Simulated Gastrointestinal Digestion of Cocoa: Detection of Resistant Peptides and In Silico/In Vitro Prediction of Their Ace Inhibitory Activity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cocoa Samples
2.2. Extraction of Peptides from Non-Digested Samples
2.3. Simulated In Vitro Gastro-Intestinal Digestion
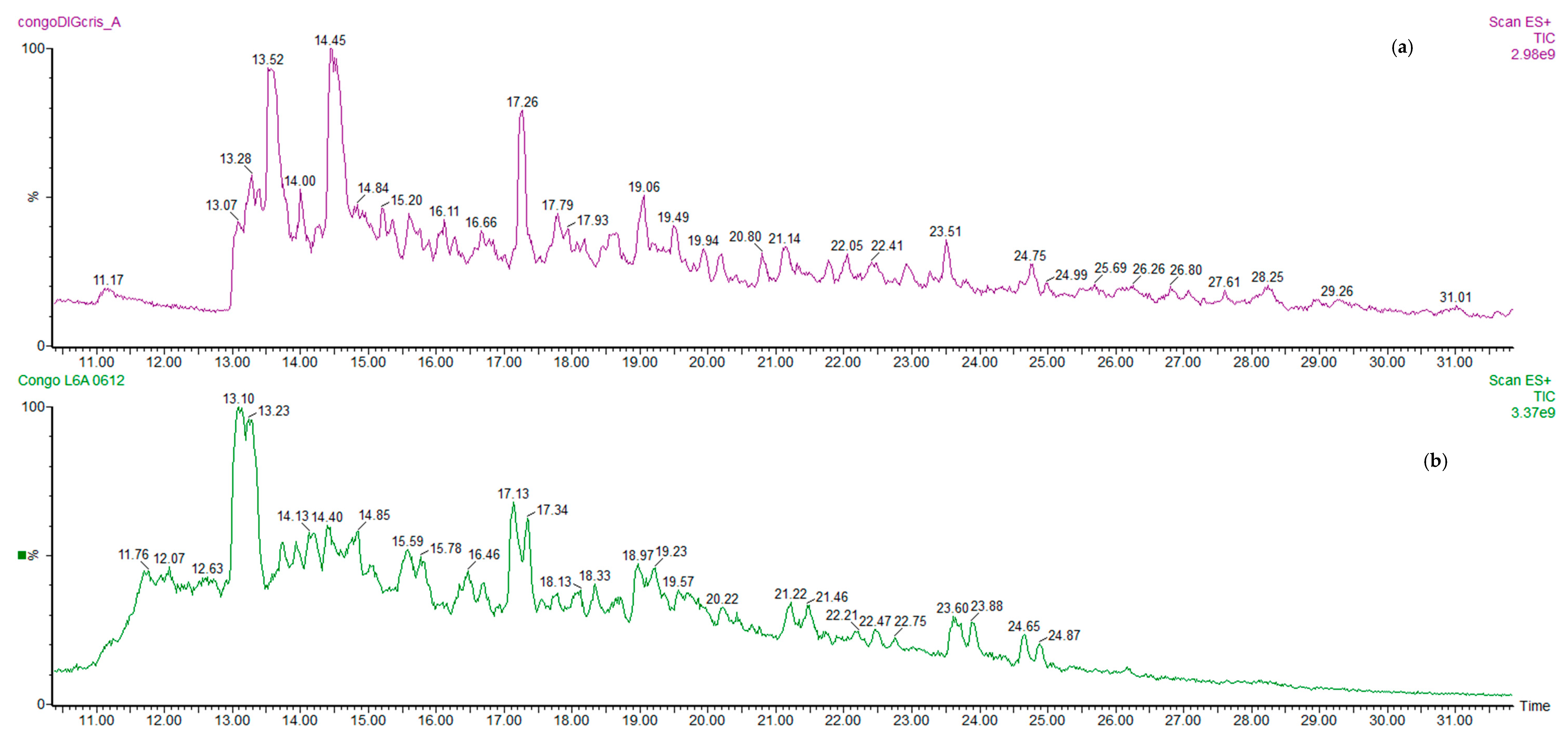
2.4. Ultra-High Performance Liquid Chromatography/Electrospray Ionisation Mass Spectrometry (UHPLC/ESI-MS) Analysis Conditions
2.5. Peptides Identification by High-Performance Liquid Chromatography/Tandem Mass Spectrometry (HPLC/MS-MS)
2.6. Peptides Quantification
2.7. Determination of In Vitro Angiotensin-Converting Enzyme (ACE) Inhibitory Activity of Cocoa Digestates
2.8. Prediction of ACE Inhibitory Activity of Cocoa Digestate Peptides by Computational Procedures
2.8.1. Pharmachopore Models
2.8.2. Molecular Modelling
2.8.3. Docking Simulations and Rescoring Procedure
2.8.4. Chemical Synthesis of Specific Peptides Predicted as Active
3. Results and Discussion
3.1. Potentially Bioactive Peptides
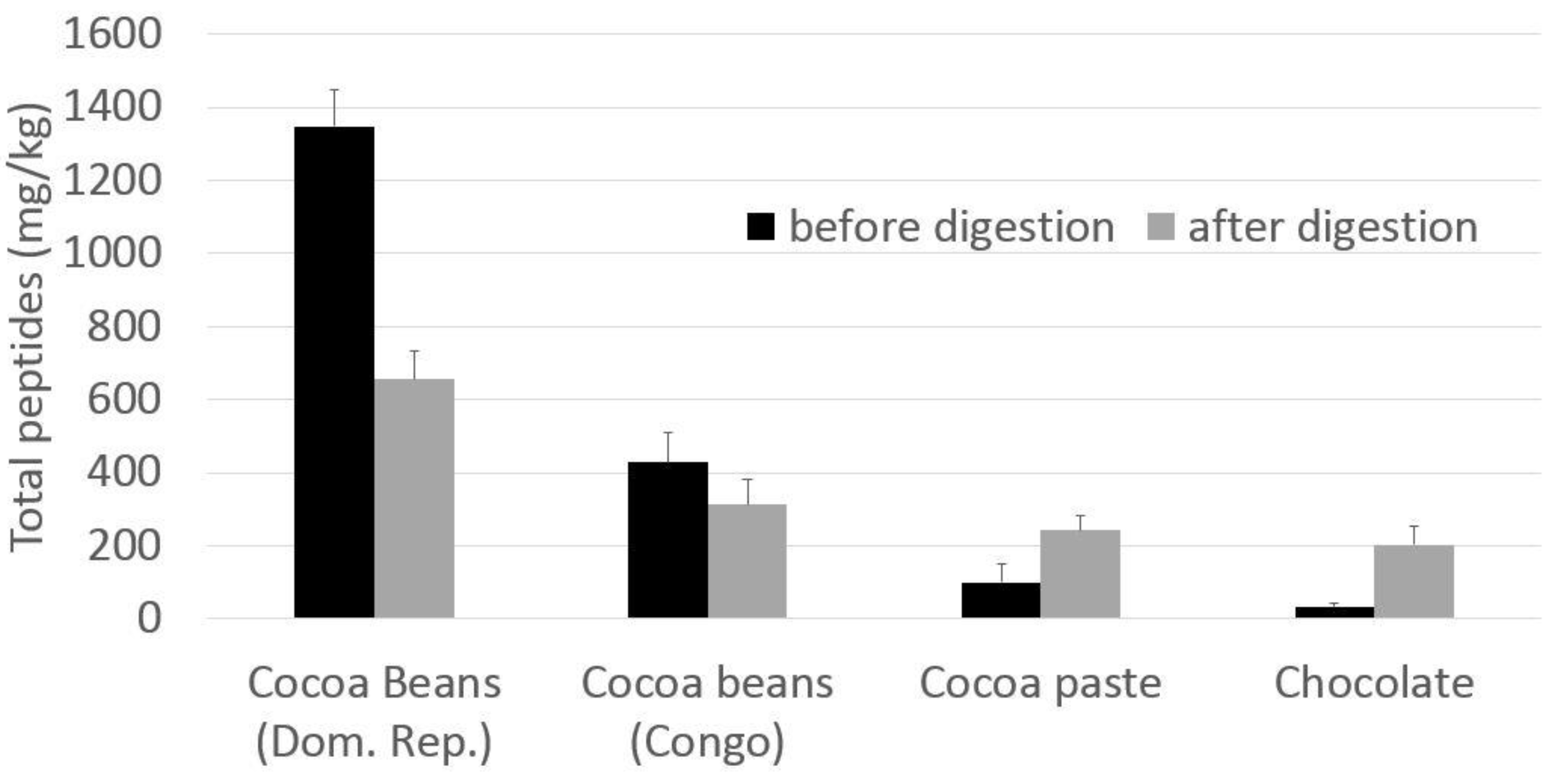
3.2. ACE Tests on Cocoa Digested Samples
3.3. In Silico Screening of Potential ACE Inhibitory Peptides
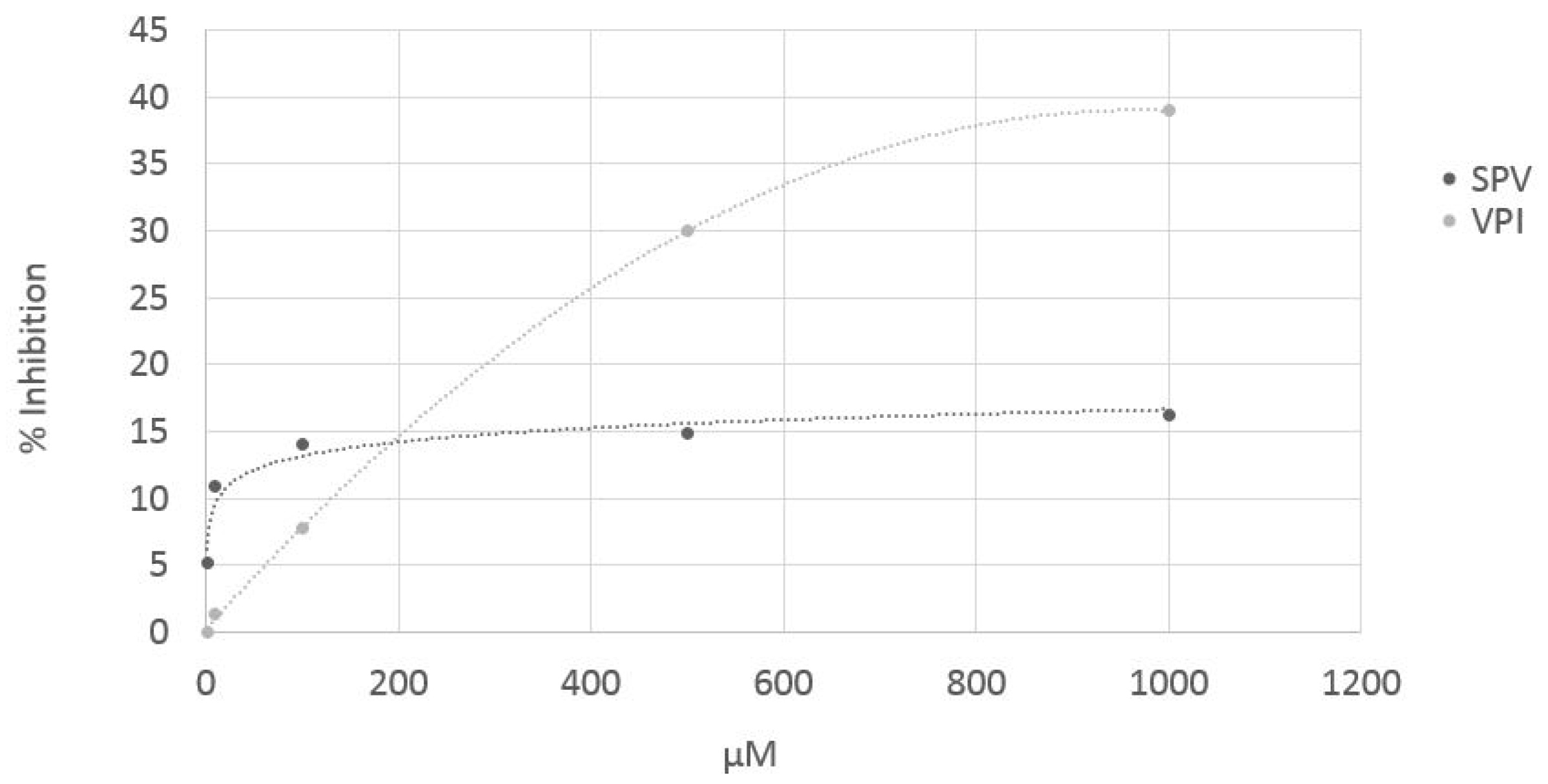
3.4. ACE in Vitro Test on Pure (Synthetized) Peptides
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Keen, C.L. Chocolate: Food as medicine/medicine as food. J. Am. Coll. Nutr. 2001, 20, 436S–439S. [Google Scholar] [CrossRef] [PubMed]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and chocolate in human health and disease. Antioxid. Redox. Signal 2011, 15, 2779–2811. [Google Scholar] [CrossRef] [PubMed]
- AAVV. Scientific Opinion on the substantiation of a health claim related to cocoa flavanols and maintenance of normal endothelium-dependent vasodilation pursuant to Article 13 (5) of Regulation (EC) No 1924/2006. EFSA J. 2012, 10, 2809. [Google Scholar]
- Rawel, H.M.; Huschek, G.; Sagu, S.T.; Homann, T. Cocoa bean proteins—Characterization, changes and modifications due to ripening and post-harvest processing. Nutrients 2019, 11, 428. [Google Scholar] [CrossRef]
