Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Samples
2.3. Membrane Chromatography (MC)
2.4. Polymer Precipitation (PP) Using Hexane
2.5. Countercurrent Chromatography (CCC)
2.6. HPLC-PDA-ESI-MS/MS
2.7. HPLC-PDA
2.8. In Vitro α-Amylase Inhibition Study
2.9. In Vitro α-Glucosidase Inhibition Study
3. Results
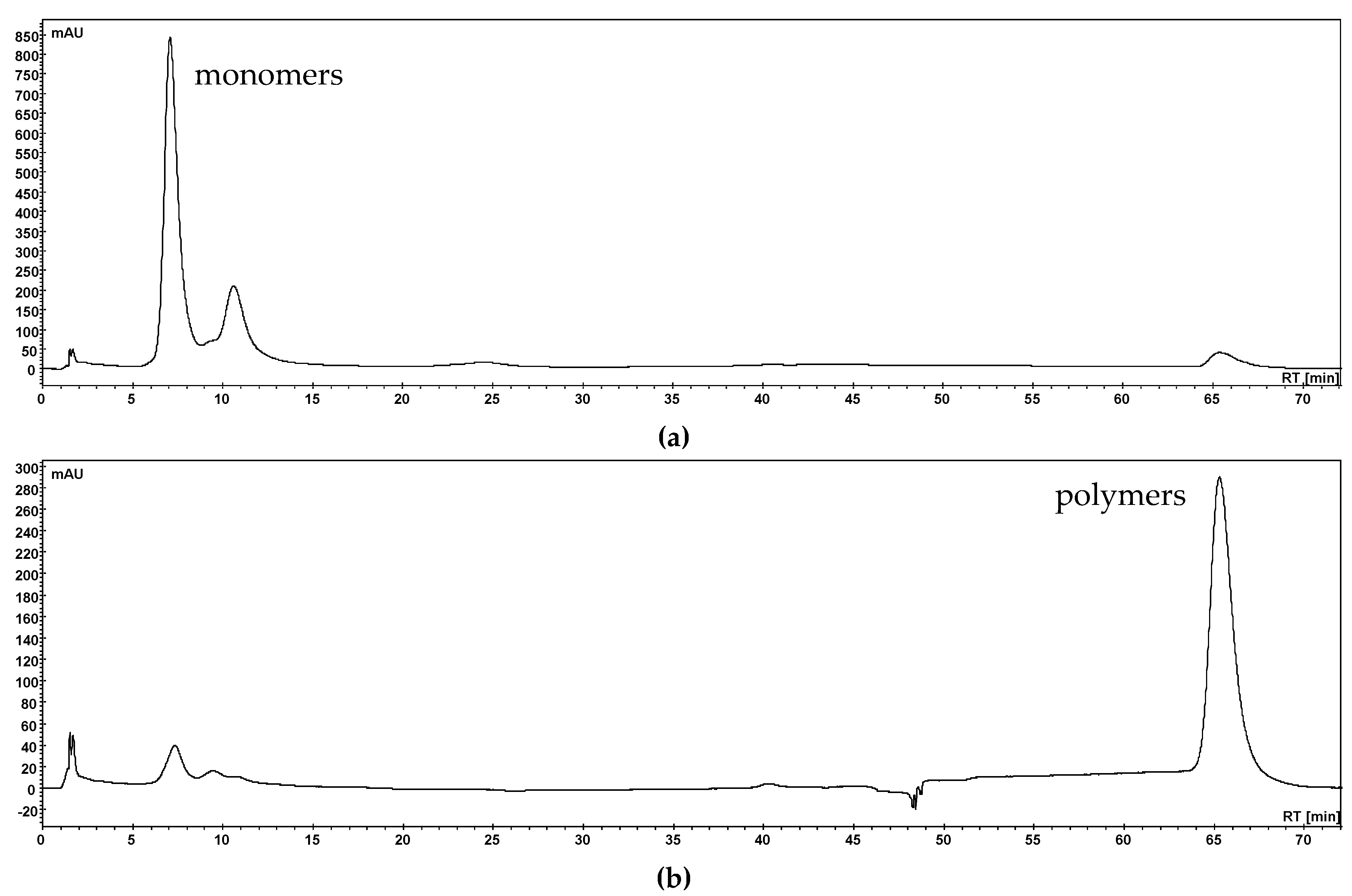
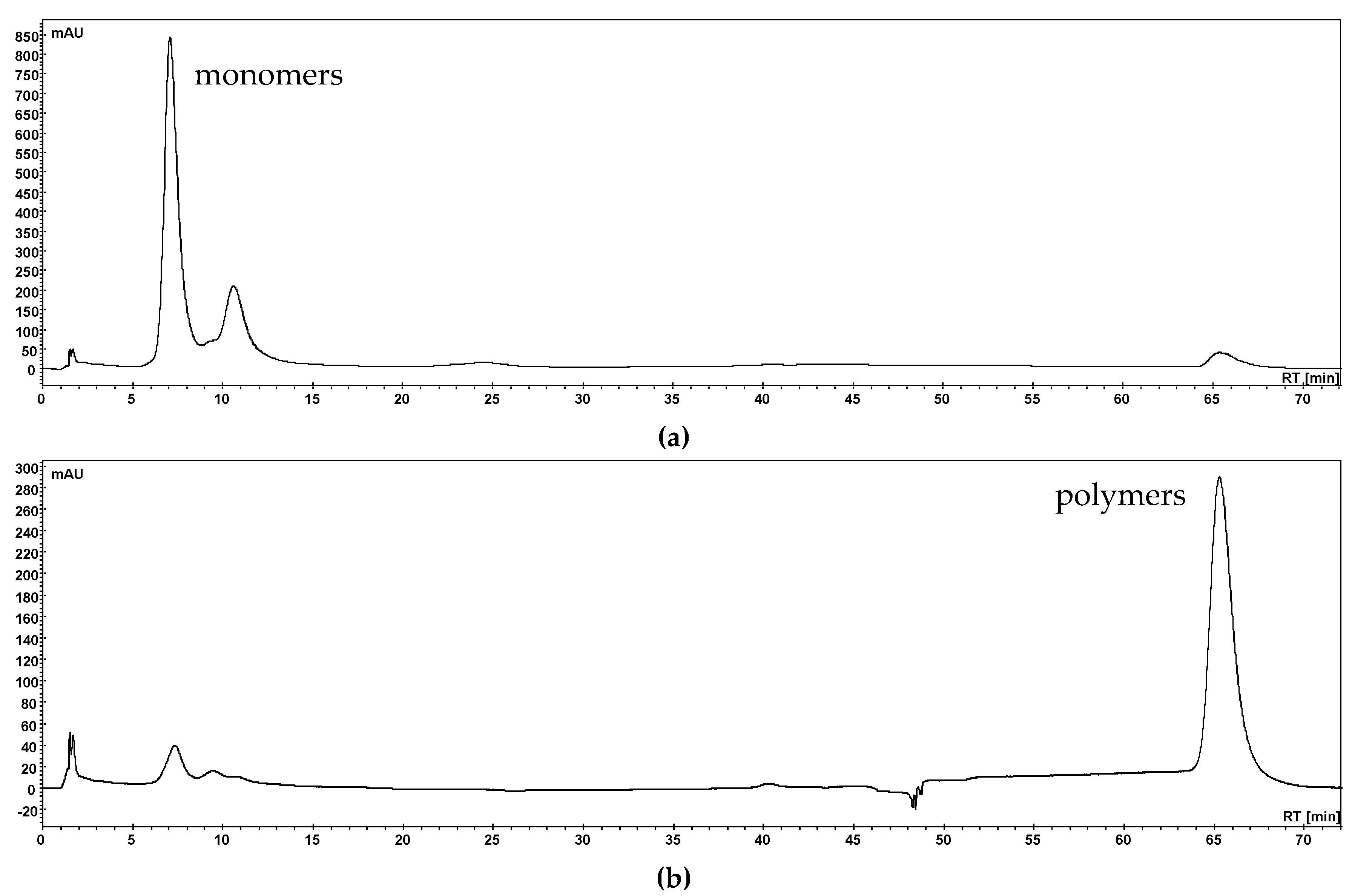
3.1. Separation of Phenolic Compounds into Three Fractions (Anthocyanins, Copigments and Polymers)
3.1.1. Separation with Membrane Chromatography
3.1.2. Polymer Precipitation
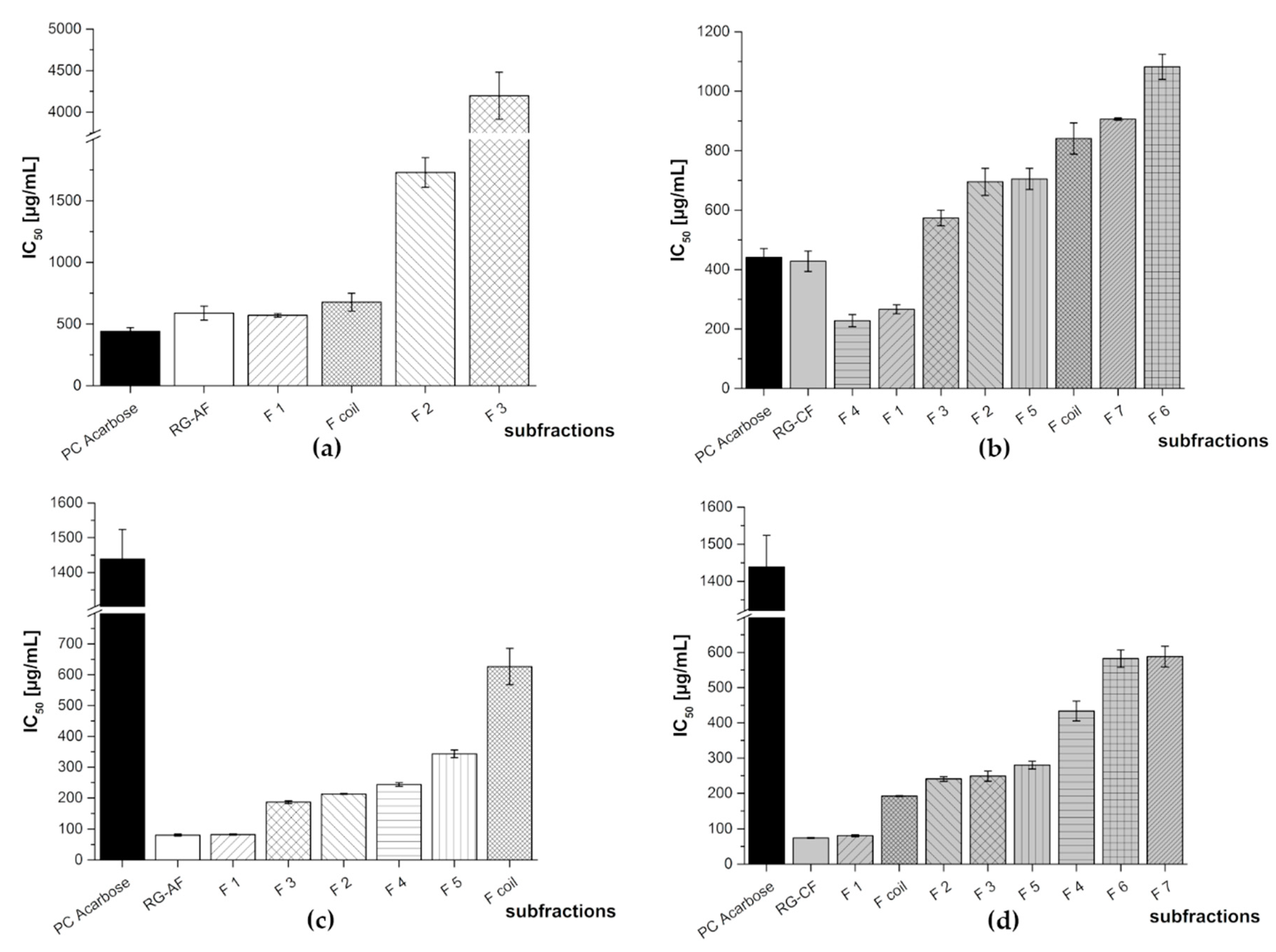
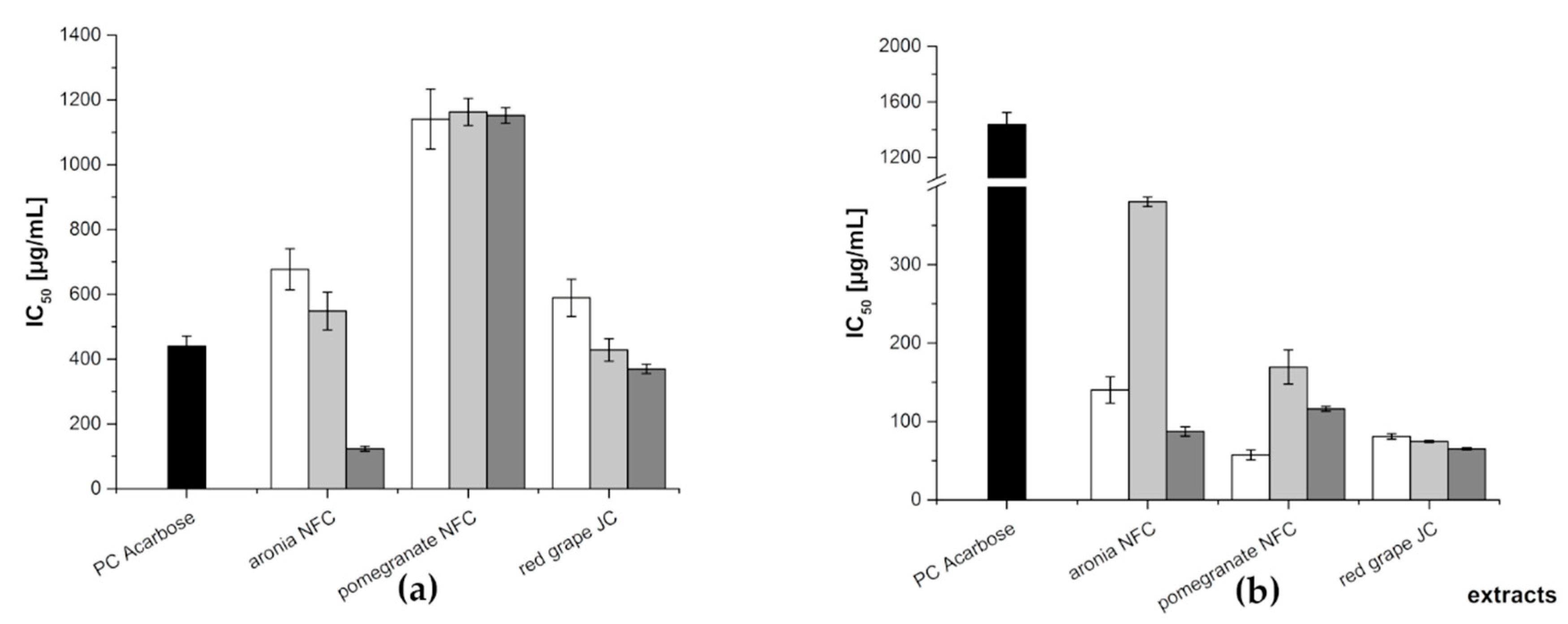
3.1.3. In Vitro Inhibition Study of Anthocyanin, Copigment, and Polymer Fractions of Aronia NFC, Pomegranate NFC and Red Grape JC
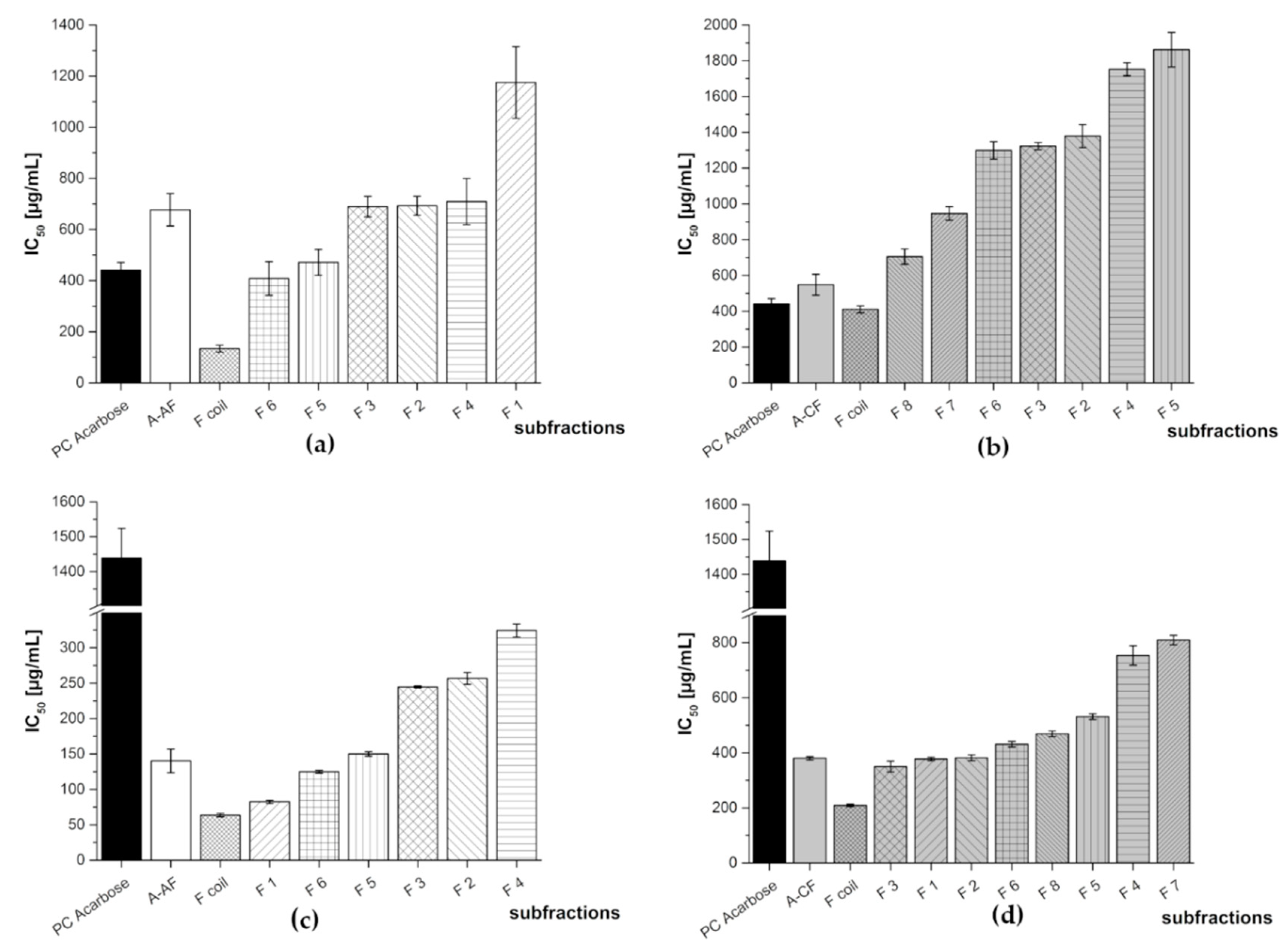
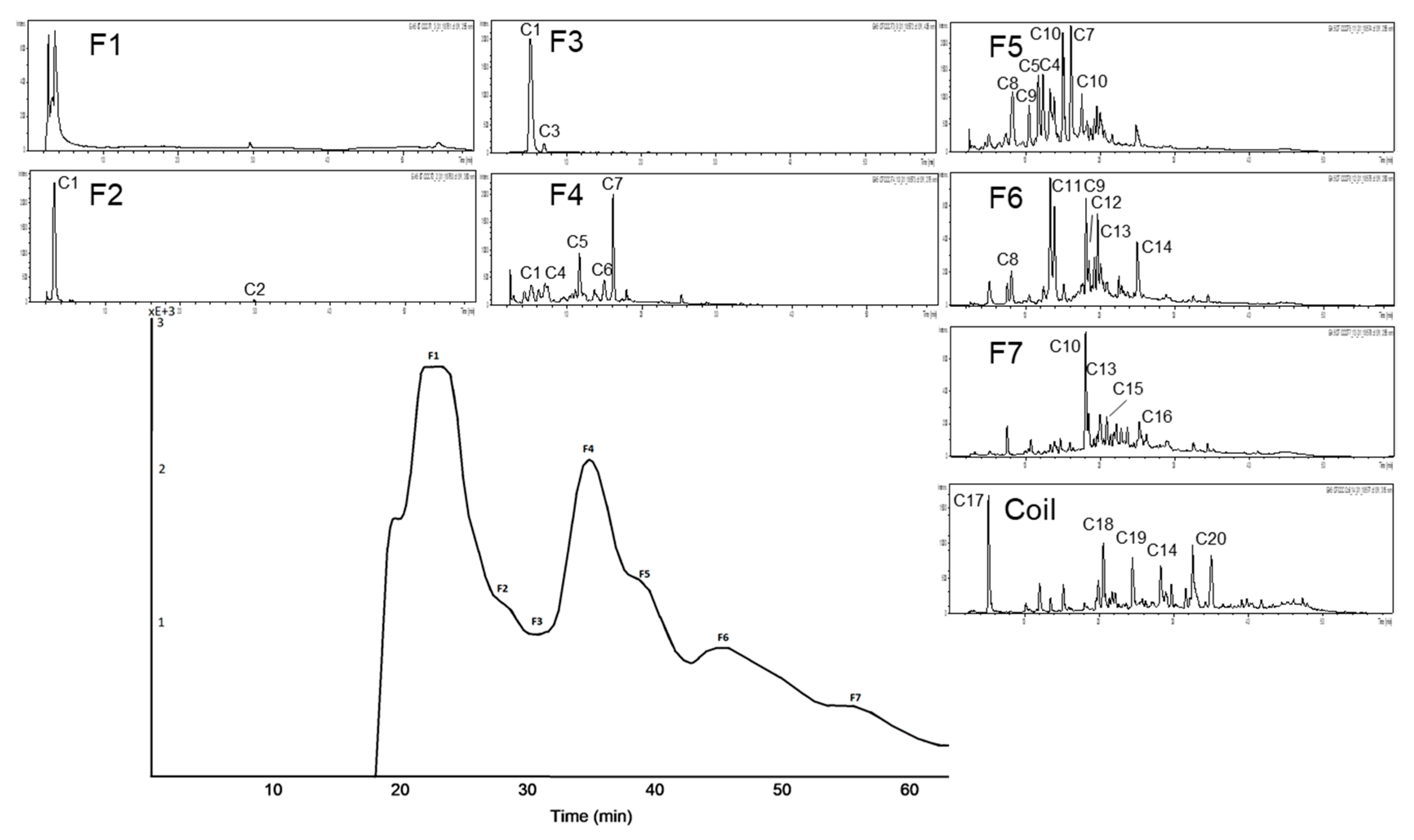
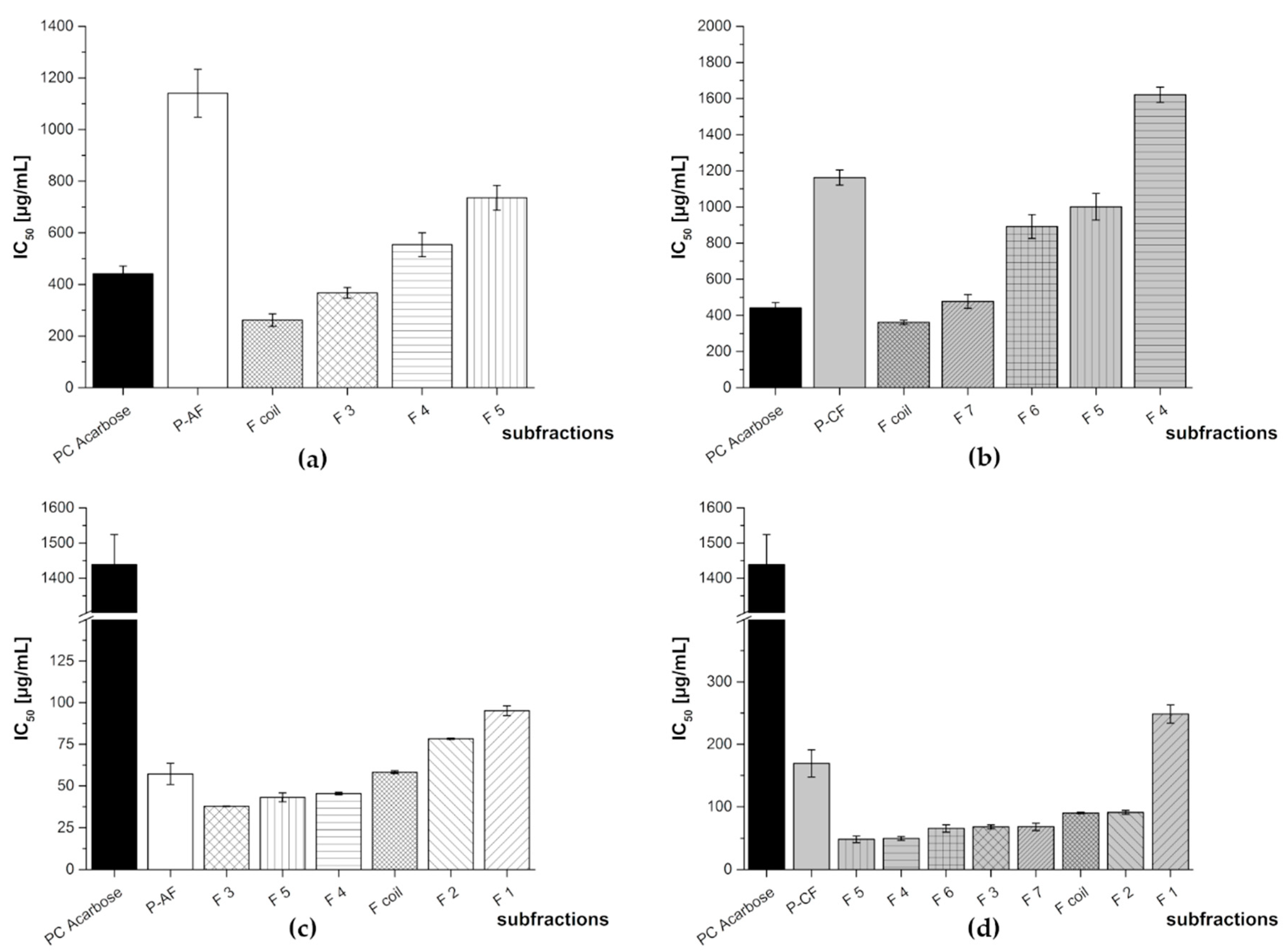
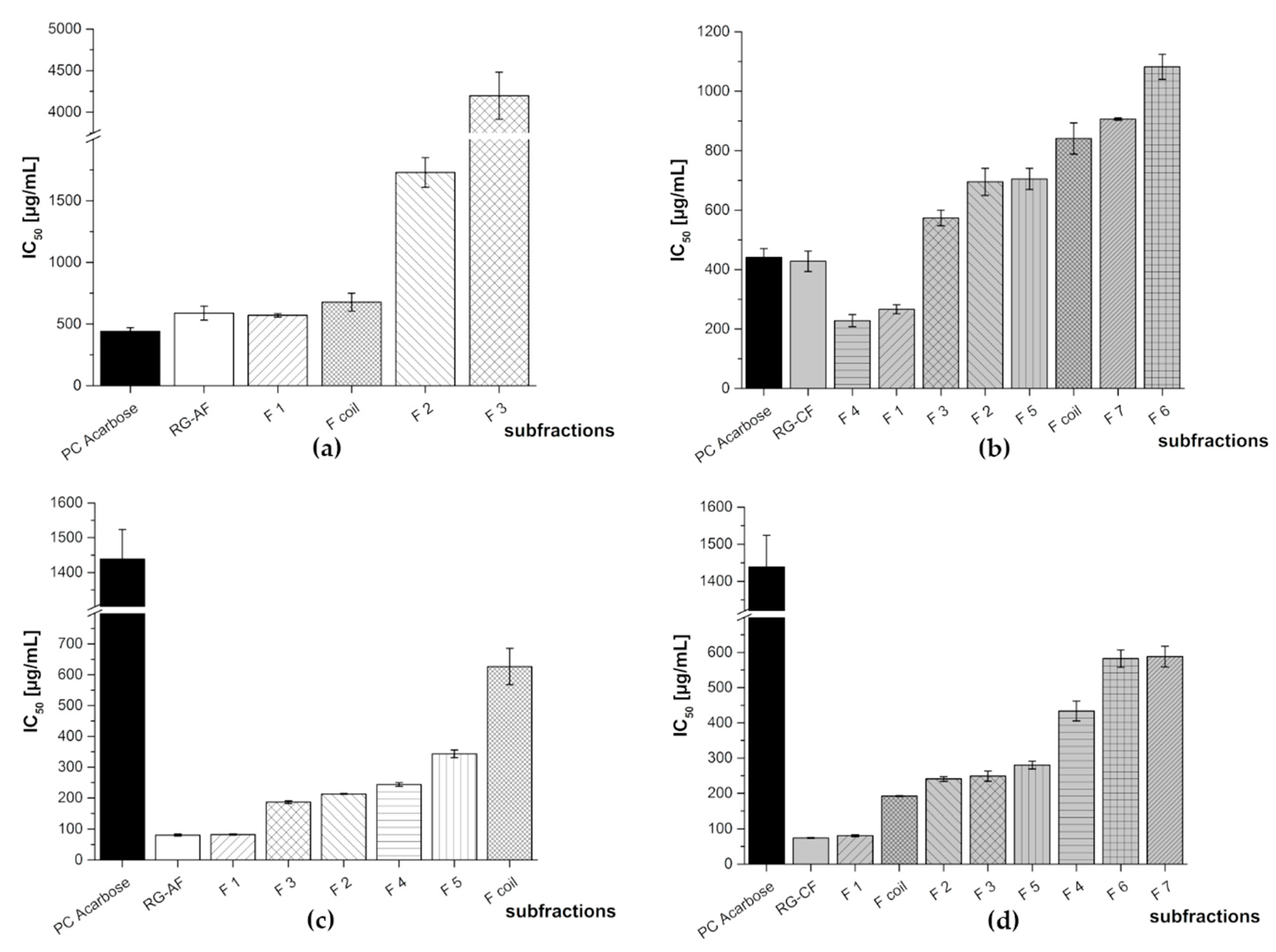
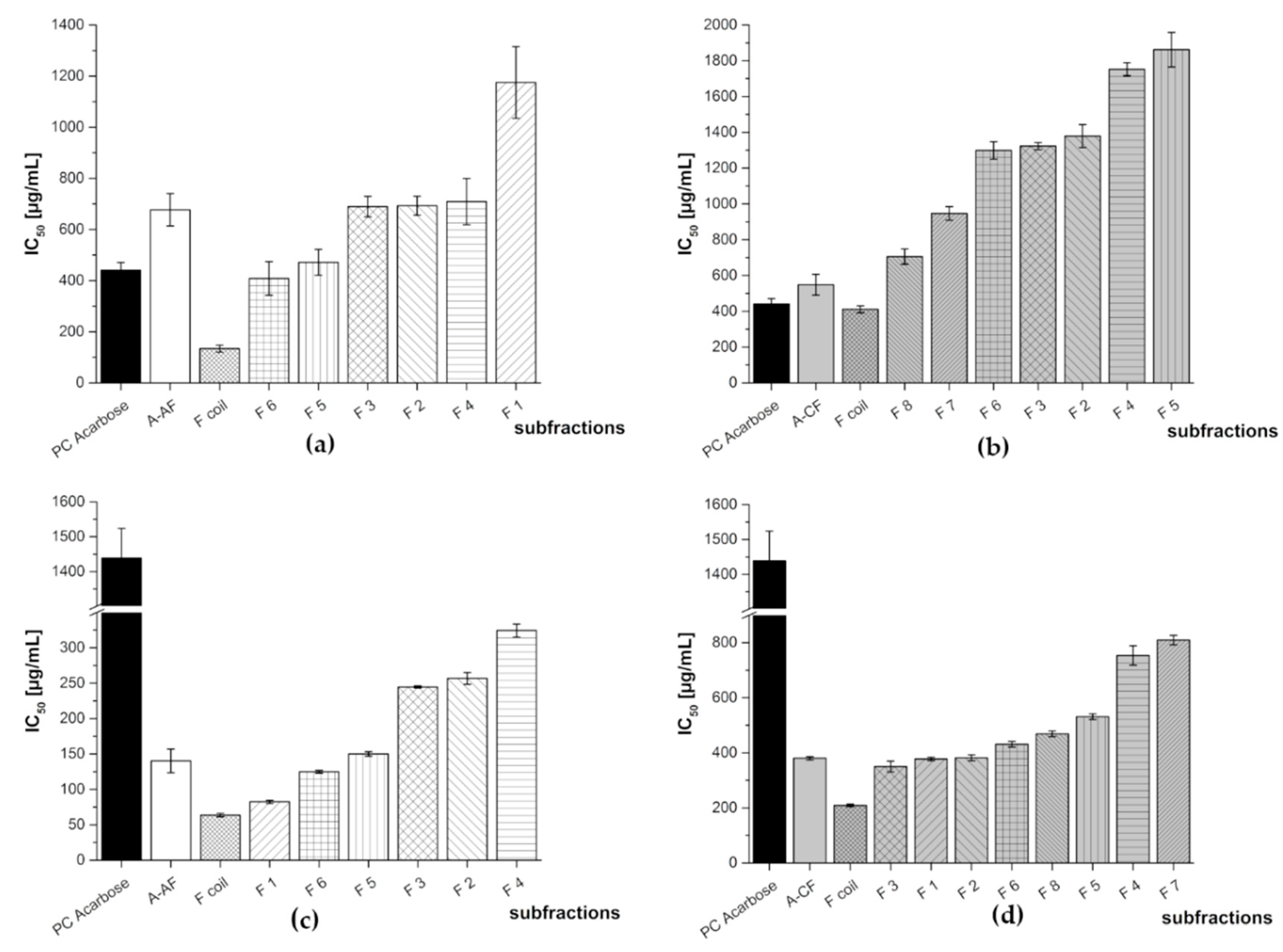
3.2. Subfractionation with Countercurrent Chromatography and In Vitro Inhibition Study
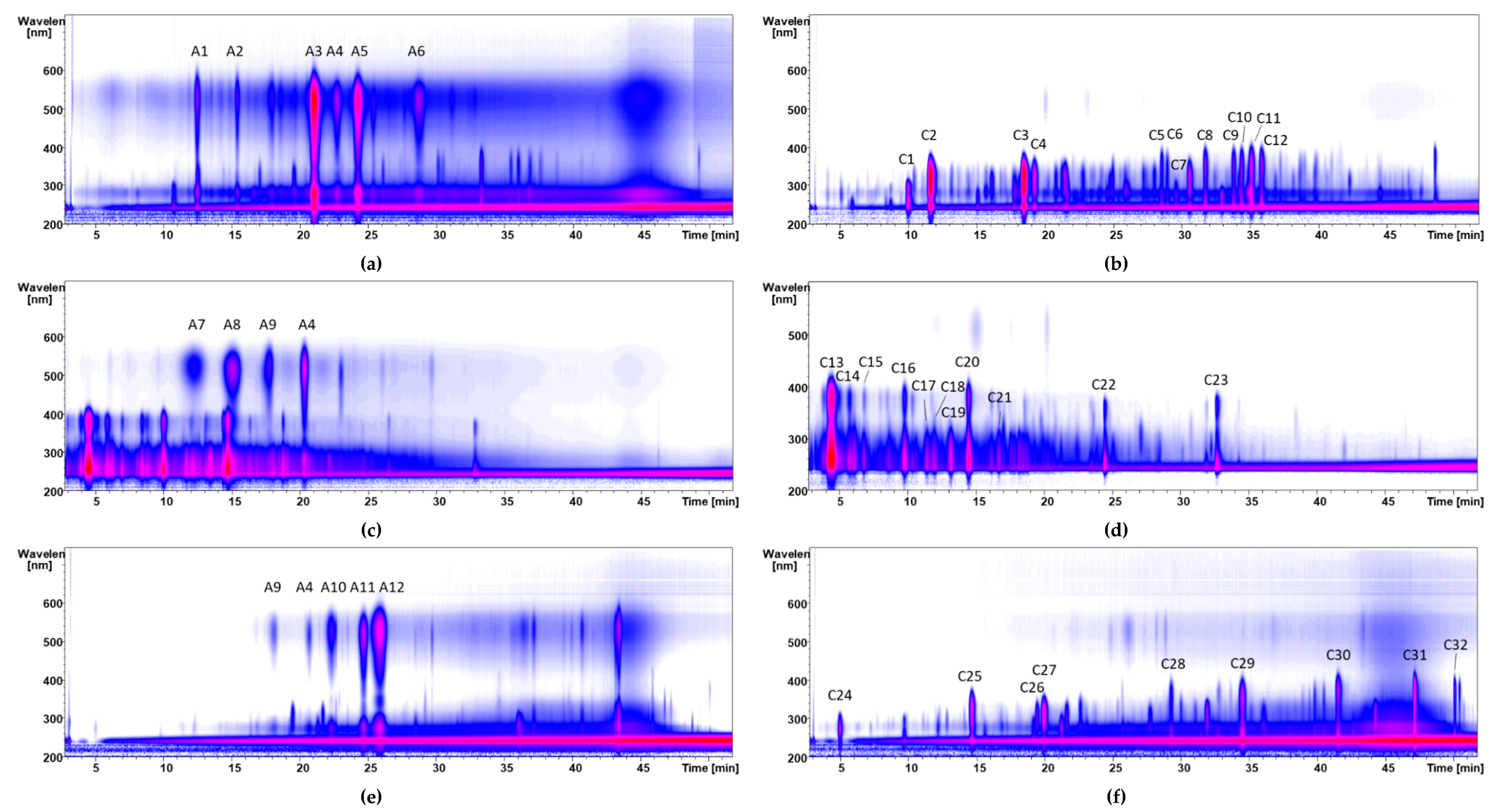
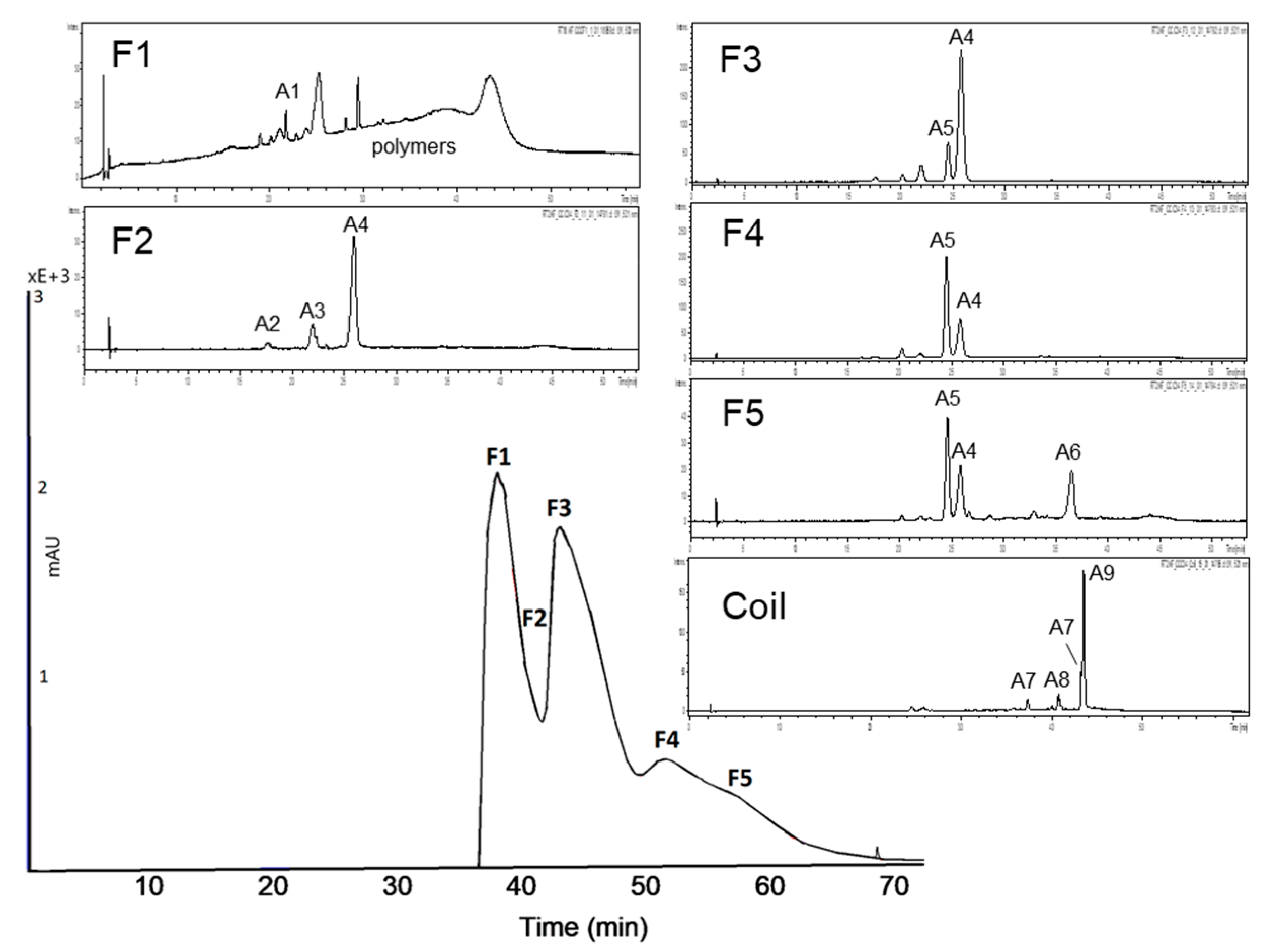
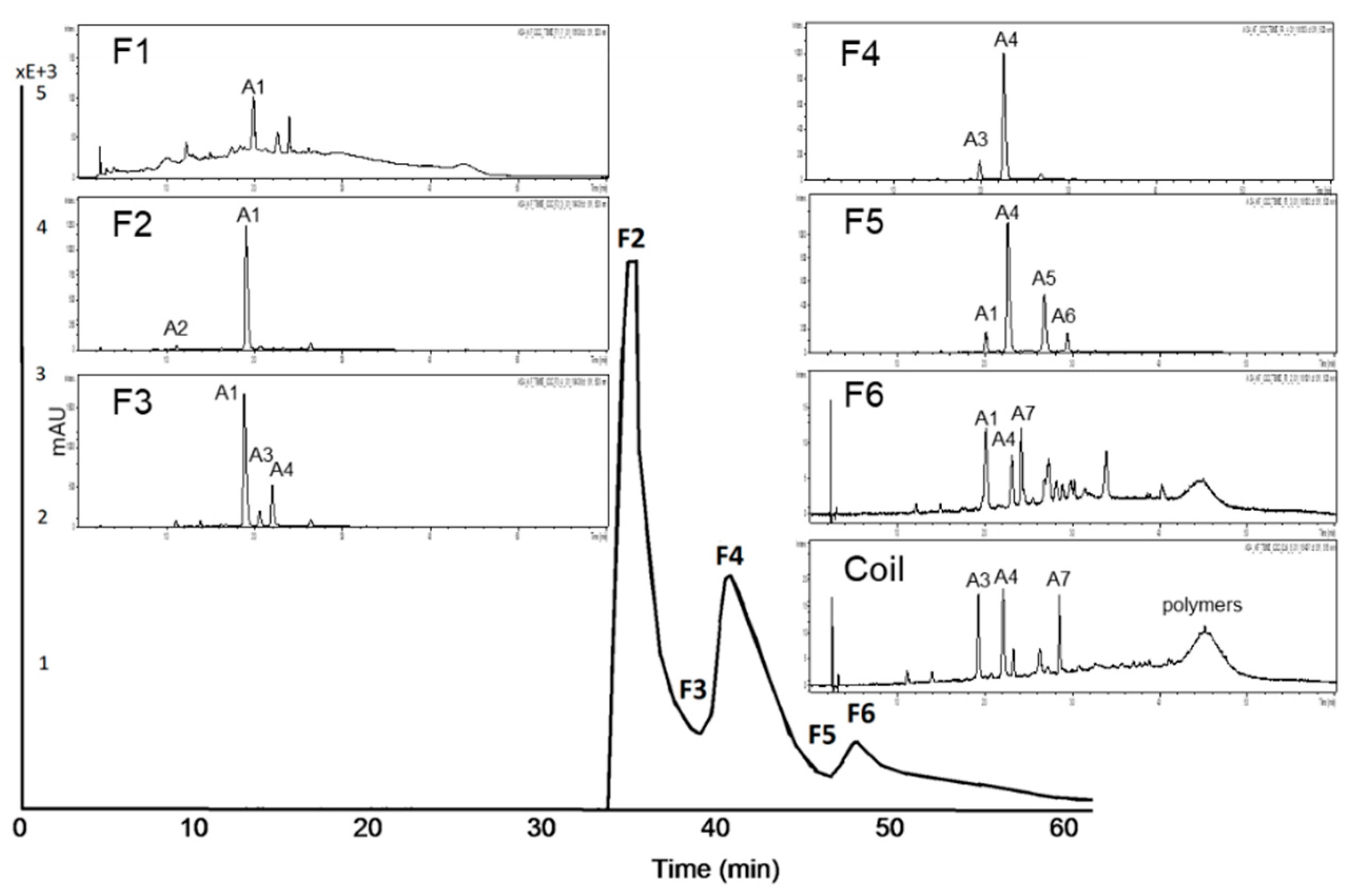
3.2.1. Subfractionation of Anthocyanin and Copigment Fraction of Red Grape JC Using HPCCC
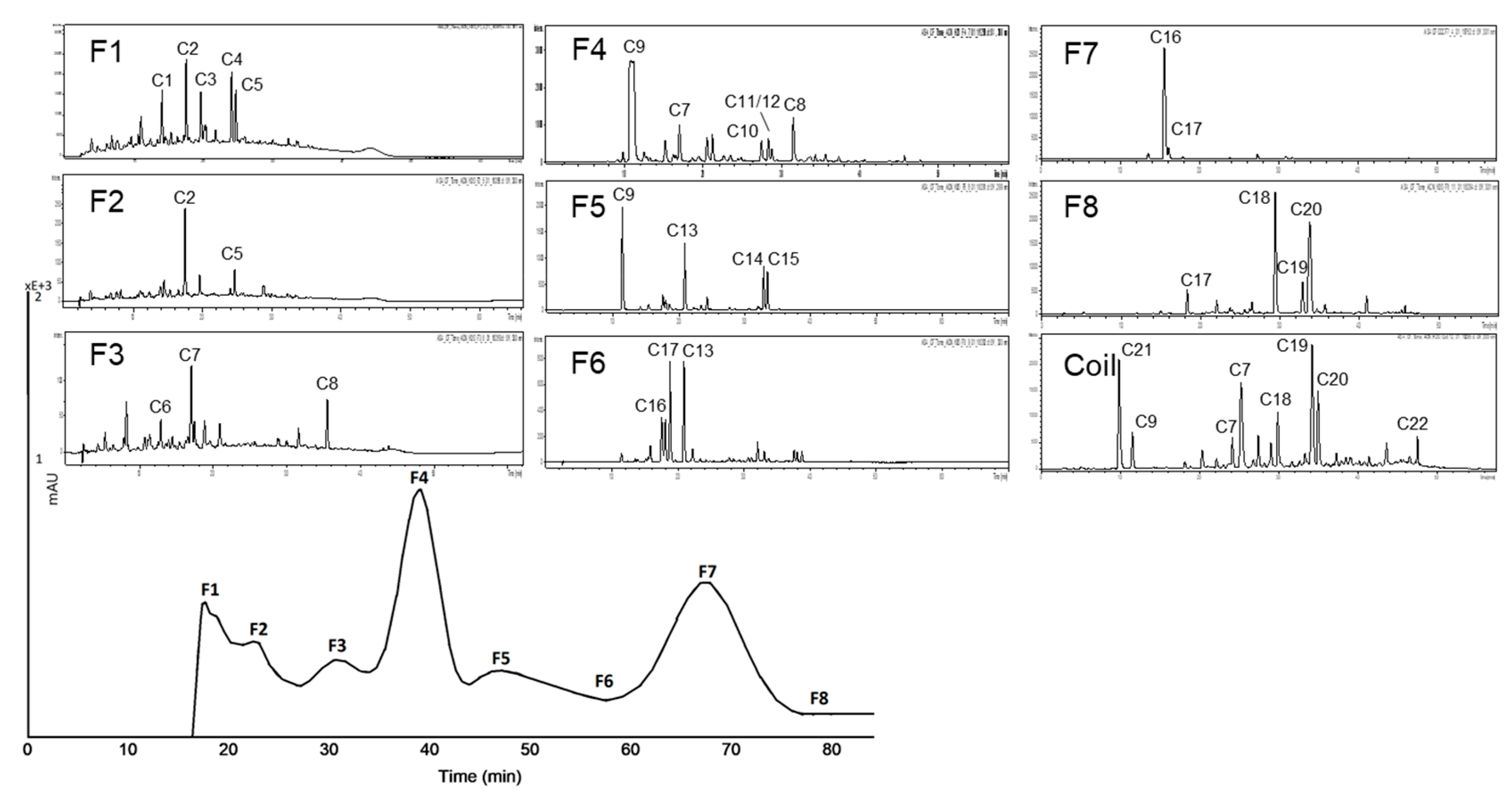
3.2.2. Subfractionation of the Anthocyanin and Copigment Fraction of Aronia NFC Using HPCCC
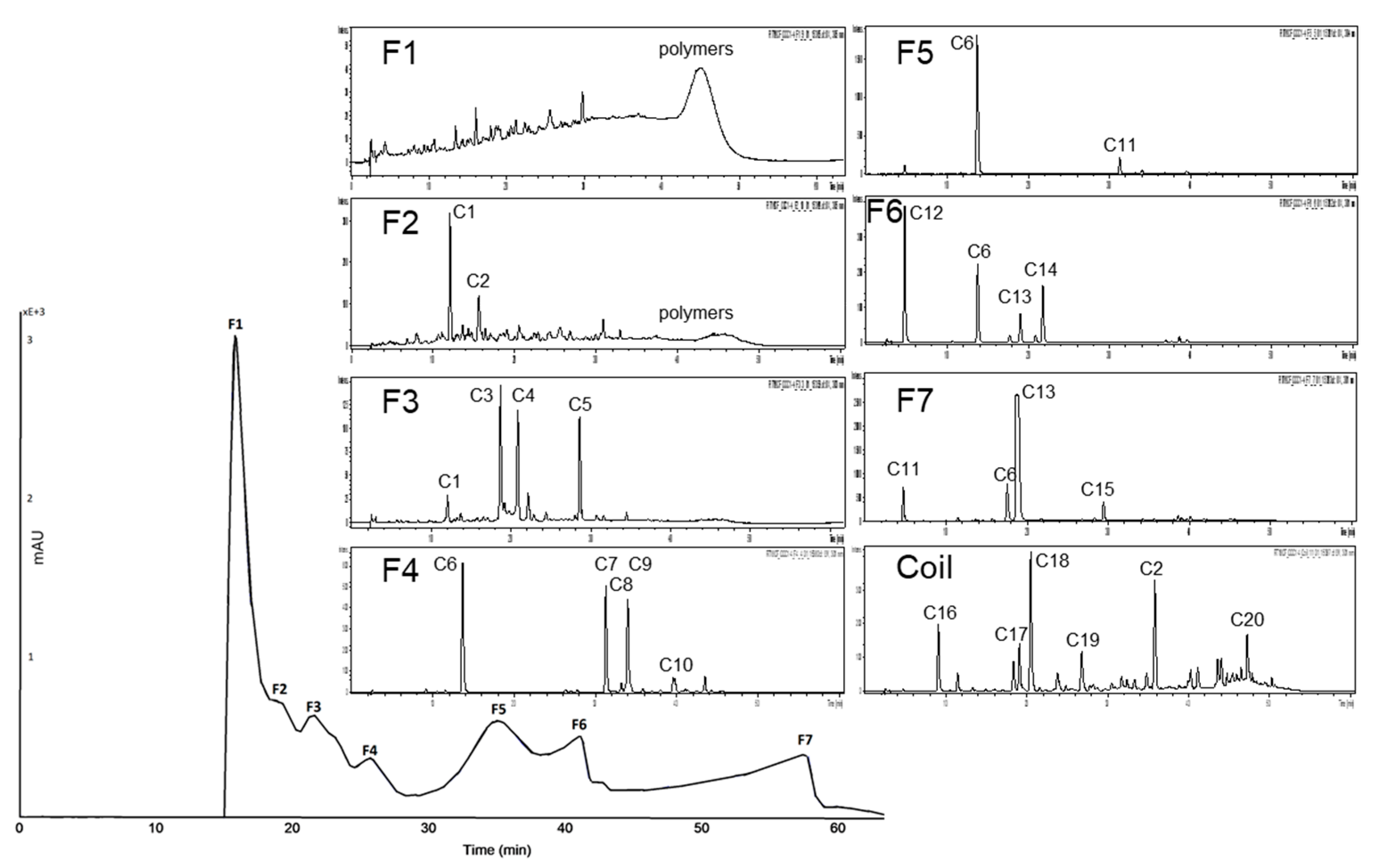
3.2.3. Subfractionation of the Anthocyanin and Copigment Fractions of Pomegranate NFC Using HPCCC
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Becker, S.; Dossus, L.; Kaaks, R. Obesity related hyperinsulinaemia and hyperglycaemia and cancer development. Arch. Physiol. Biochem. 2009, 115, 86–96. [Google Scholar] [CrossRef]
