Diurnal Variation of Markers for Cholesterol Synthesis, Cholesterol Absorption, and Bile Acid Synthesis: A Systematic Review and the Bispebjerg Study of Diurnal Variations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Systematic Review
2.1.1. Search Methods
2.1.2. Selection Procedure
2.1.3. Data Extraction
2.2. Bispebjerg Study of Diurnal Variations
2.2.1. Subjects and Study Design
2.2.2. Serum Analyses
2.2.3. Statistical Analyses
3. Results
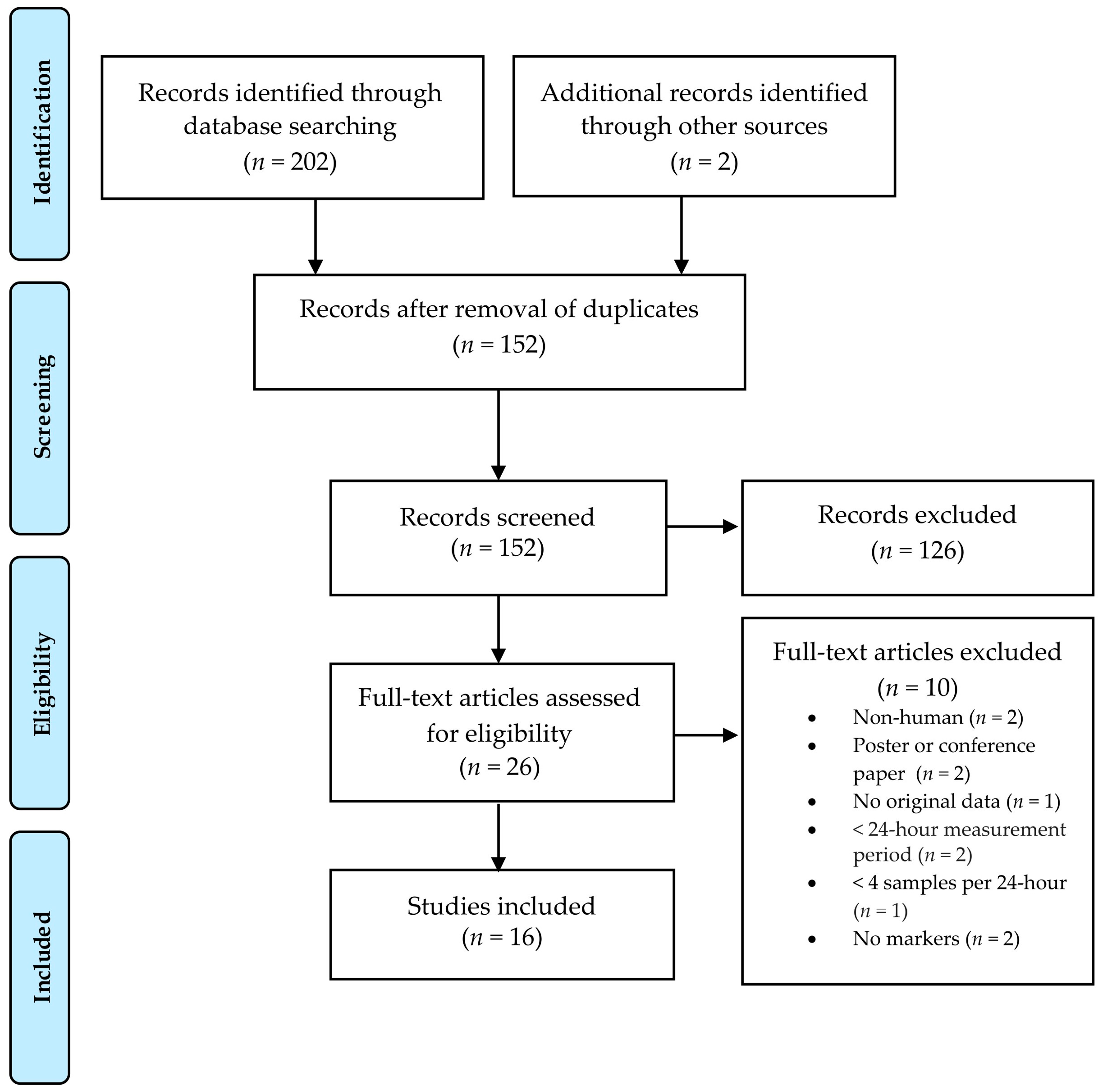
3.1. Systematic Review
3.1.1. Study Selection
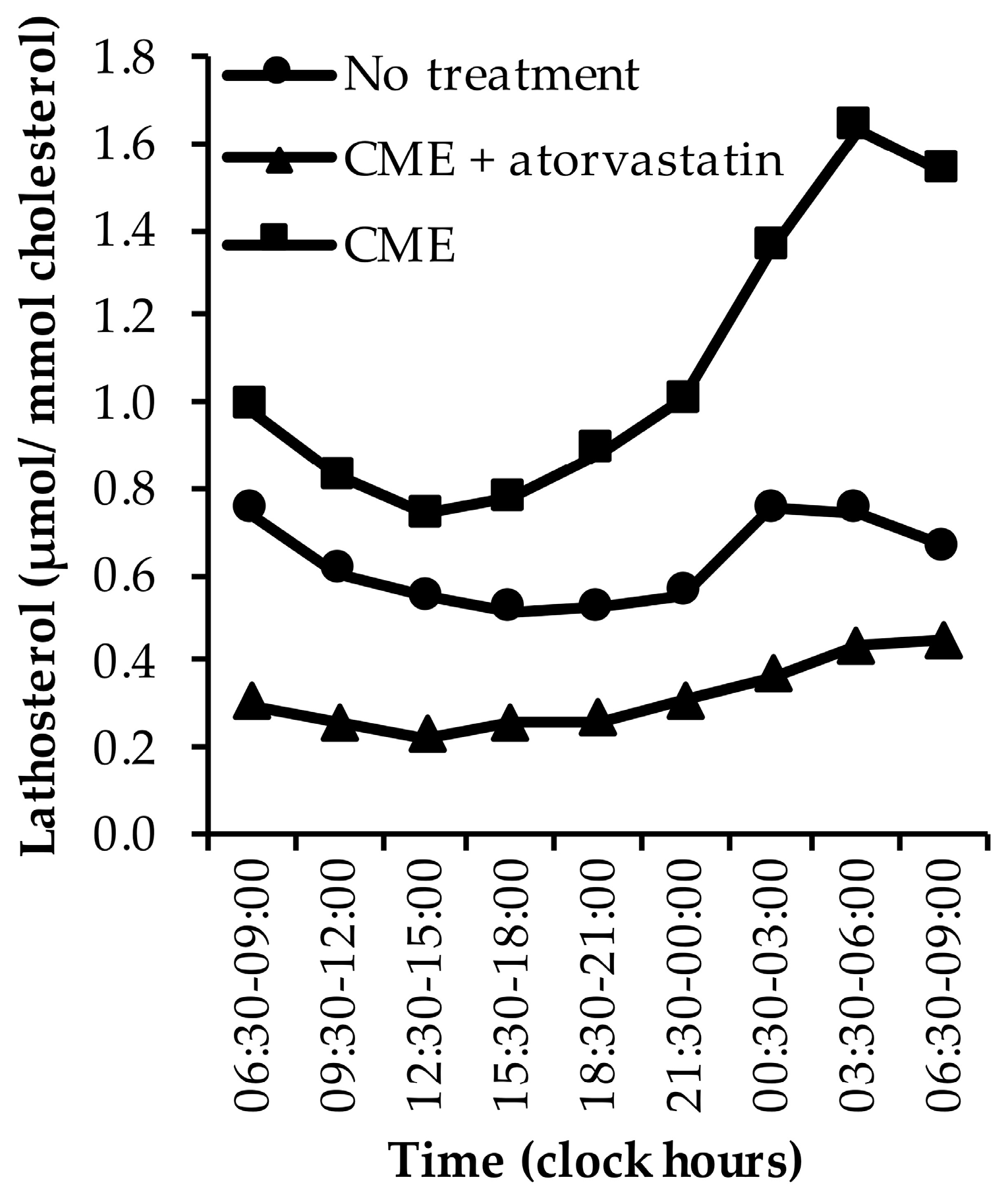
3.1.2. Lathosterol
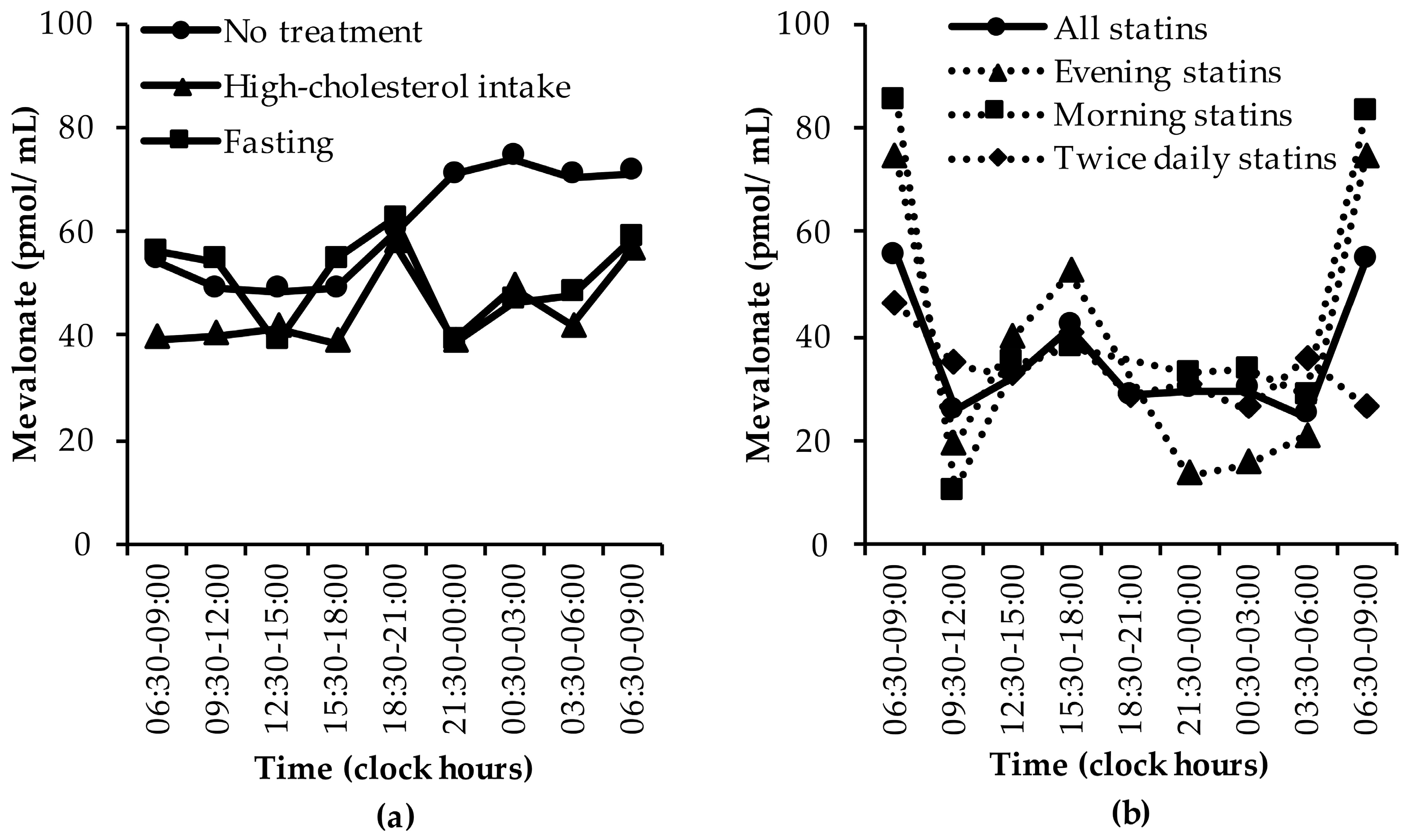
3.1.3. Mevalonate (MVA)
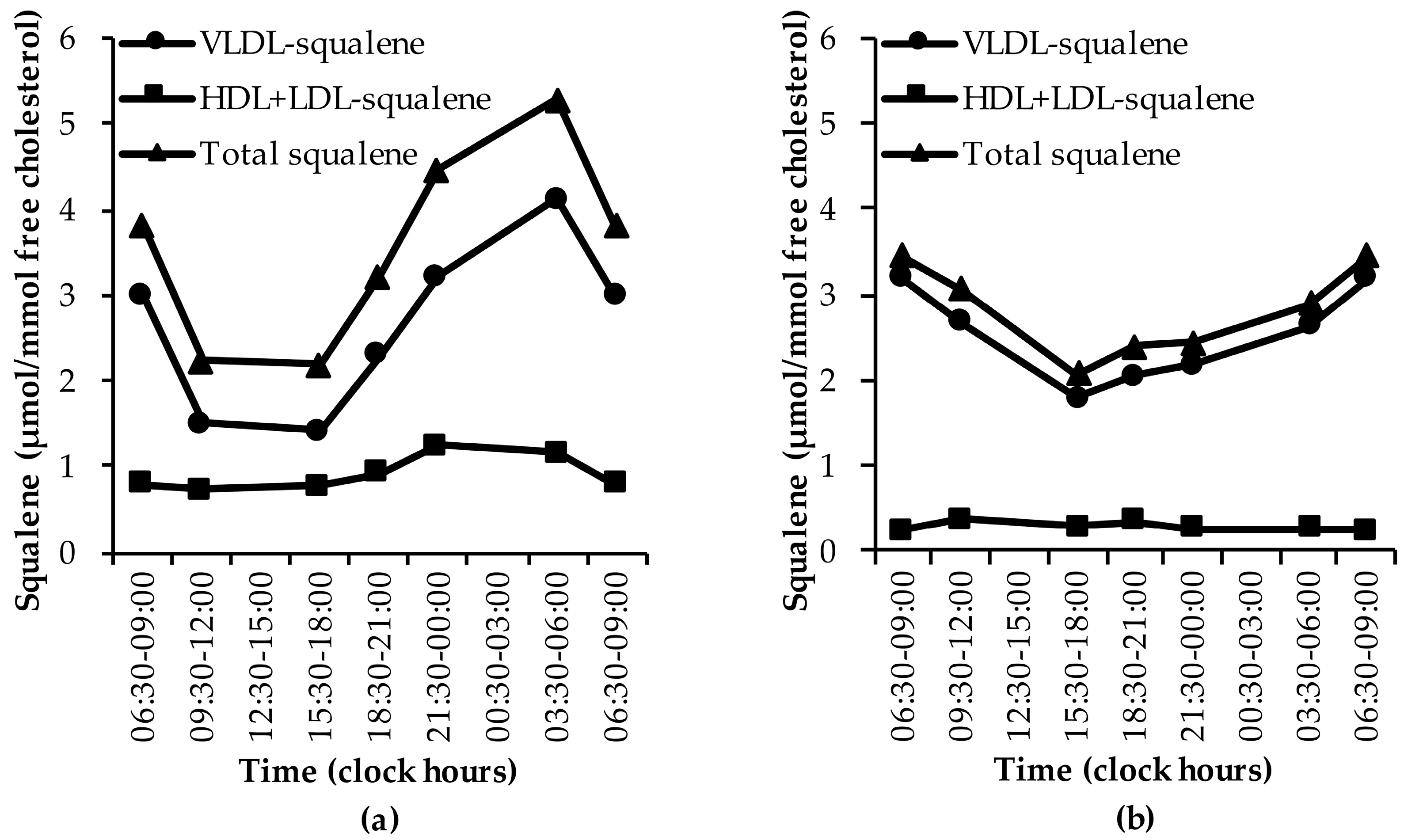
3.1.4. Squalene
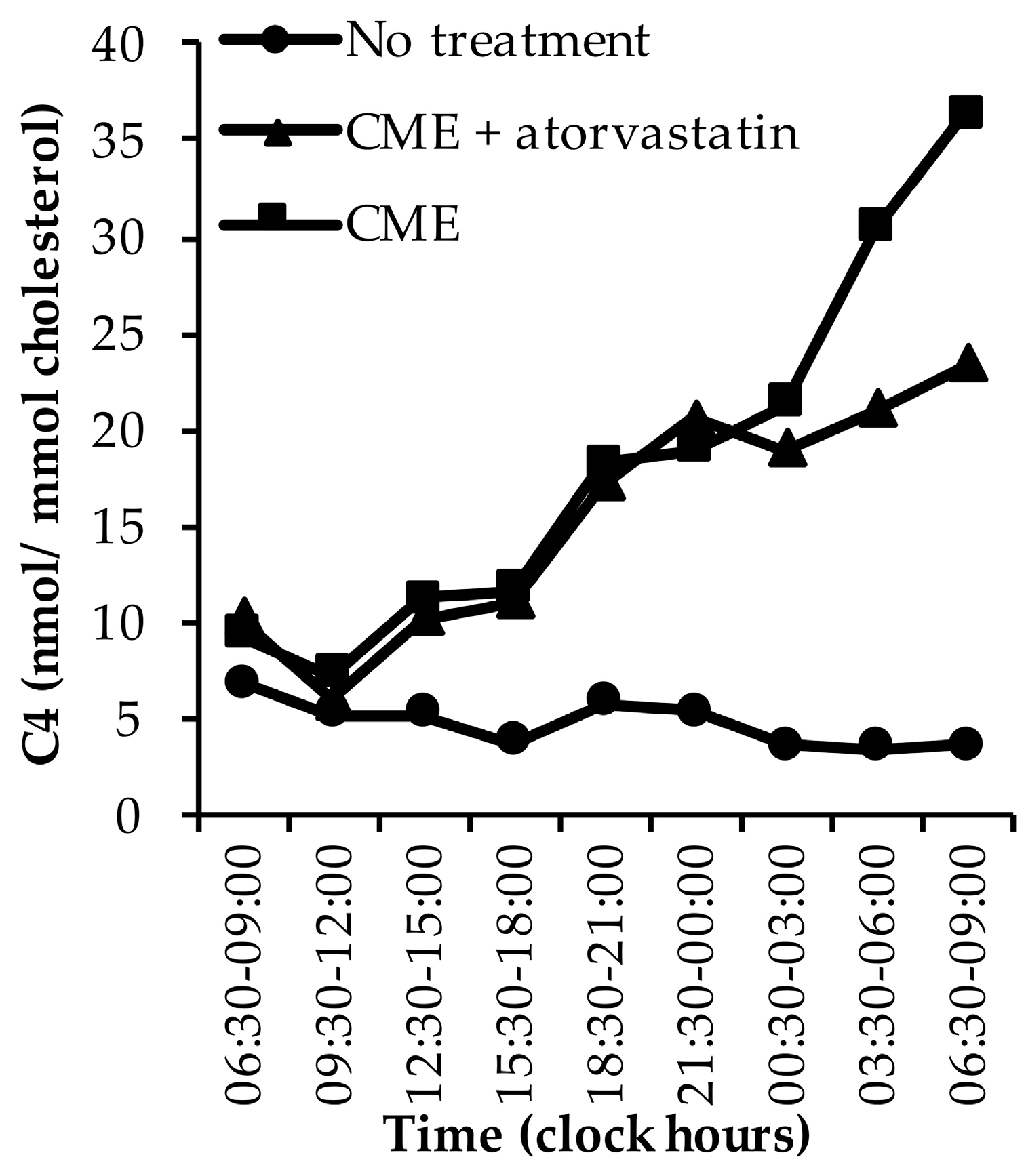
3.1.5. C4
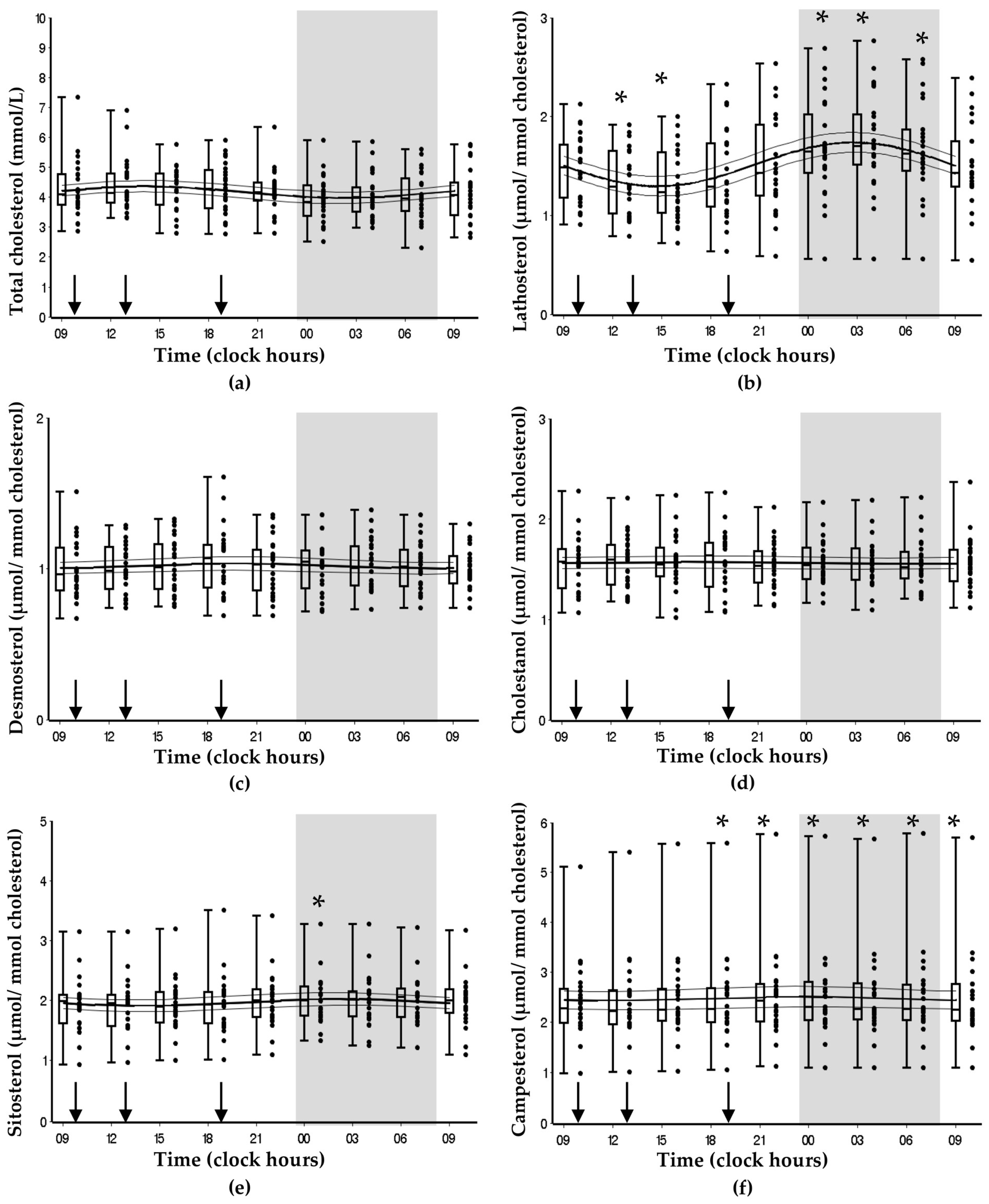
3.2. Bispebjerg Study of Diurnal Variations
4. Discussion
4.1. Endogenous Cholesterol Synthesis
4.2. Intestinal Cholesterol Absorption
4.3. Bile Acid Synthesis
4.4. Total Cholesterol
4.5. Discrepancies Between Studies Included in the Systematic Review
4.6. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reppert, S.M.; Weaver, D.R. Molecular analysis of mammalian circadian rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef] [PubMed]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metabolism 2018, 84, 11–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szosland, D. Shift work and metabolic syndrome, diabetes mellitus and ischaemic heart disease. Int. J. Occup. Med. Environ. Health 2010, 23, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, M.V.; Garg, A.X.; Iansavichus, A.V.; Costella, J.; Donner, A.; Laugsand, L.E.; Janszky, I.; Mrkobrada, M.; Parraga, G.; Hackam, D.G. Shift work and vascular events: Systematic review and meta-analysis. BMJ 2012, 345, e4800. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Purvis, T.E.; Hu, K.; Scheer, F.A. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc. Natl. Acad. Sci. USA 2016, 113, E1402–E1411. [Google Scholar] [CrossRef]
- Wefers, J.; van Moorsel, D.; Hansen, J.; Connell, N.J.; Havekes, B.; Hoeks, J.; van Marken Lichtenbelt, W.D.; Duez, H.; Phielix, E.; Kalsbeek, A.; et al. Circadian misalignment induces fatty acid metabolism gene profiles and compromises insulin sensitivity in human skeletal muscle. Proc. Natl. Acad. Sci. USA 2018, 115, 7789–7794. [Google Scholar] [CrossRef] [Green Version]
- Scheer, F.A.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [Green Version]
- Leproult, R.; Holmback, U.; Van Cauter, E. Circadian misalignment augments markers of insulin resistance and inflammation, independently of sleep loss. Diabetes 2014, 63, 1860–1869. [Google Scholar] [CrossRef]