- Hebert, E.M.; Saavedra, L.; Ferranti, P. Bioactive peptides derived from casein and whey proteins. In Biotechnology of Lactic Acid Bacteria: Novel Applications; Mozzi, F., Raya, R., Vignolo, G., Eds.; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 233–249. [Google Scholar]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, L.; Hebert, E.M.; Minahk, C.; Ferranti, P. An overview of “omic” analytical methods applied in bioactive peptide studies. Food Res. Int. 2013, 54, 925–934. [Google Scholar] [CrossRef]
- Marseglia, A.; Sforza, S.; Faccini, A.; Bencivenni, M.; Palla, G.; Caligiani, A. Extraction, identification and semi-quantification of oligopeptides in cocoa beans. Food Res. Int. 2014, 63, 382–389. [Google Scholar] [CrossRef]
- Caligiani, A.; Marseglia, A.; Prandi, B.; Palla, G.; Sforza, S. Influence of fermentation level and geographical origin on cocoa bean oligopeptide pattern. Food Chem. 2016, 211, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Grimbs, A.; D’Souza, R.N.; Verma, S.K.; Corno, M.; Kuhnert, N.; Ullrich, M.S. Origin and varietal based proteomic and peptidomic fingerprinting of Theobroma cacao in non-fermented and fermented cocoa beans. Food Res. Int. 2018, 111, 137–147. [Google Scholar] [CrossRef]
- D’Souza, R.N.; Grimbs, A.; Grimbs, S.; Behrends, B.; Corno, M.; Ullrich, M.S.; Kuhnert, N. Degradation of cocoa proteins into oligopeptides during spontaneous fermentation of cocoa beans. Food Res. Int. 2018, 109, 506–516. [Google Scholar] [CrossRef]
- Zak, D.L.; Keeney, P.G. Changes in cocoa proteins during ripening of fruit, fermentation, and further processing of cocoa beans. J. Agric. Food Chem. 1976, 24, 483–486. [Google Scholar] [CrossRef]
- Voigt, J.; Janek, K.; Textoris-Taube, K.; Niewienda, A.; Woestemeyer, J. Partial purification and characterisation of the peptide precursors of the cocoa-specific aroma components. Food Chem. 2016, 192, 706–713. [Google Scholar] [CrossRef]
- John, W.A.; Kumari, N.; Boettcher, N.L.; Koffi, K.J.; Grimbs, S.; Vrancken, G.; D’Souza, R.N.; Kuhnert, N.; Ullrich, M.S. Aseptic artificial fermentation of cocoa beans can be fashioned to replicate the peptide profile of commercial cocoa bean fermentations. Food Res. Int. 2016, 89, 764–772. [Google Scholar] [CrossRef]