- Levitan, E.B.; Song, Y.; Ford, E.S.; Liu, S. Is nondiabetic hyperglycemia a risk factor for cardiovascular disease? A meta-analysis of prospective studies. Arch. Intern. Med. 2004, 164, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Lo Piparo, E.; Scheib, H.; Frei, N.; Williamson, G.; Grigorov, M.; Chou, C.J. Flavonoids for controlling starch digestion: Structural requirements for inhibiting human alpha-amylase. J. Med. Chem. 2008, 51, 3555–3561. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asia Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, M.K.; Veerapur, V.; Nayak, Y.; Mudgal, P.P.; Mathew, G. Antidiabetic, antihyperlipidemic and antioxidant effects of the flavonoids. In Polyphenols in Human and Diseases; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Elsevier Inc.: London, UK, 2014; Volume 1, pp. 143–161. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of alpha-glucosidase and alpha-amylase by flavonoids. J. Nutr. Sci. Vitaminol. (Tokyo) 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Juadjur, A.; Winterhalter, P. Development of a novel adsorptive membrane chromatographic method for the fractionation of polyphenols from bilberry. J. Agric. Food Chem. 2012, 60, 2427–2433. [Google Scholar] [CrossRef] [PubMed]
- Plate, K.; Beutel, S.; Buchholz, H.; Demmer, W.; Fischer-Frühholz, S.; Reif, O.; Ulber, R.; Scheper, T. Isolation of bovine lactoferrin, lactoperoxidase and enzymatically prepared lactoferricin from proteolytic digestion of bovine lactoferrin using adsorptive membrane chromatography. J. Chromatogr. A 2006, 1117, 81–86. [Google Scholar] [CrossRef]
- Köhler, N.; Wray, V.; Winterhalter, P. Preparative isolation of procyanidins from grape seed extracts by high-speed counter-current chromatography. J. Chromatogr. A 2008, 1177, 114–125. [Google Scholar] [CrossRef]
- Saucier, C.; Mirabel, M.; Daviaud, F.; Longieras, A.; Glories, Y. Rapid fractionation of grape seed proanthocyanidins. J. Agric. Food Chem. 2001, 49, 5732–5735. [Google Scholar] [CrossRef]
- Winterhalter, P. Application of countercurrent chromatography (CCC) to the analysis of natural pigments. Trends Food Sci. Technol. 2007, 18, 507–513. [Google Scholar] [CrossRef]
- De Beer, D.; Malherbe, C.J.; Beelders, T.; Willenburg, E.L.; Brand, D.J.; Joubert, E. Isolation of aspalathin and nothofagin from rooibos (Aspalathus linearis) using high-performance countercurrent chromatography: Sample loading and compound stability considerations. J. Chromatogr. A 2015, 1381, 29–36. [Google Scholar] [CrossRef]