- Kruit, J.K.; Groen, A.K.; van Berkel, T.J.; Kuipers, F. Emerging roles of the intestine in control of cholesterol metabolism. World J. Gastroenterol. 2006, 12, 6429–6439. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Tilvis, R.S.; Kesaniemi, Y.A. Serum cholestanol and plant sterol levels in relation to cholesterol metabolism in middle-aged men. Metabolism 1989, 38, 136–140. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Tilvis, R.S.; Kesaniemi, Y.A. Serum plant sterols and cholesterol precursors reflect cholesterol absorption and synthesis in volunteers of a randomly selected male population. Am. J. Epidemiol. 1990, 131, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Sauter, G.; Berr, F.; Beuers, U.; Fischer, S.; Paumgartner, G. Serum concentrations of 7alpha-hydroxy-4-cholesten-3-one reflect bile acid synthesis in humans. Hepatology 1996, 24, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Simonen, P.; Gylling, H.; Miettinen, T.A. The validity of serum squalene and non-cholesterol sterols as surrogate markers of cholesterol synthesis and absorption in type 2 diabetes. Atherosclerosis 2008, 197, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Matthan, N.R.; Raeini-Sarjaz, M.; Lichtenstein, A.H.; Ausman, L.M.; Jones, P.J. Deuterium uptake and plasma cholesterol precursor levels correspond as methods for measurement of endogenous cholesterol synthesis in hypercholesterolemic women. Lipids 2000, 35, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Quintao, E.; Grundy, S.M.; Ahrens, E.H., Jr. Effects of dietary cholesterol on the regulation of total body cholesterol in man. J. Lipid Res. 1971, 12, 233–247. [Google Scholar] [PubMed]
- Thuluva, S.C.; Igel, M.; Giesa, U.; Lutjohann, D.; Sudhop, T.; von Bergmann, K. Ratio of lathosterol to campesterol in serum predicts the cholesterol-lowering effect of sitostanol-supplemented margarine. Int. J. Clin. Pharmacol. Ther. 2005, 43, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Santosa, S.; Varady, K.A.; AbuMweis, S.; Jones, P.J. Physiological and therapeutic factors affecting cholesterol metabolism: Does a reciprocal relationship between cholesterol absorption and synthesis really exist? Life Sci. 2007, 80, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, T.A.; Strandberg, T.E.; Gylling, H. Noncholesterol sterols and cholesterol lowering by long-term simvastatin treatment in coronary patients: Relation to basal serum cholestanol. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1340–1346. [Google Scholar] [CrossRef]
- Jones, P.J.; Schoeller, D.A. Evidence for diurnal periodicity in human cholesterol synthesis. J. Lipid Res. 1990, 31, 667–673. [Google Scholar]
- Duane, W.C.; Levitt, D.G.; Mueller, S.M.; Behrens, J.C. Regulation of bile acid synthesis in man. Presence of a diurnal rhythm. J. Clin. Investig. 1983, 72, 1930–1936. [Google Scholar] [CrossRef]
- Sennels, H.P.; Jorgensen, H.L.; Fahrenkrug, J. Diurnal changes of biochemical metabolic markers in healthy young males-the Bispebjerg study of diurnal variations. Scand. J. Clin. Lab. Investig. 2015, 75, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Sennels, H.P.; Jorgensen, H.L.; Hansen, A.L.; Goetze, J.P.; Fahrenkrug, J. Diurnal variation of hematology parameters in healthy young males: The Bispebjerg study of diurnal variations. Scand. J. Clin. Lab. Investig. 