- Spencer, M.E.; Hodge, R. Cloning and sequencing of a cDNA encoding the major storage proteins of Theobroma cacao. Planta 1992, 186, 567–576. [Google Scholar] [CrossRef]
- Kochhar, S.; Gartenmann, K.; Juillerat, M.A. Primary structure of the abundant seed albumin of theobroma cacao by mass spectrometry. J. Agric. Food Chem. 2000, 48, 5593–5599. [Google Scholar] [CrossRef]
- Amin, I.; Jinap, S.; Jamilah, B.; Harikrisna, K.; Biehl, B. Oligopeptide patterns produced from Theobroma cacao L of various genetic origins. J. Sci. Food Agric. 2002, 82, 733–737. [Google Scholar] [CrossRef]
- Sarmadi, B.; Amin, I.; Muhajir, H. Antioxidant and angiotensin converting enzyme (ACE) inhibitory activities of cocoa (Theobroma cacao L.) autolysates. Food Res. Int. 2011, 44, 290–296. [Google Scholar] [CrossRef]
- Sarmadi, B.; Farhana, A.; Muhajir, H.; Nazamid, S.; Azizah, A.H.; Amin, I. Hypoglycemic effects of cocoa (Theobroma cacao L.) autolysates. Food Chem. 2012, 134, 905–911. [Google Scholar] [CrossRef]
- Tovar-Perez, E.G.; Guerrero-Becerra, L.; Lugo-Cervantes, E. Antioxidant activity of hydrolysates and peptide fractions of glutelin from cocoa (Theobroma cacao L.) seed. CyTA J. Food 2017, 15, 489–496. [Google Scholar] [CrossRef]
- Muguerza Marquinez, B.; Monzo Oltra, H.; Alepuz Rico, N.; Bataller Leiva, E.; Genoves Martinez, S.; Enrique Lopez, M.; Martorell Guerola, P.; Ramon Vidal, D. Bioactive Peptides with ACE- and PEP-Inhibiting Activity Obtained from Cocoa Extracts. PCT Int. Appl. WO 2010012845 A1. 4 February 2010. [Google Scholar]
- Minekus, M.; Alminger, M.; Alvito, P.; Balance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Prandi, B.; Varani, M.; Faccini, A.; Lambertini, F.; Suman, M.; Leporati, A.; Tedeschi, T.; Sforza, S. Species specific marker peptides for meat authenticity assessment: A multispecies quantitative approach applied to Bolognese sauce. Food Contr. 2019, 97, 15–24. [Google Scholar] [CrossRef]
- Sforza, S.; Galaverna, G.; Schivazappa, C.; Marchelli, R.; Dossena, A.; Virgili, R. Effect of extended aging of parma dry-cured ham on the content of oligopeptides and free amino acids. J. Agric. Food Chem. 2006, 54, 9422–9429. [Google Scholar] [CrossRef]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Dellafiora, L.; Paolella, S.; Dall’Asta, C.; Dossena, A.; Cozzini, P.; Galaverna, G. Hybrid in silico/in vitro approach for the identification of angiotensin i converting enzyme inhibitory peptides from parma dry-cured ham. J. Agric. Food Chem. 2015, 63, 6366–6375. [Google Scholar] [CrossRef]