- Sutherland, I.; Thickitt, C.; Douillet, N.; Freebairn, K.; Johns, D.; Mountain, C.; Wood, P.; Edwards, N.; Rooke, D.; Harris, G.; et al. Scalable Technology for the Extraction of Pharmaceutics: Outcomes from a 3 year collaborative industry/academia research programme. J. Chromatogr. A 2013, 1282, 84–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhardt, A.; Knapp, H.; Winterhalter, P. Separation and purification of anthocyanins by High-Speed countercurrent chromatography and screening for antioxidant activity. J. Agric. Food Chem. 2000, 48, 338–343. [Google Scholar] [CrossRef]
- Ashok Kumar, B.S.; Lakshman, K.; Nandeesh, R.; Arun Kumar, P.A.; Manoj, B.; Kumar, V.; Sheshadri Shekar, D. In vitro alpha-amylase inhibition and in vivo antioxidant potential of Amaranthus spinosus in alloxan-induced oxidative stress in diabetic rats. Saudi J. Biol. Sci. 2011, 18, 1–5. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Chen, F.; Wang, X.; Luo, P.G.; Jiang, Y. Inhibitory effects of muscadine anthocyanins on α-glucosidase and pancreatic lipase activities. J. Agric. Food Chem. 2011, 59, 9506–9511. [Google Scholar] [CrossRef] [PubMed]
- Cuevas Montilla, E.; Hillebrand, S.; Antezana, A.; Winterhalter, P. Soluble and bound phenolic compounds in different bolivian purple corn (Zea mays L.) cultivars. J. Agric. Food Chem. 2011, 59, 7068–7074. [Google Scholar] [CrossRef]
- Oszmiański, J.; Lachowicz, S. Effect of the production of dried fruits and juice from chokeberry (Aronia melanocarpa L.) on the content and antioxidative activity of bioactive compounds. Molecules 2016, 21, 1098. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Hillebrand, S.; Winterhalter, P. Analytik von phenolischen Inhaltsstoffen in Aroniasäften (Aronia melanocarpa). Dtsch. Lebensm. Rundsch. 2010, 106, 374–382. [Google Scholar]
- Kulling, S.E.; Rawel, H.M. Chokeberry (Aronia melanocarpa)—A review on the characteristic components and potential health effects. Planta Med. 2008, 74, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD-ESI/MS(n). Food Chem. 2011, 127, 807–821. [Google Scholar] [CrossRef]
