.31 ± 0.03 | 0.21 ± 0.03b | |||||
| S16AA-Met | 0.59 ± 0.03c | 0.32 ± 0.03 | 0.20 ± 0.03c | |||||
| Treatment | Diet | Time | Drug | Energy Balance | |
|---|---|---|---|---|---|
| Energy Intake kcal × h | Energy Expenditure (kcal/ h) × h | ||||
| Vehicle (V) | V16AA | Dark | Saline | 76 ± 6 | 36 ± 1 |
| Propranolol | 62 ± 6 | 32 ± 1 * | |||
| Light | Saline | 16 ± 2 | 21 ± 1 | ||
| Propranolol | 17 ± 2 | 20 ± 1 | |||
| V5AA | Dark | Saline | 73 ± 3 | 39 ± 1 | |
| Propranolol | 68 ± 7 | 33 ± 1 * | |||
| Light | Saline | 16 ± 1 | 23 ± 1 | ||
| Propranolol | 13 ± 2 | 21 ± 1 | |||
| V5AA+Met | Dark | Saline | 74 ± 4 | 37 ± 0.9 | |
| Propranolol | 56 ± 2 * | 33 ± 0.9 * | |||
| Light | Saline | 11 ± 2 | 20 ± 0.9 | ||
| Propranolol | 10 ± 2 | 20 ± 0.9 | |||
| V16AA-Met | Dark | Saline | 79 ± 9 | 30 ± 1 | |
| Propranolol | 81 ± 10 | 28 ± 1 | |||
| Light | Saline | 10 ± 2 | 19 ± 1 | ||
| Propranolol | 16 ± 2 | 18 ± 1 | |||
| 6-OHDA (S) | S16AA | Dark | Saline | 74 ± 3 | 35 ± 1.8 |
| Propranolol | 69 ± 8 | 31 ± 1.8 | |||
| Light | Saline | 16 ± 1 | 19 ± 1.8 | ||
| Propranolol | 13 ± 2 | 18 ± 1.8 | |||
| S5AA | Dark | Saline | 65 ± 4 | 37 ± 1.7 | |
| Propranolol | 53 ± 3 * | 28 ± 1.7 * | |||
| Light | Saline | 21 ± 4 | 21 ± 1.7 | ||
| Propranolol | 12 ± 3 | 18 ± 1.7 | |||
| S5AA+Met | Dark | Saline | 76 ± 4 | 37 ± 1.7 | |
| Propranolol | 55 ± 4 * | 30 ± 1.7 * | |||
| Light | Saline | 14 ± 2 | 20 ± 1.7 | ||
| Propranolol | 12 ± 3 | 20 ± 1.7 | |||
| S16AA-Met | Dark | Saline | 69 ± 4 | 35 ± 1.7 | |
| Propranolol | 48 ± 5 * | 29 ± 1.7 | |||
| Light | Saline | 11 ± 2 | 19 ± 1.7 | ||
| Propranolol | 11 ± 2 | 18 ± 1.7 | |||
| Treatment | Diet | Time | Drug | Energy Balance | |
|---|---|---|---|---|---|
| Energy Intake | Energy Expenditure | ||||
| kcal × h | (kcal/ h) × h | ||||
| Vehicle (V) | V-16AA | Dark | Saline | 63 ± 3 | 37 ± 3 |
| Ondansetron | 74 ± 5 | 34 ± 3 | |||
| Light | Saline | 20 ± 3 | 21 ± 3 | ||
| Ondansetron | 14 ± 1 * | 20 ± 3 | |||
| V-5AA | Dark | Saline | 56 ± 5 | 30 ± 2 | |
| Ondansetron | 69 ± 3 | 38 ± 3 | |||
| Light | Saline | 19 ± 3 | 17 ± 2 | ||
| Ondansetron | 17 ± 1 | 22 ± 3 | |||
| V-5AA+Met | Dark | Saline | 60 ± 3 | 41 ± 2 | |
| Ondansetron | 68 ± 4 | 37 ± 2 | |||
| Light | Saline | 16 ± 3 | 21 ± 2 | ||
| Ondansetron | 10 ± 1 | 20 ± 2 | |||
| V-16AA-Met | Dark | Saline | 69 ± 7 | 41 ± 2 | |
| Ondansetron | 76 ± 4 | 33 ± 2 | |||
| Light | Saline | 20 ± 3 | 23 ± 2 | ||
| Ondansetron | 10 ± 2 * | 19 ± 2 | |||
| 6-OHDA (S) | S-16AA | Dark | Saline | 67 ± 3 | 38 ± 1 |
| Ondansetron | 65 ± 5 | 31 ± 1 * | |||
| Light | Saline | 17 ± 2 | 21 ± 1 | ||
| Ondansetron | 14 ± 1 | 18 ± 1 | |||
| S-5AA | Dark | Saline | 70 ± 7 | 41 ± 1 | |
| Ondansetron | 61 ± 3 | 34 ± 1 * | |||
| Light | Saline | 14 ± 3 | 22 ± 1 | ||
| Ondansetron | 15 ± 1 | 20 ± 1 | |||
| S-5AA+Met | Dark | Saline | 69 ± 4 | 40 ± 1 | |
| Ondansetron | 73 ± 4 | 34 ± 1 * | |||
| Light | Saline | 15 ± 2 | 21 ± 1 | ||
| Ondansetron | 12 ± 2 | 19 ± 1 | |||
| S-16AA-Met | Dark | Saline | 55 ± 3 | 37 ± 1 | |
| Ondansetron | 69 ± 6 | 31 ± 1 * | |||
| Light | Saline | 16 ± 2 | 21 ± 1 | ||
| Ondansetron | 12 ± 2 | 18 ± 1 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spring, S.; Singh, A.; Zapata, R.C.; Chelikani, P.K.; Pezeshki, A. Methionine Restriction Partly Recapitulates the Sympathetically Mediated Enhanced Energy Expenditure Induced by Total Amino Acid Restriction in Rats. Nutrients 2019, 11, 707. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030707
Spring S, Singh A, Zapata RC, Chelikani PK, Pezeshki A. Methionine Restriction Partly Recapitulates the Sympathetically Mediated Enhanced Energy Expenditure Induced by Total Amino Acid Restriction in Rats. Nutrients. 2019; 11(3):707. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030707
Chicago/Turabian StyleSpring, Shelby, Arashdeep Singh, Rizaldy C. Zapata, Prasanth K. Chelikani, and Adel Pezeshki. 2019. "Methionine Restriction Partly Recapitulates the Sympathetically Mediated Enhanced Energy Expenditure Induced by Total Amino Acid Restriction in Rats" Nutrients 11, no. 3: 707. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030707