- Baroni, M.; Cruciani, G.; Sciabola, S.; Perruccio, F.; Mason, J.S. A Common reference framework for analyzing/comparing proteins and ligands. fingerprints for ligands and proteins (FLAP): Theory and application. J. Chem. Inform. Mod. 2007, 47, 279–294. [Google Scholar] [CrossRef]
- Amadasi, A.; Mozzarelli, A.; Meda, C.; Maggi, A.; Cozzini, P. Identification of xenoestrogens in food additives by an integrated in silico and in vitro approach. Chem. Res. Toxicol. 2009, 22, 52–63. [Google Scholar] [CrossRef]
- Cozzini, P.; Dellafiora, L. In silico approach to evaluate molecular interaction between mycotoxins and the estrogen receptors ligand binding domain: a case study on zearalenone and its metabolites. Toxicol. Lett. 2012, 214, 81–85. [Google Scholar] [CrossRef]
- Dellafiora, L.; Mena, P.; Cozzini, P.; Brighenti, F.; Del Rio, D. Modelling the possible bioactivity of ellagitannin-derived metabolites. In silico tools to evaluate their potential xenoestrogenic behavior. Food Funct. 2013, 4, 1442–1451. [Google Scholar] [CrossRef]
- Dellafiora, L.; Mena, P.; Del Rio, D.; Cozzini, P. Modeling the effect of phase II conjugations on topoisomerase I poisoning: Pilot study with luteolin and quercetin. J. Agric. Food Chem. 2014, 62, 5881–5886. [Google Scholar] [CrossRef]
- Salsi, E.; Bayden, A.S.; Spyrakis, F.; Amadasi, A.; Campanini, B.; Bettati, S.; Dodatko, T.; Cozzini, P.; Kellogg, G.E.; Cook, P.F.; et al. Design of O-acetylserine sulfhydrylase inhibitors by mimicking nature. J. Med. Chem. 2010, 53, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, G.E.; Abraham, D.J. Hydrophobicity: Is LogP(o/w) more than the sum of its parts? Eur. J. Med. Chem. 2000, 37, 651–661. [Google Scholar] [CrossRef]
- Cozzini, P.; Fornabaio, M.; Marabotti, A.; Abraham, D.J.; Kellogg, G.E.; Mozzarelli, A. Simple, intuitive calculations of free energy of binding for protein-ligand complexes. 1. Models without explicit constrained water. J. Med. Chem. 2002, 45, 2469–2483. [Google Scholar] [CrossRef] [PubMed]