- Kammerer, D.; Claus, A.; Carle, R.; Schieber, A. Polyphenol screening of pomace from red and white grape varieties (Vitis vinifera L.) by HPLC-DAD-MS/MS. J. Agric. Food Chem. 2004, 52, 4360–4367. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Mardones, C.; Hermosín-Gutiérrez, I.; Von Baer, D. Comparison of high-performance liquid chromatography separation of red wine anthocyanins on a mixed-mode ion-exchange reversed-phase and on a reversed-phase column. J. Agric. Food Chem. 2010, 1217, 5710–5717. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sutherland, I.A. How to achieve rapid separations in counter-current chromatography. J. Agric. Food Chem. 2006, 1114, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.; Ndou, T.; Hughey, C.A.; Straut, C.; Howell, A.; Dai, Z.; Kaletunc, G. Inhibition of α-amylase and glucoamylase by tannins extracted from cocoa, pomegranates, cranberries, and grapes. J. Agric. Food Chem. 2013, 61, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Kato, C.G.; Gonçalves, G.A.; Peralta, R.A.; Seixas, F.A.V.; de Sá-Nakanishi, A.B.; Bracht, L.; Comar, J.F.; Bracht, A.; Peralta, R.M. Inhibition of α-Amylases by Condensed and Hydrolysable Tannins: Focus on Kinetics and Hypoglycemic Actions. Enzyme Res. 2017, 2017, 5724902. [Google Scholar] [CrossRef] [PubMed]
- Akkarachiyasit, S.; Charoenlertkul, P.; Yibchok-anun, S.; Adisakwattana, S. Inhibitory activities of cyanidin and its glucosides and synergistic effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Int. J. Mol. Sci. 2010, 11, 3387–3396. [Google Scholar] [CrossRef]
- Cheplick, S.; Kwon, Y.-I.; Bhowmik, P.; Shetty, K. Clonal variation in raspberry fruit phenolics and revelance for diabetes and hypertension management. J. Food Biochem. 2007, 31, 656–679. [Google Scholar] [CrossRef]
- Ueno, H.; Tsuchimochi, W.; Wang, H.-W.; Yamashita, E.; Tsubouchi, C.; Nagamine, K.; Sakoda, H.; Nakazato, M. Effects of miglitol, acarbose, and sitagliptin on plasma insulin and gut peptides in Type 2 diabetes mellitus: A crossover study. Diabetes Ther. 2015, 6, 187–196. [Google Scholar] [CrossRef] [PubMed]