2011, 71, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Oosterman, J.E.; Kalsbeek, A.; la Fleur, S.E.; Belsham, D.D. Impact of nutrients on circadian rhythmicity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R337–R350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, D.S.; Jones, P.J.; Myrie, S.B.; Plat, J.; Lutjohann, D. Methodological considerations for the harmonization of non-cholesterol sterol bio-analysis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 957, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, G. Cosinor-based rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef]
- Nelson, W.; Tong, Y.L.; Lee, J.K.; Halberg, F. Methods for cosinor-rhythmometry. Chronobiologia 1979, 6, 305–323. [Google Scholar]
- Kopito, R.R.; Weinstock, S.B.; Freed, L.E.; Murray, D.M.; Brunengraber, H. Metabolism of plasma mevalonate in rats and humans. J. Lipid Res. 1982, 23, 577–583. [Google Scholar]
- Miettinen, T.A. Diurnal variation of cholesterol precursors squalene and methyl sterols in human plasma lipoproteins. J. Lipid Res. 1982, 23, 466–473. [Google Scholar]
- Parker, T.S.; McNamara, D.J.; Brown, C.; Garrigan, O.; Kolb, R.; Batwin, H.; Ahrens, E.H., Jr. Mevalonic acid in human plasma: Relationship of concentration and circadian rhythm to cholesterol synthesis rates in man. Proc. Natl. Acad. Sci. USA 1982, 79, 3037–3041. [Google Scholar] [CrossRef]
- Parker, T.S.; McNamara, D.J.; Brown, C.D.; Kolb, R.; Ahrens, E.H., Jr.; Alberts, A.W.; Tobert, J.; Chen, J.; De Schepper, P.J. Plasma mevalonate as a measure of cholesterol synthesis in man. J. Clin. Investig. 1984, 74, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, T.A. Cholesterol precursors and their diurnal rhythm in lipoproteins of patients with jejuno-ileal bypass and ileal dysfunction. Metabolism 1985, 34, 425–430. [Google Scholar] [CrossRef]
- Scoppola, A.; Maher, V.M.; Thompson, G.R.; Rendell, N.B.; Taylor, G.W. Quantitation of plasma mevalonic acid using gas chromatography-electron capture mass spectrometry. J. Lipid Res. 1991, 32, 1057–1060. [Google Scholar] [PubMed]
- Jones, P.J.; Pappu, A.S.; Illingworth, D.R.; Leitch, C.A. Correspondence between plasma mevalonic acid levels and deuterium uptake in measuring human cholesterol synthesis. Eur. J. Clin. Investig. 1992, 22, 609–613. [Google Scholar] [CrossRef]
- Pappu, A.S.; Illingworth, D.R. Diurnal variations in the plasma concentrations of mevalonic acid in patients with abetalipoproteinaemia. Eur. J. Clin. Investig. 1994, 24, 698–702. [Google Scholar] [CrossRef]
- Yoshida, T.; Honda, A.; Tanaka, N.; Matsuzaki, Y.; Shoda, J.; He, B.; Osuga, T.; Miyazaki, H. Determination of 7 alpha-hydroxy-4-cholesten-3-one level in plasma using isotope-dilution mass spectrometry and monitoring its circadian rhythm in human as an index of bile acid biosynthesis. J. Chromatogr. B Biomed. Appl. 1994, 655, 179–187. [Google Scholar] [CrossRef]