- Escudero, E.; Sentandreu, M.A.; Arihara, K.; Toldra, F. Angiotensin I-converting enzyme inhibitory peptides generated from in vitro gastrointestinal digestion of pork meat. J. Agric. Food Chem. 2010, 58, 2895–2901. [Google Scholar] [CrossRef]
- Paolella, S.; Falavigna, C.; Faccini, A.; Virgili, R.; Sforza, S.; Dall’Asta, C.; Dossena, A.; Galaverna, G. Effect of dry-cured ham maturation time on simulated gastrointestinal digestion: Characterization of the released peptide fraction. Food Res. Int. 2015, 67, 136–144. [Google Scholar] [CrossRef]
- Agudelo, R.A.; Gauthier, S.F.; Pouliot, Y.; Marin, J.; Savoie, L. Kinetics of peptide fraction release during in vitro digestion of casein. J. Sci. Food Agric. 2004, 84, 325–332. [Google Scholar] [CrossRef]
- Roblet, C.; Amiot, J.; Lavigne, C.; Marette, A.; Lessard, M.M.; Jean, J.; Ramassamy, C.; Moresoli, C.; Bazinet, L. Screening of in vitro bioactivities of a soy protein hydrolysate separated by hollow fiber and spiral-wound ultrafiltration membranes. Food Res. Int. 2012, 46, 237–249. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides-Opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef]
- Miner-Williams, W.M.; Stevens, B.R.; Moughan, P.J. Are intact peptides absorbed from the healthy gut in the adult human? Nutr. Res. Rev. 2014, 27, 308–329. [Google Scholar] [CrossRef]
- Gonzalez de Mejia, E.; Martınez-Villaluenga, C.; Fernandez, D.; Urado, D.; Sato, K. Bioavailability and safety of food peptides. In Food Proteins and Peptides: Chemistry, Functionality Interactions, and Commercialization; Hettiarachchy, N.S., Sato, K., Kannan, A., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Chapter 12; pp. 297–330. [Google Scholar]
- Kannel, W.B.; Higgins, M. Smoking and hypertension as predictors of cardiovascular risk in population studies. J. Hypertens. Suppl. 1990, 8, S3–S8. [Google Scholar]
- Riordan, J.F. Angiotensin-I-converting enzyme and its relatives. Genome Biol. 2003, 4, 225. [Google Scholar] [CrossRef]
- Livelli di Assunzione Raccomandata di Nutrienti per la Popolazione Italiana. 2014. Available online: http://www.sinu.it/public/20141111_LARN_Porzioni.pdf (accessed on 29 April 2019).