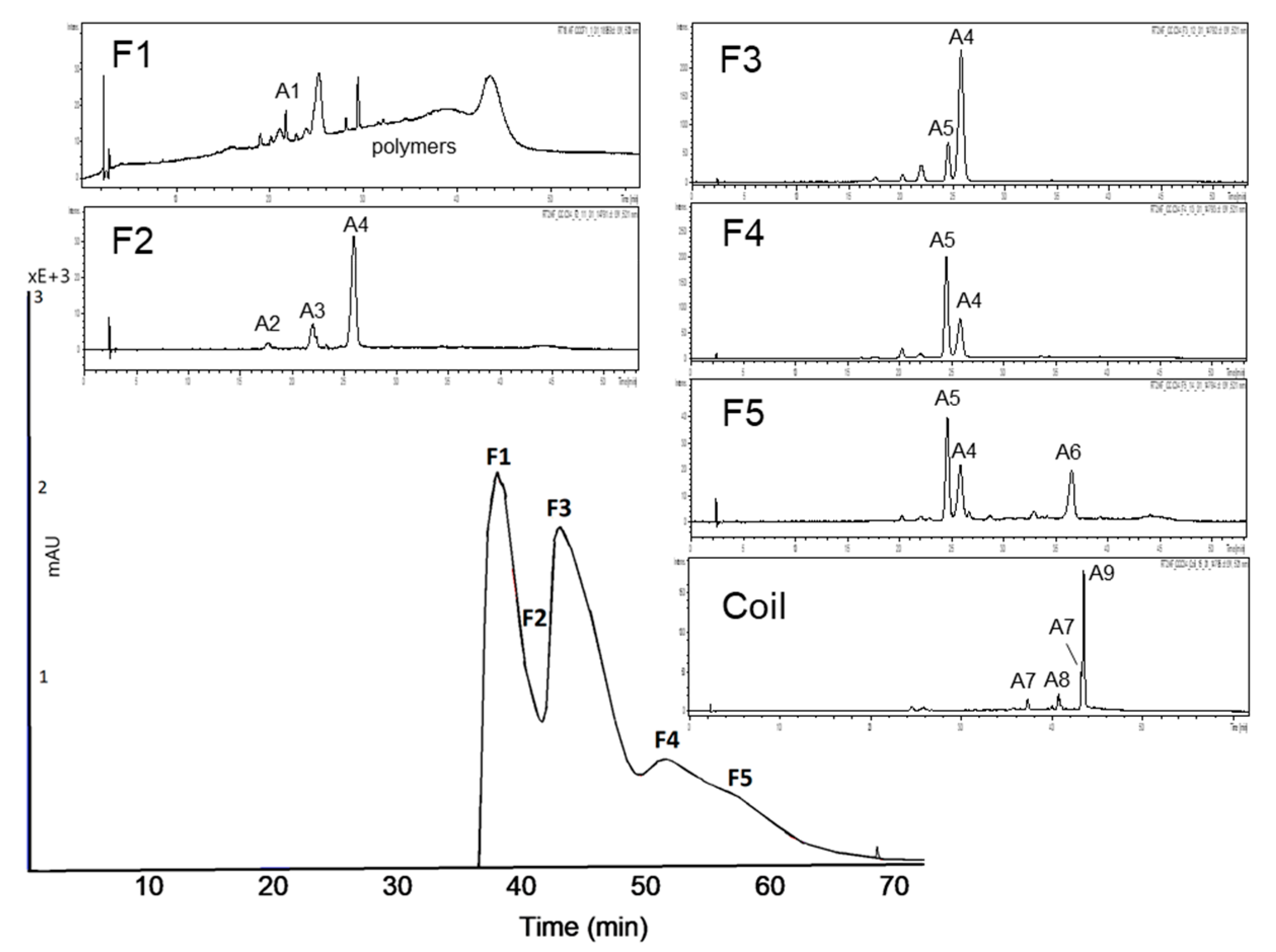
| Peak | Compound | Peak | Compound |
| A1 | delphinidin-3-glucoside | A6 | malvidin-3-acetylglucoside |
| A2 | cyanidin-3-glucoside | A7 | peonidin-3-coumaroylhexoside I+II |
| A3 | petunidin-3-glucoside | A8 | petunidin-3-coumaroylhexoside |
| A4 | malvidin-3-glucoside | A9 | malvidin-3-coumaroylhexoside |
| A5 | peonidin-3-glucoside |

| Peak | Compound | Peak | Compound | Peak | Compound |
| C1 | coumaroyl derivative | C8 | quercetin-3-galactoside | C15 | unknown |
| C2 | unknown | C9 | quercetin-3-glucoside | C16 | protocatechuic acid |
| C3 | coumaroyl-hexoside I | C10 | quercetin derivative | C17 | resveratrol |
| C4 | coumaroyl-hexoside II | C11 | quercetin-3-glucuronide | C18 | coumaric acid |
| C5 | isorhamnetin-3-hexoside | C12 | gallic acid | C19 | syringic acid |
| C6 | caftaric acid | C13 | coutaric acid | C20 | epicatechin |
| C7 | resveratrol-3-glucoside (piceid) | C14 | fertaric acid |
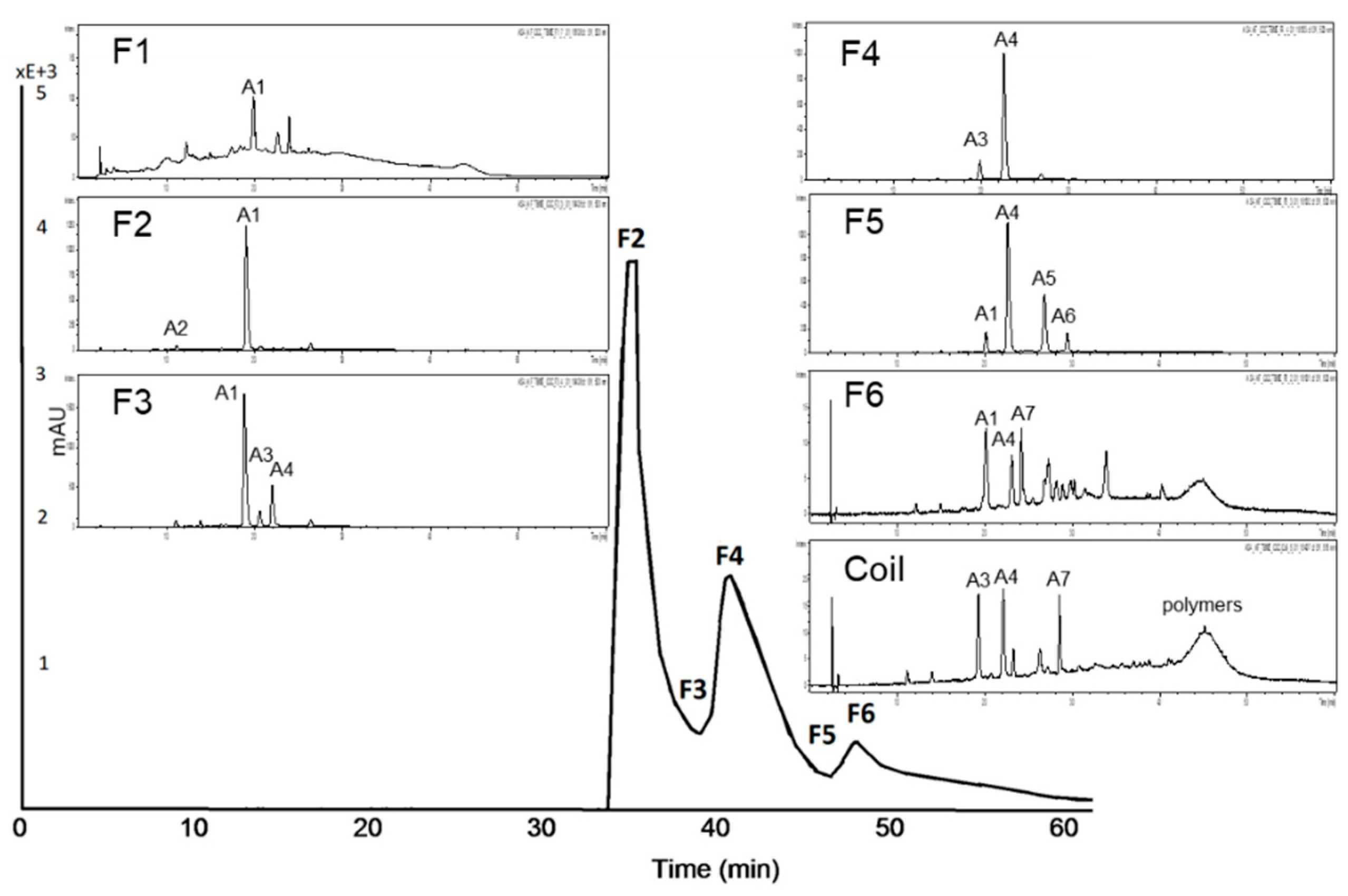
| Peak | Compound | Peak | Compound |
| A1 | cyanidin-3-galactoside | A5 | cyanidin-3-xyloside |
| A2 | cyanidin derivative | A6 | cyanidin derivative |
| A3 | cyanidin-3-glucoside | A7 | unknown |
| A4 | cyanidin-3-arabinoside |

| Peak | Compound | Peak | Compound | Peak | Compound |
| C1 | dicaffeoylquinic ester I | C9 | neochlorogenic acid (3-CQA) | C17 | cryptochlorogenic acid (4-CQA) |
| C2 | dicaffeoylquinic ester II | C10 | coumaroyl quinic ester | C18 | feruloylquinic acid II |
| C3 | quercetin derivative | C11 | quercetin-dihexoside I | C19 | quercetin-pentoside |
| C4 | caffeic acid-derivative | C12 | quercetin-dihexoside II | C20 | quercetin-hexoside |
| C5 | unknown | C13 | feruloylquinic acid I | C21 | protocatechuic acid |
| C6 | eriodictoyl-7-glucoronide | C14 | quercetin-3-robinobioside | C22 | quercetin |
| C7 | coumaroyl derivative I + II | C15 | quercetin-3-rutinoside | ||
| C8 | quercetin-3-vivianoside | C16 | chlorogenic acid (5-CQA) |