- Nozaki, S.; Nakagawa, T.; Nakata, A.; Yamashita, S.; Kameda-Takemura, K.; Nakamura, T.; Keno, Y.; Tokunaga, K.; Matsuzawa, Y. Effects of pravastatin on plasma and urinary mevalonate concentrations in subjects with familial hypercholesterolaemia: A comparison of morning and evening administration. Eur. J. Clin. Pharmacol. 1996, 49, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Pappu, A.S.; Illingworth, D.R. The effects of lovastatin and simvastatin on the diurnal periodicity of plasma mevalonate concentrations in patients with heterozygous familial hypercholesterolemia. Atherosclerosis 2002, 165, 137–144. [Google Scholar] [CrossRef]
- Martin, P.D.; Mitchell, P.D.; Schneck, D.W. Pharmacodynamic effects and pharmacokinetics of a new HMG-CoA reductase inhibitor, rosuvastatin, after morning or evening administration in healthy volunteers. Br. J. Clin. Pharmacol. 2002, 54, 472–477. [Google Scholar] [CrossRef]
- Galman, C.; Angelin, B.; Rudling, M. Bile acid synthesis in humans has a rapid diurnal variation that is asynchronous with cholesterol synthesis. Gastroenterology 2005, 129, 1445–1453. [Google Scholar] [CrossRef]
- Persson, L.; Cao, G.; Stahle, L.; Sjoberg, B.G.; Troutt, J.S.; Konrad, R.J.; Galman, C.; Wallen, H.; Eriksson, M.; Hafstrom, I.; et al. Circulating proprotein convertase subtilisin kexin type 9 has a diurnal rhythm synchronous with cholesterol synthesis and is reduced by fasting in humans. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Steiner, C.; Othman, A.; Saely, C.H.; Rein, P.; Drexel, H.; von Eckardstein, A.; Rentsch, K.M. Bile acid metabolites in serum: Intraindividual variation and associations with coronary heart disease, metabolic syndrome and diabetes mellitus. PLoS ONE 2011, 6, e25006. [Google Scholar] [CrossRef] [PubMed]
- Al-Khaifi, A.; Straniero, S.; Voronova, V.; Chernikova, D.; Sokolov, V.; Kumar, C.; Angelin, B.; Rudling, M. Asynchronous rhythms of circulating conjugated and unconjugated bile acids in the modulation of human metabolism. J. Intern. Med. 2018, 284, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Cella, L.K.; Van Cauter, E.; Schoeller, D.A. Effect of meal timing on diurnal rhythm of human cholesterol synthesis. Am. J. Physiol. 1995, 269, E878–E883. [Google Scholar] [CrossRef] [PubMed]
- Cella, L.K.; Van Cauter, E.; Schoeller, D.A. Diurnal rhythmicity of human cholesterol synthesis: Normal pattern and adaptation to simulated “jet lag”. Am. J. Physiol. 1995, 269, E489–E498. [Google Scholar] [CrossRef]
- Lindenthal, B.; Simatupang, A.; Dotti, M.T.; Federico, A.; Lutjohann, D.; von Bergmann, K. Urinary excretion of mevalonic acid as an indicator of cholesterol synthesis. J. Lipid Res. 1996, 37, 2193–2201. [Google Scholar] [PubMed]
- Gnocchi, D.; Pedrelli, M.; Hurt-Camejo, E.; Parini, P. Lipids around the Clock: Focus on Circadian Rhythms and Lipid Metabolism. Biology 2015, 4, 104–132. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.J.; Pappu, A.S.; Hatcher, L.; Li, Z.C.; Illingworth, D.R.; Connor, W.E. Dietary cholesterol feeding suppresses human cholesterol synthesis measured by deuterium incorporation and urinary mevalonic acid levels. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 1222–1228. [Google Scholar] [CrossRef]
- Sakakura, Y.; Shimano, H.; Sone, H.; Takahashi, A.; Inoue, N.; Toyoshima, H.; Suzuki, S.; Yamada, N. Sterol regulatory element-binding proteins induce an entire pathway of cholesterol synthesis. Biochem. Biophys. Res. Commun. 2001, 286, 176–183. [Google Scholar] [CrossRef]