{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | MH+ | Sequence | Cocoa Protein (Vicilin, V; Albumin, 21k) | Bioactivity | Cocoa Bean (Congo) before Digestion | Cocoa Bean (Congo) after Digestion | Cocoa Bean (Dominican Rep.) before Digestion | Cocoa Bean (Dominican Rep.) after Digestion | Cocoa Paste before Digestion | Cocoa Paste after Digestio | Chocolate before Digestion | Chocolate after Digestion |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10.62 | 203.2 | AI | v | Angiotensin-converting enzyme (ACE) inhibitor | 35.8 | 6.0 | 6.8 | 2.2 | 5.3 | 5.9 | 1.7 | 4.1 |
| 12.99 | 403.4 | RLD | 21k | 2.2 | 0.0 | 11.7 | 1.4 | 0.5 | 0.3 | 0.1 | 0.3 | |
| 13.02 | 295 | FE | 7.8 | 5.0 | 41.4 | 21.6 | 2.2 | 4.0 | 0.8 | 3.1 | ||
| 13.13 | 485.3 | 4.0 | 0.2 | 43.7 | 0.3 | 2.8 | 0.2 | 0.6 | 0.0 | |||
| 13.23 | 302 | SPV | 4.9 | 3.8 | 29.6 | 21.1 | 0.4 | 2.6 | 0.2 | 1.5 | ||
| 13.26 | 290.2 | GTI | v | 0.2 | 1.6 | 1.5 | 0.2 | 0.2 | 1.9 | 0.0 | 2.1 | |
| 13.35 | 237.2 | MS | 1.4 | 1.3 | 19.4 | 8.8 | 1.0 | 1.6 | 0.4 | 1.0 | ||
| 13.42 | 281.2 | DF | ACE inhibitor | 3.1 | 4.4 | 28.4 | 14.1 | 1.6 | 4.2 | 0.6 | 3.4 | |
| 13.5 | 223.2 | FG | V | ACE inhibitor (FG) | 14.2 | 5.6 | 51.4 | 27.3 | 3.5 | 4.2 | 1.2 | 2.8 |
| 13.74 | 229.3 | PI (L) | 21k/v | dipeptidyl peptidase IV inhibitor (PI, PL); ACE inhibitor (PL) | 4.0 | 7.1 | 20.3 | 12.7 | 2.3 | 12.0 | 0.8 | 13.3 |
| 13.81 | 231.2 | L (I) V | v/21k | glucose uptake stimulating peptide (IV, LV); dipeptidyl peptidase IV inhibitor (LV) | 13.7 | 17.4 | 2.0 | 0.0 | 2.0 | 27.0 | 0.7 | 30.2 |
| 14 | 276.2 | QE | dipeptidyl peptidase IV inhibitor | 9.1 | 6.1 | 28.2 | 8.9 | 2.6 | 1.7 | 1.1 | 1.3 | |
| 14.35 | 761.4 | RRSDLD | 21k | 0.0 | 0.3 | 0.3 | 0.4 | 0.0 | 0.3 | 0.0 | 0.3 | |
| 14.35 | 231.2 | VI (L) | v/21k | dipeptidyl peptidase IV inhibitor (VI, VL), glucose uptake stimulating peptide (VL) | 8.2 | 7.1 | 32.7 | 26.1 | 1.9 | 6.1 | 0.7 | 5.3 |
| 14.45 | 223.2 | GF | ACE inhibitor, dipeptidyl peptidase IV inhibitor | 3.5 | 4.0 | 0.0 | 4.2 | 0.3 | 5.8 | 0.1 | 6.7 | |
| 14.64 | 237.2 | AF | dipeptidyl peptidase IV inhibitor; ACE inhibitor | 17.0 | 4.3 | 33.7 | 14.6 | 2.9 | 4.4 | 0.9 | 2.8 | |
| 14.83 | 231.2 | L (I) V | v/21k | glucose uptake stimulating peptide (IV, LV); dipeptidyl peptidase IV inhibitor (LV) | 8.1 | 12.0 | 17.7 | 11.8 | 1.9 | 8.7 | 0.7 | 7.4 |
| 14.9 | 634.3 | VSTDVN | 21k | 0.3 | 0.3 | 11.8 | 5.1 | 0.3 | 0.9 | 0.0 | 0.5 | |
| 14.95 | 229 | PI (L) | dipeptidyl peptidase IV inhibitor (PI, PL); ACE inhibitor (PL) | 11.8 | 7.4 | 23.4 | 18.9 | 3.5 | 8.2 | 1.3 | 8.8 | |
| 15.1 | 431 | unk | 7.5 | 9.2 | 52.5 | 28.4 | 2.5 | 2.7 | 0.7 | 1.2 | ||
| 15.24 | 487.3 | ANSPV | 21k | 2.6 | 2.7 | 27.5 | 25.9 | 1.5 | 4.9 | 0.4 | 2.9 | |
| 15.32 | 295 | EF | CaMPDE inhibitor; Renin inhibitor (HYPOTENSIVE) | 6.3 | 4.4 | 23.1 | 11.9 | 1.3 | 2.8 | 0.3 | 2.7 | |
| 15.44 | 838.4 | DEEGNFK | v | 0.1 | 0.0 | 2.4 | 0.0 | 0.1 | 0.0 | 0.0 | 0.0 | |
| 15.77 | 231.2 | VI (L) | v/21 | dipeptidyl peptidase IV inhibitor (VI, VL), glucose uptake stimulating peptide (VL) | 44.6 | 12.5 | 17.7 | 11.8 | 6.7 | 11.6 | 2.3 | 6.8 |
| 15.8 | 488.3 | GAGGGGL | v | 4.8 | 3.6 | 28.4 | 12.5 | 0.6 | 0.8 | 0.1 | 0.5 | |
| 16.06 | 296.2 | YN | dipeptidyl peptidase IV inhibitor, ACE inhibitor | 6.9 | 7.3 | 14.8 | 13.1 | 1.7 | 3.7 | 0.7 | 1.8 | |