| Peak | Compound | Peak | Compound |
| A1 | delphindin-3,5-diglucoside | A5 | delphinidin-3-glucoside |
| A2 | cyanidin-3,5-diglucoside | A6 | pelargonidin-glucoside |
| A3 | cyanidin-3-glucoside | A7 | cyanidin-pentoside |
| A4 | pelargonidin-3,5-diglucoside |
| Peak | Compound | Peak | Compound | Peak | Compound |
| C1 | punicalin | C8 | digalloyl-hexoside | C15 | caffeic acid hexoside |
| C2 | ellagic acid | C9 | granatin B I | C16 | galloyl-HHDP-hexoside-pentoside |
| C3 | punicalagin I | C10 | pedunculagin I+II | C17 | gallic acid |
| C4 | ellagic acid derivatives | C11 | granatin B II+III | C18 | brevifolin carboxylic acid |
| C5 | punicalagin II | C12 | casuarinin | C19 | valoneic acid bilactone |
| C6 | galloyl-HHDP-hexoside | C13 | punigluconin | C20 | ellagic acid deoxyhexoside |
| C7 | punicalagin III | C14 | ellagic acid hexoside |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | [M-H]+ | Fragments (m/z) | Compound | Peak | [M-H]− | Fragments (m/z) | Compound |
|---|---|---|---|---|---|---|---|
| A1 | 737 | 575, 287 | cyanidin derivative | C1 | 153 | 109 | protocatechuic acid |
| A2 | 707 | 575, 287 | cyanidin derivative | C2 | 353 | 191, 179,135 | neochlorogenic acid (3-CQA) |
| A3 | 449 | 287 | cyanidin-3-galactoside | C3 | 353 | 191, 179, 161 | chlorogenic acid (5-CQA) |
| A4 | 449 | 287 | cyanidin-3-glucoside | C4 | 353 | 191, 179, 173, 135 | cryptochlorogenic acid (4-CQA) |
| A5 | 419 | 287 | cyanidin-3-arabinoside | C5 | 625 | 301 | quercetin dihexoside I |
| A6 | 419 | 287 | cyanidin-3-xyloside | C6 | 625 | 301 | quercetin dihexoside II |
| A7 | 627 | 465, 303 | delphinidin-3,5-diglucoside | C7 | 367 | 191, 179, 135 | feruloylquinic acid |
| A8 | 611 | 449, 287 | cyanidin-3,5-diglucoside | C8 | 595 | 301 | quercetin-3-vivianoside |
| A9 | 465 | 303 | delphinidin-3-glucoside | C9 | 609 | 301 | quercetin-3-robinobioside |
| A10 | 479 | 317 | petunidin-3-glucoside | C10 | 609 | 301 | quercetin-3-rutinoside |
| A11 | 463 | 301 | peonidin-3-glucoside | C11 | 463 | 301 | quercetin-3-galactoside |