- Horton, J.D.; Bashmakov, Y.; Shimomura, I.; Shimano, H. Regulation of sterol regulatory element binding proteins in livers of fasted and refed mice. Proc. Natl. Acad. Sci. USA 1998, 95, 5987–5992. [Google Scholar] [CrossRef] [Green Version]
- Espenshade, P.J. SREBPs: Sterol-regulated transcription factors. J. Cell Sci. 2006, 119, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.K.; Yang, F.; Jiang, K.; Ji, J.Y.; Watts, J.L.; Purushotham, A.; Boss, O.; Hirsch, M.L.; Ribich, S.; Smith, J.J.; et al. Conserved role of SIRT1 orthologs in fasting-dependent inhibition of the lipid/cholesterol regulator SREBP. Genes Dev. 2010, 24, 1403–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Schachter, M. Chemical, pharmacokinetic and pharmacodynamic properties of statins: An update. Fundam. Clin. Pharmacol. 2005, 19, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Björkhem, I.; Miettinen, A.; Reihner, E.; Ewerth, S.; Angelin, B.; Einarsson, K. Correlation between serum levels of some cholesterol precursors and activity of HMG-CoA reductase in human liver. J. Lipid Res. 1987, 28, 1137–1143. [Google Scholar] [PubMed]
- Hallikainen, M.; Simonen, P.; Gylling, H. Cholesterol metabolism and serum non-cholesterol sterols: Summary of 13 plant stanol ester interventions. Lipids Health Dis. 2014, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Luu, W.; Hart-Smith, G.; Sharpe, L.J.; Brown, A.J. The terminal enzymes of cholesterol synthesis, DHCR24 and DHCR7, interact physically and functionally. J. Lipid Res. 2015, 56, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Acimovic, J.; Kosir, R.; Kastelec, D.; Perse, M.; Majdic, G.; Rozman, D.; Kosmelj, K.; Golicnik, M. Circadian rhythm of cholesterol synthesis in mouse liver: A statistical analysis of the post-squalene metabolites in wild-type and Crem-knock-out mice. Biochem. Biophys. Res. Commun. 2011, 408, 635–641. [Google Scholar] [CrossRef]
- Bae, S.H.; Paik, Y.K. Cholesterol biosynthesis from lanosterol: Development of a novel assay method and characterization of rat liver microsomal lanosterol delta 24-reductase. Biochem. J. 1997, 326, 609–616. [Google Scholar] [CrossRef]
- Hussain, M.M.; Pan, X. Circadian Regulation of Macronutrient Absorption. J. Biol. Rhythms 2015, 30, 459–469. [Google Scholar] [CrossRef]
- Altmann, S.W.; Davis, H.R., Jr.; Zhu, L.J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Gylling, H.; Vanhanen, H.; Ollus, A. Cholesterol absorption, elimination, and synthesis related to LDL kinetics during varying fat intake in men with different apoprotein E phenotypes. Arterioscler. Thromb. Vasc. Biol. 1992, 12, 1044–1052. [Google Scholar] [CrossRef]
- Reddy, B.S. Diet and excretion of bile acids. Cancer Res. 1981, 41, 3766–3768. [Google Scholar]
- Pooler, P.A.; Duane, W.C. Effects of bile acid administration on bile acid synthesis and its circadian rhythm in man. Hepatology 1988, 8, 1140–1146. [Google Scholar] [CrossRef]
