1. Introduction
The simple phenol hydroxytyrosol (HT) is the most abundant phenolic compound in extra virgin olive oil (EVOO) [
1] and thus one of the strongest antioxidants present in the Mediterranean diet [
2]. In 2006, the EUROLIVE clinical trial described a dose-dependent relationship between the EVOO phenolic fraction and beneficial health effects, particularly regarding protection against low density lipoprotein (LDL) oxidation [
3]. In 2011, the European Food Safety Authority (EFSA) released a health claim concerning the benefits of the daily ingestion of 5 mg of HT and its derivates in olive oil in the prevention of LDL oxidation [
4].
Fermented beverages such as wine and beer are sources of the simple phenol tyrosol (TYR) [
5]. TYR is produced during fermentation as a byproduct of tyramine metabolism and can be endogenously converted into HT in the human body after ingestion [
6]. This biotransformation is of relevance since in vitro studies have demonstrated that HT possesses higher antioxidant capacity than TYR [
7]. Both molecules share the same structure; however, TYR lacks a catechol group (-OH). The extra catechol group of HT enhances its capacity to stabilize free radicals. In the case of TYR, the single hydroxyl group does not provide any direct antioxidant activity [
8].
TYR hydroxylation to produce HT is catalyzed by two isoforms of cytochrome P450 (CYP):
CYP2A6 and
CYP2D6 [
9]. Their enzymatic activities present considerable inter-individual variation in the general population, primarily due to genetic polymorphisms [
10]. To date, few investigations have examined the impact of
CYP2A6 and
CYPD26 genotypic profiles on the metabolism of dietary phenolic compounds [
11].
TYR bioavailability and HT biotransformation have never been described within the context of beer consumption. Better knowledge of their metabolism would help to determine beer contribution to human antioxidant status. Therefore, we designed a study aimed at (i) confirming TYR absorption and biotransformation into HT within the context of beer consumption, (ii) determining the effect of alcohol present in beer on the absorption of TYR, and (iii) analyzing the impact of sex and CYP2A6/CYP2D6 polymorphisms on TYR conversion efficiency into HT.
4. Discussion
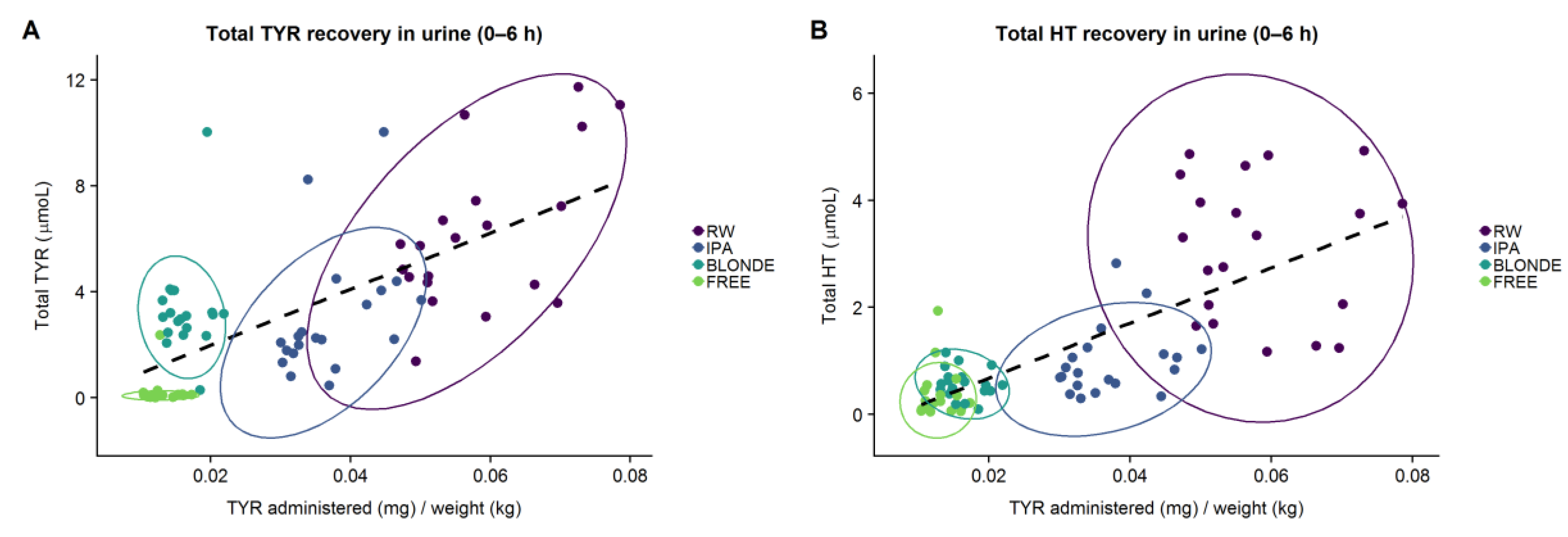
The present study demonstrates that, in humans, the TYR present in beer is well absorbed and endogenously biotransformed into HT. Our results also suggest that alcoholic content and beer matrix composition, in addition to sex and genetics, are factors that affect the metabolic disposition of TYR and HT, and could help to understand the variability observed in the metabolism of these simple phenols.