| A12 | 493 | 331 | malvidin-3-glucoside | C12 | 463 | 301 | quercetin-3-glucoside |
| C13 | 781 | 601, 271 | punicalin | ||||
| C14 | 1083 | 601, 299, 271 | punicalagin I | ||||
| C15 | 783 | 601, 301 | pedunculagin I | ||||
| C16 | 1083 | 601, 299, 271 | punicalagin II | ||||
| C17 | 783 | 601, 301 | pedunculagin II | ||||
| C18 | 951 | 907, 301 | granatin B | ||||
| C19 | 783 | 601, 301 | pedunculagin III | ||||
| C20 | 1083 | 601, 301 | punicalagin III | ||||
| C21 | 799 | 479, 301 | ellagic acid-derivatives | ||||
| C22 | 463 | 301, 257, 163 | ellagic acid-hexoside | ||||
| C23 | 301 | 258 | ellagic acid | ||||
| C24 | 169 | 125 | gallic acid | ||||
| C25 | 311 | 179, 149 | caftaric acid | ||||
| C26 | 325 | 175, 163 | coutaric acid-hexoside | ||||
| C27 | 295 | 163 | isorhamnetin-3-glucoside | ||||
| C28 | 477 | 316 | quercetin-hexoside | ||||
| C29 | 477 | 301 | quercetin-glucuronide | ||||
| C30 | 317 | 179 | isorhamnetin | ||||
| C31 | 301 | - | quercetin | ||||
| C32 | 285 | 151 | kaempferol |
| Extract | Fraction | IC50 [µg/mL] | |
|---|---|---|---|
| α-Amylase Assay | α-Glucosidase Assay | ||
| PC Acarbose | 440 ± 30 | 1439 + 85 | |
| aronia NFC | AF | 677 ± 63 | 140 ± 17 |
| CF | 548 ± 58 | 380 ± 6 | |
| PF | 123 ± 8 | 87.2 ± 5.9 | |
| pomegranate NFC | AF | 1141 ± 93 | 57.2 ± 6.4 |
| CF | 1163 ± 42 | 169 ± 22 | |
| PF | 1152 ± 24 | 116 ± 3 | |
| red grape JC | AF | 589 ± 57 | 80.9 ± 3.6 |
| CF | 428 ± 35 | 74.5 ± 1.4 | |
| PF | 369 ± 14 | 65.0 ± 1.4 | |
| Red Grape JC Fraction | Subfraction | IC50 [µg/mL] | |
|---|---|---|---|
| α-Amylase Assay | α-Glucosidase Assay | ||
| PC Acarbose | 440 ± 30 | 1439 + 85 | |
| RG-AF | F 1 | 571 ± 14 | 82.8 ± 1.9 |
| F 2 | 1730 ± 120 | 213 ± 1 | |
| F 3 | 4198 ± 283 | 187 ± 5 | |
| F 4 | N/A | 244 ± 6 | |
| F 5 | N/A | 343 ± 12 | |
| F coil | 677 ± 72 | 626 ± 59 | |
| RG-CF | F 1 | 266 ± 15 | 80.7 ± 2.8 |
| F 2 | 695 ± 45 | 241 ± 7 | |
| F 3 | 574 ± 26 | 249 ± 14 | |
| F 4 | 228 ± 20 | 433 ± 28 | |
| F 5 | 705 ± 36 | 280 ± 11 | |
| F6 | 1082 ± 42 | 582 ± 25 | |
| F 7 | 907 ± 4 | 588 ± 29 | |
| F coil | 841 ± 52 | 192 ± 1 | |
| Aronia NFC Fraction | Subfraction | IC50 [µg/mL] | |