- Kovar, J.; Lenicek, M.; Zimolova, M.; Vitek, L.; Jirsa, M.; Pitha, J. Regulation of diurnal variation of cholesterol 7alpha-hydroxylase (CYP7A1) activity in healthy subjects. Physiol. Res. 2010, 59, 233–238. [Google Scholar]
- Holt, J.A.; Luo, G.; Billin, A.N.; Bisi, J.; McNeill, Y.Y.; Kozarsky, K.F.; Donahee, M.; Wang, D.Y.; Mansfield, T.A.; Kliewer, S.A.; et al. Definition of a novel growth factor-dependent signal cascade for the suppression of bile acid biosynthesis. Genes Dev. 2003, 17, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Lundasen, T.; Galman, C.; Angelin, B.; Rudling, M. Circulating intestinal fibroblast growth factor 19 has a pronounced diurnal variation and modulates hepatic bile acid synthesis in man. J. Intern. Med. 2006, 260, 530–536. [Google Scholar] [CrossRef]
- Al-Khaifi, A.; Rudling, M.; Angelin, B. An FXR Agonist Reduces Bile Acid Synthesis Independently of Increases in FGF19 in Healthy Volunteers. Gastroenterology 2018, 155, 1012–1016. [Google Scholar] [CrossRef]
- Demacker, P.N.; Schade, R.W.; Jansen, R.T.; Van’t Laar, A. Intra-individual variation of serum cholesterol, triglycerides and high density lipoprotein cholesterol in normal humans. Atherosclerosis 1982, 45, 259–266. [Google Scholar] [CrossRef]
- Henderson, L.O.; Saritelli, A.L.; LaGarde, E.; Herbert, P.N.; Shulman, R.S. Minimal within-day variation of high density lipoprotein cholesterol and apolipoprotein A-I levels in normal subjects. J. Lipid Res. 1980, 21, 953–955. [Google Scholar]
- van Kerkhof, L.W.; Van Dycke, K.C.; Jansen, E.H.; Beekhof, P.K.; van Oostrom, C.T.; Ruskovska, T.; Velickova, N.; Kamcev, N.; Pennings, J.L.; van Steeg, H.; et al. Diurnal Variation of Hormonal and Lipid Biomarkers in a Molecular Epidemiology-Like Setting. PLoS ONE 2015, 10, e0135652. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, S.; Singh, R.K.; Cornelissen, G. Circadian Time Structure of Circulating Plasma Lipid Components in Healthy Indians of Different Age Groups. Indian J. Clin. Biochem. 2016, 31, 215–223. [Google Scholar] [CrossRef]
- Rivera-Coll, A.; Fuentes-Arderiu, X.; Diez-Noguera, A. Circadian rhythmic variations in serum concentrations of clinically important lipids. Clin. Chem. 1994, 40, 1549–1553. [Google Scholar]
- Jones, P.J.; Leitch, C.A.; Pederson, R.A. Meal-frequency effects on plasma hormone concentrations and cholesterol synthesis in humans. Am. J. Clin. Nutr. 1993, 57, 868–874. [Google Scholar] [CrossRef]
- Duffy, J.F.; Cain, S.W.; Chang, A.M.; Phillips, A.J.; Munch, M.Y.; Gronfier, C.; Wyatt, J.K.; Dijk, D.J.; Wright, K.P., Jr.; Czeisler, C.A. Sex difference in the near-24-h intrinsic period of the human circadian timing system. Proc. Natl. Acad. Sci. USA 2011, 108, 15602–15608. [Google Scholar] [CrossRef]
- Hofman, M.A.; Swaab, D.F. Living by the clock: The circadian pacemaker in older people. Ageing Res. Rev. 2006, 5, 33–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Author (year) | Study Design | Population | Intervention | n | Age (years) | Male (%) | Marker | Study Duration (Hours) | Dietary Information | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type of Meals | Breakfast | Lunch | Dinner | Other | |||||||||