| 16.25 | 263.3 | FP | dipeptidyl peptidase IV inhibitor; ACE inhibitor | 3.8 | 5.3 | 10.7 | 7.5 | 0.8 | 3.1 | 0.3 | 3.5 | |
| 16.37 | 379.9 (757.4) | ASKDQPL | v | 1.3 | 0.2 | 4.4 | 0.7 | 1.4 | 0.6 | 0.3 | 0.3 | |
| 16.8 | 265.3 | FV | v/21k | 7.6 | 6.6 | 30.2 | 18.7 | 2.3 | 4.7 | 1.0 | 3.0 | |
| 17.16 | 245.2 | II IL LL LI | ACE inhibitor (IL), glucose uptake stimulating peptide; dipeptidyl peptidase IV inhibitor | 5.3 | 7.1 | 9.1 | 6.8 | 0.8 | 7.2 | 0.2 | 7.8 | |
| 17.3 | 276.1 | AW | 21k | ACE inhibitor; antioxidant; dipeptidyl peptidase IV inhibitor | 8.6 | 5.7 | 29.1 | 13.9 | 1.1 | 1.9 | 0.1 | 0.8 |
| 17.86 | 360.3 | VLE | V | 0.1 | 4.4 | 0.6 | 0.0 | 0.5 | 0.8 | 0.1 | 0.7 | |
| 18.1 | 265.2 | VF | V | ACE inhibitor; dipeptidyl peptidase IV inhibitor | 19.6 | 10.7 | 56.6 | 27.8 | 3.8 | 6.8 | 1.2 | 4.6 |
| 18.19 | 393.3 | FLN/SSIS | V/21k | 2.3 | 0.6 | 17.6 | 8.1 | 1.2 | 1.4 | 0.3 | 0.5 | |
| 18.19 | 245.2 | II IL LL LI | ACE inhibitor (IL), glucose uptake stimulating peptide; dipeptidyl peptidase IV inhibitor | 4.1 | 19.0 | 0.0 | 10.8 | 0.8 | 7.5 | 0.3 | 7.5 | |
| 18.22 | 710.4 | DEEGNF | V | 1.1 | 0.1 | 20.8 | 0.6 | 0.8 | 0.2 | 0.1 | 0.3 | |
| 18.57 | 243.2 | Pyroglu-LEU | 9.3 | 13.9 | 29.8 | 35.2 | 2.7 | 6.8 | 1.1 | 5.6 | ||
| 18.69 | 245 | II IL LL LI | ACE inhibitor (IL), glucose uptake stimulating peptide; dipeptidyl peptidase IV inhibitor | 19.6 | 15.8 | 0.0 | 9.9 | 4.1 | 11.8 | 1.3 | 8.5 | |
| 18.93 | 690.3 | NGKGTIT | V | 0.2 | 0.1 | 5.5 | 2.0 | 0.1 | 0.1 | 0.0 | 0.1 | |
| 19.05 | 328 | VPI | 35.7 | 25.5 | 133.3 | 72.4 | 1.9 | 7.7 | 0.6 | 5.6 | ||
| 19.53 | 243.2 | Pyroglu-ILE | 6.5 | 7.9 | 24.2 | 17.2 | 2.5 | 4.4 | 1.0 | 3.9 | ||
| 19.7 | 245.2 | II IL LL LI | ACE inhibitor (IL), glucose uptake stimulating peptide; dipeptidyl peptidase IV inhibitor | 7.2 | 9.3 | 20.2 | 12.8 | 1.8 | 9.8 | 0.5 | 8.8 | |
| 19.85 | 534.3 | PGDVF | V | 0.0 | 0.0 | 17.5 | 0.0 | 0.6 | 0.1 | 0.2 | 0.0 | |
| 19.89 | 933.6 | DSKDDVVR | 21k | 0.1 | 0.0 | 2.8 | 0.0 | 0.1 | 0.1 | 0.0 | 0.1 | |
| 20.16 | 564.3 | RRSF | V | 0.6 | 0.0 | 5.5 | 1.1 | 0.2 | 0.1 | 0.1 | 0.2 | |
| 20.22 | 279.3 | L (I) F | ACE inhibitor | 7.1 | 7.7 | 27.3 | 13.3 | 1.6 | 4.3 | 0.8 | 3.2 | |
| 20.28 | 563.3 | DEEGN | V | 3.9 | 0.0 | 21.8 | 0.0 | 0.8 | 0.3 | 0.1 | 0.3 | |
| 20.32 | 360.3 | EVL | V | 2.6 | 0.0 | 4.4 | 3.4 | 0.3 | 0.3 | 0.1 | 0.2 | |
| 20.58 | 862.5 | SSISGAGGGGL | 21K | 0.3 | 0.1 | 12.5 | 3.8 | 0.3 | 0.7 | 0.1 | 0.4 | |
| 21 | 437.3 | GDVF | V | 0.6 | 0.2 | 5.5 | 0.0 | 0.6 | 0.1 | 0.2 | 0.2 | |
| 21.16 | 279.3 | L (I) F | ACE inhibitor | 24.5 | 18.3 | 74.7 | 60.0 | 4.9 | 19.1 | 1.8 | 11.8 | |
| 21.19 | 600.4 | KDQPL | V | 0.8 | 0.0 | 29.4 | 0.0 | 0.9 | 0.0 | 0.2 | 0.2 | |
| 21.31 | 277 | Pyroglu-PHE | 8.8 | 9.0 | 24.1 | 22.0 | 3.2 | 4.2 | 1.3 | 2.2 | ||
| 21.4 | 380.2 | DVF | V | 8.0 | 0.1 | 25.0 | 1.9 | 1.6 | 0.3 | 0.6 | 0.2 | |
| 22.09 | 279.3 | FL (I) | dipeptidyl peptidase IV inhibitor (FL) | 3.6 | 3.3 | 30.2 | 0.0 | 2.3 | 4.2 | 0.7 | 3.2 | |
| 22.41 | 279.3 | FL (I) | v | 9.4 | 2.2 | 30.2 | 0.0 | 2.3 | 3.2 | 0.7 | 1.9 | |
| 23 | 621.5 | SPGDVF | V | 0.4 | 0.0 | 37.7 | 0.5 | 1.6 | 0.1 | 0.5 | 0.0 | |
| 23.96 | 408.3 | IEF | 21K | 1.9 | 0.0 | 17.9 | 0.4 | 1.8 | 0.1 | 0.5 | 0.0 | |