|---|---|---|---|
| α-Amylase Assay | α-Glucosidase Assay | ||
| PC Acarbose | 440 ± 30 | 1439 + 85 | |
| A-AF | F 1 | 1175 ± 140 | 82.4 ± 2.2 |
| F 2 | 693 ± 37 | 257 ± 8 | |
| F 3 | 690 ± 40 | 245 ± 1 | |
| F 4 | 709 ± 91 | 324 ± 9 | |
| F 5 | 471 ± 50 | 150 ± 3 | |
| F 6 | 408 ± 66 | 125 ± 2 | |
| F coil | 134 ± 14 | 63.7 ± 2.4 | |
| A-CF | F 1 | N/A | 377 ± 6 |
| F 2 | 1379 ± 64 | 382 ± 11 | |
| F 3 | 1323 ± 20 | 350 ± 19 | |
| F 4 | 1753 ± 36 | 753 ± 35 | |
| F 5 | 1862 ± 96 | 531 ± 10 | |
| F6 | 1299 ± 49 | 431 ± 10 | |
| F 7 | 947 ± 38 | 809 ± 17 | |
| F 8 | 706 ± 42 | 469 ± 11 | |
| F coil | 411 ± 20 | 209 ± 5 | |
| Pomegranate NFC Fraction | Subfraction | IC50 [µg/mL] | |
|---|---|---|---|
| α-Amylase Assay | α-Glucosidase Assay | ||
| PC Acarbose | 440 ± 30 | 1439 + 85 | |
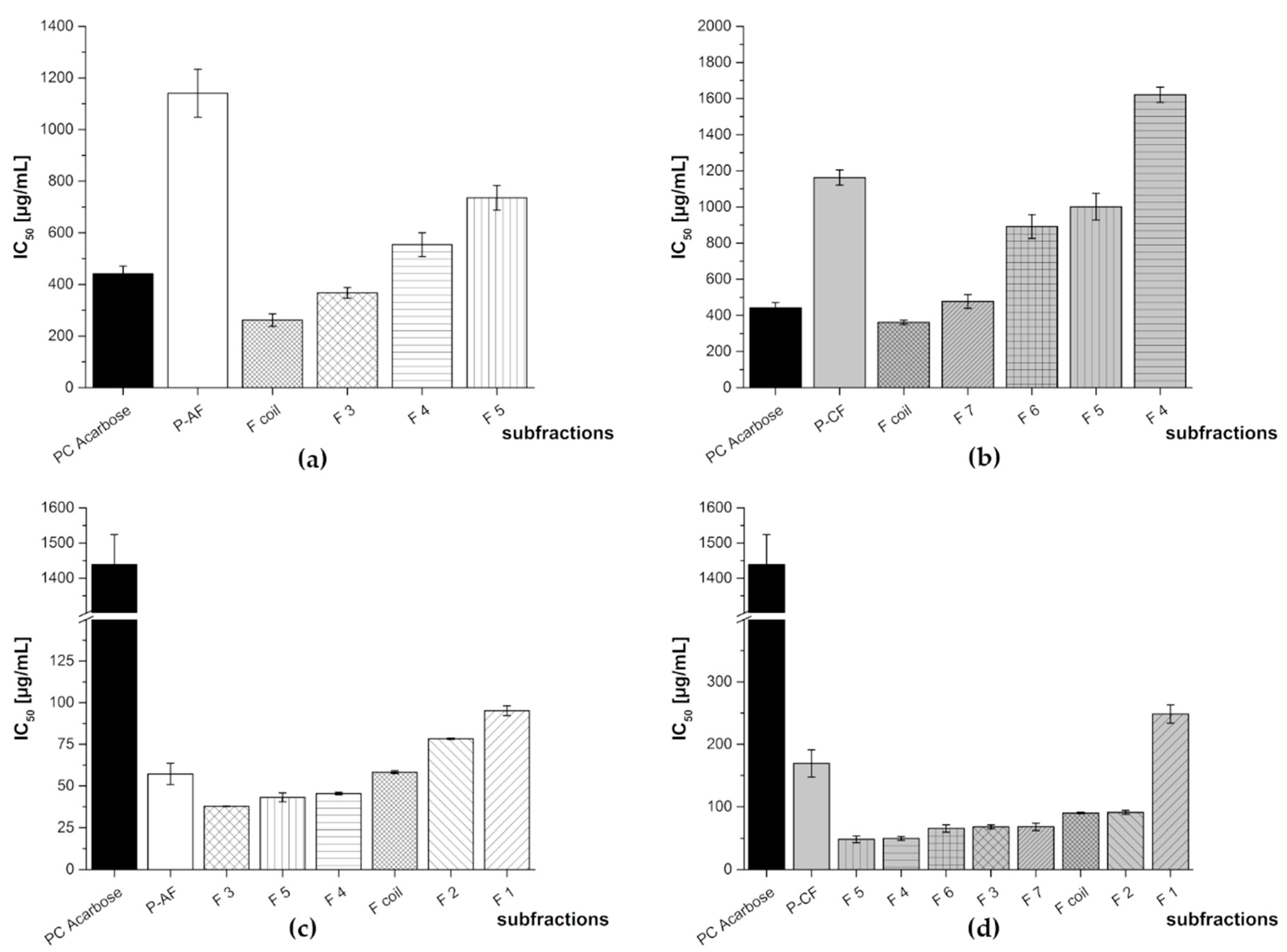
| P-AF | F1 | N/A | 95.2 ± 2.9 |
| F2 | N/A | 78.3 ± 0.4 | |
| F3 | 367 ± 21 | 37.8 ± 0.2 | |
| F4 | 554 ± 46 | 45.4 ± 0.8 | |
| F5 | 736 ± 48 | 43.1 ± 2.7 | |
| Fcoil | 262 ± 24 | 58.2 ± 0.8 | |
| P-CF | F1 | N/A | 249 ± 15 |
| F2 | N/A | 90.9 ± 3.2 | |
| F3 | N/A | 67.7 ± 3.1 | |
| F4 | 1621 ± 42 | 48.5 ± 5.4 | |
| F5 | 1001 ± 74 | 43.1 ± 2.7 | |
| F6 | 892 ± 66 | 65.4 ± 5.7 | |
| F7 | 477 ± 39 | 68.1 ± 5.7 | |
| Fcoil | 362 ± 12 | 90.1 ± 1.2 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ostberg-Potthoff, J.J.; Berger, K.; Richling, E.; Winterhalter, P. Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism. Nutrients 2019, 11, 1166. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051166
Ostberg-Potthoff JJ, Berger K, Richling E, Winterhalter P. Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism. Nutrients. 2019; 11(5):1166. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051166
Chicago/Turabian StyleOstberg-Potthoff, Johanna Josefine, Kirsten Berger, Elke Richling, and Peter Winterhalter. 2019. "Activity-Guided Fractionation of Red Fruit Extracts for the Identification of Compounds Influencing Glucose Metabolism" Nutrients 11, no. 5: 1166. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051166