| Kopito (1982) [28] | Experimental, crossover | Healthy subjects |
| 1 | 40 | 100 | MVA | 72 |
| 19:00 | |||
| Miettinen (1982) [29] | Longitudinal | Healthy subjects | 7 | Range: 16–49 | N/A | Squalene + methyl sterols | 24 | Three meals and evening snacks. Fat intake provided approximately 35% of total calories | 08:00 | 12:00 | 16:00 | 20:00 | |
| Parker (1982) [30] | Experimental, crossover | Subject with hyper-cholesterolemia and ischemic heart disease | 1 | 55 | 100 | MVA | 72 | Four daily liquid-formula meals in equal portions; caloric intakes adjusted to maintain BW within ± 1.5 kg over 3–5-weeks | 08:00 | 13:00 | 19:00 | 10:00 | |
| Subject with hypertri-glyceridemia, obesity, and maturity-onset diabetes |
| 1 | 66 | 100 | MVA | 48 |
| 08:00 | 13:00 | 19:00 | 10:00/17:00 | ||
| Subject with heterozygous familial hyper-cholesterolemia and ischemic heart disease |
| 1 | 45 | 100 | MVA | 48 |
| 08:00 | 13:00 | 19:00 | 10:00/17:00 | ||
| Parker (1984) [31] | Experimental, crossover | Subject with hypertri-glyceridemia |
| 1 | 68 | 100 | MVA | 72 |
| 08:00 | 13:00 | 19:00 | 10:00/17:00 |
| Miettinen (1985) [32] | Longitudinal | Subjects with jejunoileal bypass | 4 | 26 ± 8 | N/A | Squalene + methyl sterols | 24 | Low-cholesterol diet, 125 mg cholesterol/ 2400 kcal, 100 g fat/day | 08:00 | 12:00 | 16:00 | 20:00 | |
| Subjects with ileal exclusion | 4 | 38 ± 12 | N/A | ||||||||||
| Scoppola (1991) [33] | Longitudinal | Healthy subjects | 1 | N/A | N/A | MVA | 24 | Low fat (<5%), cholesterol-free meals | 09:30 | 12:30 | 19:00 | ||
| Jones (1992) [34] | Longitudinal | Healthy subjects | 5 | 26 ± 4 | 100 | MVA | 48 | Three self-selected, habitual meals/day for three days prior to and 48 h during the study | 08:00 –09:00 | 12:00 –13:00 | 18:00 –19:00 | ||
| Pappu (1994) [35] | Longitudinal | Healthy subjects | 6 | 30 ± 2 | 50 | MVA | 24 | Three meals/day (40% fat, 25% CHO, 15% protein) | 08:00 | 12:00 | 18:00 | ||
| Patients with abetalipoproteinemia | 3 | 24 ± 10 | 66.6 | Three meals/ day (12–15% fat, 70–75% CHO, 13–17% protein) | 08:00 | 12:00 | 18:00 | ||||||
| Yoshida (1994) [36] | Longitudinal | Patients with cholelithiasis and patients with early cancer of the GI-tract | 3 | Range: 24 – 28 | N/A | C4 | 24 | Normal hospital diets | 08:00 | 12:30 | 17:30 | ||
| Nozaki (1996) [37] | Experimental, crossover | Subjects with heterozygous familial hypercholesterolemia |
| 8 | 58 ± 9 | 37.5 | MVA | 24 | Cholesterol intake ± 300 mg/day; 20% fat intake; ratio polyunsaturated to saturated FAs was 1.5; single dose pravastatin (20 mg) taken after breakfast or after dinner | 08:00 | 12:00 | 18:00 | |
| Pappu (2002) [38] | Experimental, crossover | Subjects with heterozygous familial hypercholesterolemia |
| 9 5 | 41 ± 4 N/A | 0 | MVA | 24 | Low-cholesterol, low-fat diet conforming to phase I of the American Heart Association Diet; statins (40 mg) given after breakfast and dinner for eight weeks | 08:00 | 12:00 | 18:00 | |
| Martin (2002) [39] | Experimental, crossover | Healthy subjects |