| 25.69 | 603.3 (1204) | SNADSKDDVVR | 21k | 0.0 | 2.3 | 0.3 | 0.0 | 0.0 | 2.2 | 0.0 | 4.1 | |
| 26.31 | 789.6 | TVWRLD | 21K | 0.0 | 0.6 | 0.2 | 0.1 | 0.0 | 0.0 | 0.0 | 0.3 | |
| 27.59 | 601.3 | NNKPE | 0.0 | 3.6 | 0.7 | 0.0 | 0.0 | 1.3 | 0.0 | 1.7 | ||
| 27.92 | 747.5 | NGTPVIF | 21K | 0.6 | 0.0 | 12.7 | 0.4 | 0.1 | 0.0 | 0.0 | 0.1 | |
| 28.13 | 533.1 (1063.5) | DEEGNFKIL | V | 0.0 | 0.0 | 2.3 | 0.4 | 0.2 | 0.1 | 0.0 | 0.0 | |
| 28.54 | 902.6 | APLSPGDVF | V | 0.0 | 0.0 | 1.2 | 0.0 | 0.3 | 0.0 | 0.1 | 0.0 | |
| 29.76 | 820.5 | DNEWAW | 21K | 0.1 | 0.0 | 2.2 | 0.0 | 0.0 | 0.1 | 0.0 | 0.2 | |
| Total peptides | 427.9 | 313.2 | 1348.7 | 655.0 | 100.0 | 242.3 | 33.1 | 203.2 |
| Solution | Concentration (mg/mL) | Esteemed Peptide Concentration (μg/mL) | I% |
|---|---|---|---|
| Theobromine | 0.3 | 0 | 19 ± 5 |
| Epicatechin | 0.2 | 0 | 20 ± 5 |
| Amino acid mixture | 5 | 0 | 5 ± 4 |
| Cocoa bean digestate | 50 | 33 | 95 ± 4 |
| Cocoa paste digestate | 50 | 12 | 63 ± 10 |
| Chocolate digestate | 50 | 10 | 75 ± 8 |
| Cdominio (cutoff 260) | Ndominio (cutoff 540) | TOT | |||
|---|---|---|---|---|---|
| Compound | HS | Interaction | HS | Interaction | |
| FE | 2995 | Positive | 2443 | positive | Active |
| FV | 2494 | Positive | 2776 | Positive | Active |
| II | 2042 | Positive | 2380 | Positive | Active |
| IL | 1935 | Positive | 2262 | Positive | Active |
| LI | 2053 | Positive | 2045 | Positive | Active |
| LL | 1792 | Positive | 2062 | Positive | Active |
| LY | 3110 | Positive | 2709 | Positive | Active |
| PI | 1811 | Positive | 1717 | positive | Active |
| PL | 1992 | Positive | 1550 | Positive | Active |
| QE | 2091 | Positive | 1466 | Positive | Active |
| SPV | 1171 | Positive | 2075 | Positive | Active |
| VEL | 757 | Positive | 2160 | Positive | Active |
| VF | 2406 | Positive | 2366 | Positive | Active |
| VI | 1475 | positive | 2364 | positive | Active |
| VL | 1584 | positive | 2212 | positive | Active |
| VPI | 1258 | positive | 1707 | positive | Active |
| VPL | 1200 | positive | 1203 | positive | Active |
| LLDR | −913 | negative | 1224 | positive | Active |
| QLGN | 799 | positive | 413 | negative | Active |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marseglia, A.; Dellafiora, L.; Prandi, B.; Lolli, V.; Sforza, S.; Cozzini, P.; Tedeschi, T.; Galaverna, G.; Caligiani, A. Simulated Gastrointestinal Digestion of Cocoa: Detection of Resistant Peptides and In Silico/In Vitro Prediction of Their Ace Inhibitory Activity. Nutrients 2019, 11, 985. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11050985
Marseglia A, Dellafiora L, Prandi B, Lolli V, Sforza S, Cozzini P, Tedeschi T, Galaverna G, Caligiani A. Simulated Gastrointestinal Digestion of Cocoa: Detection of Resistant Peptides and In Silico/In Vitro Prediction of Their Ace Inhibitory Activity. Nutrients. 2019; 11(5):985. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11050985
Chicago/Turabian StyleMarseglia, Angela, Luca Dellafiora, Barbara Prandi, Veronica Lolli, Stefano Sforza, Pietro Cozzini, Tullia Tedeschi, Gianni Galaverna, and Augusta Caligiani. 2019. "Simulated Gastrointestinal Digestion of Cocoa: Detection of Resistant Peptides and In Silico/In Vitro Prediction of Their Ace Inhibitory Activity" Nutrients 11, no. 5: 985. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11050985