| 21 | N/A | N/A | MVA | 24 | Individual caloric and fat intake was stabilized; Rosuvastatin (10 mg) taken each morning (~07:00 h) or evening (~18:00 h) for 14 days | N/A | N/A | N/A | |
| Gälman (2005) [40] | Longitudinal | Healthy subjects (n = 5) and cholecystecto-mized subjects (n = 3) | 8 | Range: 25–58 | 50 | C4, latho-sterol | 24 | Standardised meals | 09:00 | 12:00 | 18:00 | ||
| Persson (2010) [41] | Experimental | Healthy subjects |
| 10 | N/A | 75 | Latho-sterol | 33 | Standardised meals; CME was taken with meals day 1 (4 × 4 g) | 08:30 | 12:30 | 16:00 | 21:30 |
| Steiner (2011) [42] | Longitudinal | Healthy subjects | 4 | Range: 27–29 | 50 | C4 | 24 | Subjects consumed identical meals | 09:15 (day 1) 08:45 (day 2) | 11:30 | 20:15 | ||
| Al-Khaifi (2018) [43] | Experimental, crossover | Healthy subjects |
| 8 | Range: 20–45 | 100 | C4, latho-sterol | 32 | Standardised meals; CME (4 × 4 g) taken before meals day 1; atorvastatin (four daily doses of 40 mg) dose 1 and 2 taken on the morning two days before CME treatment, dose 3 and 4 on the morning of the first and second study day | 08:30 | 13:00 | 18:00 | 20:30 |
| Marker | Mean ± SD (µmol/mmol cholesterol) | Lathosterol | Desmosterol | Campesterol | Sitosterol |
|---|---|---|---|---|---|
| Lathosterol | 1.5 ± 0.3 | ||||
| Desmosterol | 1.0 ± 0.2 | 0.190 | |||
| Campesterol | 2.4 ± 0.8 | −0.521 * | −0.148 | ||
| Sitosterol | 1.9 ± 0.4 | −0.438 * | −0.309 | 0.868 ** | |
| Cholestanol | 1.6 ± 0.3 | −0.368 | −0.427 * | 0.755 ** | 0.853 ** |
| Marker | Mesor (SE) | Amplitude (SE) | Peak Time (Clock Hours) | Cosinor p |
|---|---|---|---|---|
| Cholesterol | 4.18 (0.06) | 0.19 (0.04) | 14:08 | 0.070 |
| Lathosterol | 1.52 (0.03) | 0.22 (0.02) | 02:47 | <0.001 |
| Desmosterol | 1.02 (0.01) | 0.02 (0.01) | 19:57 | 0.640 |
| Campesterol | 2.47 (0.06) | 0.04 (0.04) | 23:39 | 0.890 |
| Sitosterol | 1.97 (0.03) | 0.05 (0.02) | 01:57 | 0.460 |
| Cholestanol | 1.57 (0.02) | 0.01 (0.01) | 17:00 | 0.950 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroor, M.M.; Sennels, H.P.; Fahrenkrug, J.; Jørgensen, H.L.; Plat, J.; Mensink, R.P. Diurnal Variation of Markers for Cholesterol Synthesis, Cholesterol Absorption, and Bile Acid Synthesis: A Systematic Review and the Bispebjerg Study of Diurnal Variations. Nutrients 2019, 11, 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071439
Schroor MM, Sennels HP, Fahrenkrug J, Jørgensen HL, Plat J, Mensink RP. Diurnal Variation of Markers for Cholesterol Synthesis, Cholesterol Absorption, and Bile Acid Synthesis: A Systematic Review and the Bispebjerg Study of Diurnal Variations. Nutrients. 2019; 11(7):1439. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071439
Chicago/Turabian StyleSchroor, Maite M., Henriette P. Sennels, Jan Fahrenkrug, Henrik L. Jørgensen, Jogchum Plat, and Ronald P. Mensink. 2019. "Diurnal Variation of Markers for Cholesterol Synthesis, Cholesterol Absorption, and Bile Acid Synthesis: A Systematic Review and the Bispebjerg Study of Diurnal Variations" Nutrients 11, no. 7: 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071439