We first analyzed whether the TYR in beer was as bioavailable as that found in RW. When the same dose of alcohol was administered (RW vs. IPA beer), the recovery of TYR (corrected by doses administered) was lower after beer intake (22.4 vs. 17.4, percental recovery). As previously reported, blonde beer was the treatment with highest recovery of the administered TYR (50.5%; CI95% 39.5%–61.5%), in the form of both TYR and HT, followed by RW, IPA, and free beers, which would suggest a matrix effect on TYR absorption. Beers are rich in other polyphenols and their content is known to vary widely among styles [
19,
20,
21]. It is postulated that they could compete with TYR for their absorption, leading to a distinct TYR bioavailability. Remarkably, in our study, whilst free beer intake was associated with the lowest TYR recovery, the amount of TYR was similar to that present in the blonde beer although their alcohol content differed. Our findings confirm that the presence of alcohol is a key contributor to TYR bioavailability although not in a dose-dependent manner. The enhanced phenol absorption due to the presence of ethanol within the matrix has already been reported [
6].
The present results confirm beer as a source of HT even though its presence was almost undetectable. We observed a positive correlation between the ingested dose of TYR and HT urinary recovery. These findings further support the hypothesis of TYR being an endogenous precursor of HT in vivo. Nonetheless, a higher HT recovery adjusted by the given dose was described following RW (11.3%) compared to beer intake (5.6% to 7.5%). It is known that alcohol consumption produces a shift in dopamine metabolism, resulting in endogenous HT formation as a byproduct of dopamine in a dose-response manner to the alcohol consumed [
16]. This could explain part of the different HT recoveries following the intake of beverages with varying alcohol content. Nevertheless, the present differences in HT recovery suggest that TYR to HT conversion could be affected by other factors, as it was more efficient in RW than in IPA and blonde beers. A distinctive ingredient of beer is hops, especially rich in prenylated phenols. They have been reported to be potent and selective inhibitors of multiple members of the cytochrome P450 (CYP) family [
22]. Interestingly, different beer extracts have been studied for their inhibitory effect on
CYP2D6. Its activity was inhibited by all beer extracts with the greatest inhibition obtained with porter beer followed by ales whilst lager beers were the least inhibitors [
23]. There is no study assessing hop constituent effect on
CYP2A6. Based on these findings, hops present in beer could interfere in the TYR to HT conversion rate which would explain the lower production of HT observed following beer consumption and the lower impact of the genetic polymorphisms of these enzymes.
The urinary kinetics of TYR differs from HT in that the quantitative recovery of TYR is essentially concentrated in the first 0 to 4 h, while HT recovery is still relevant at 4 to 6 h. This is not surprising, since HT originates from the metabolic conversion of TYR into HT.
Our study confirms the extensive phase-II metabolism that TYR and HT undergo following their absorption. As has been described, once ingested, they go through an extensive first-pass hepatic metabolism, resulting in almost undetectable free forms in systemic biological fluids [
24]. In our study, free forms of HT and TYR were excreted at very low concentrations compared to their metabolites. Previous authors have suggested that the low concentrations of the free forms observed in vivo cannot explain the biological activities associated with phenolic consumption in clinical and epidemiological studies [
25]. Therefore, metabolites have been proposed as exhibiting relevant biological properties, either by having activity themselves or being deconjugated intracellularly giving rise to free active forms, as has been described for other dietary phenols [
26,
27]. In the present study we show an increase in TYR and HT phase II metabolite recovery following RW and beer treatments. The main metabolites recovered were TYR-4-sulfate, TYR-4-glucuronide, and HT-3-sulfate. They have been shown to possess in vitro biological activity equivalent to their parent compounds [
28,
29,
30,
31]. The fact that beer produced a rise in the same metabolites as RW suggests that it could trigger equivalent health effects.
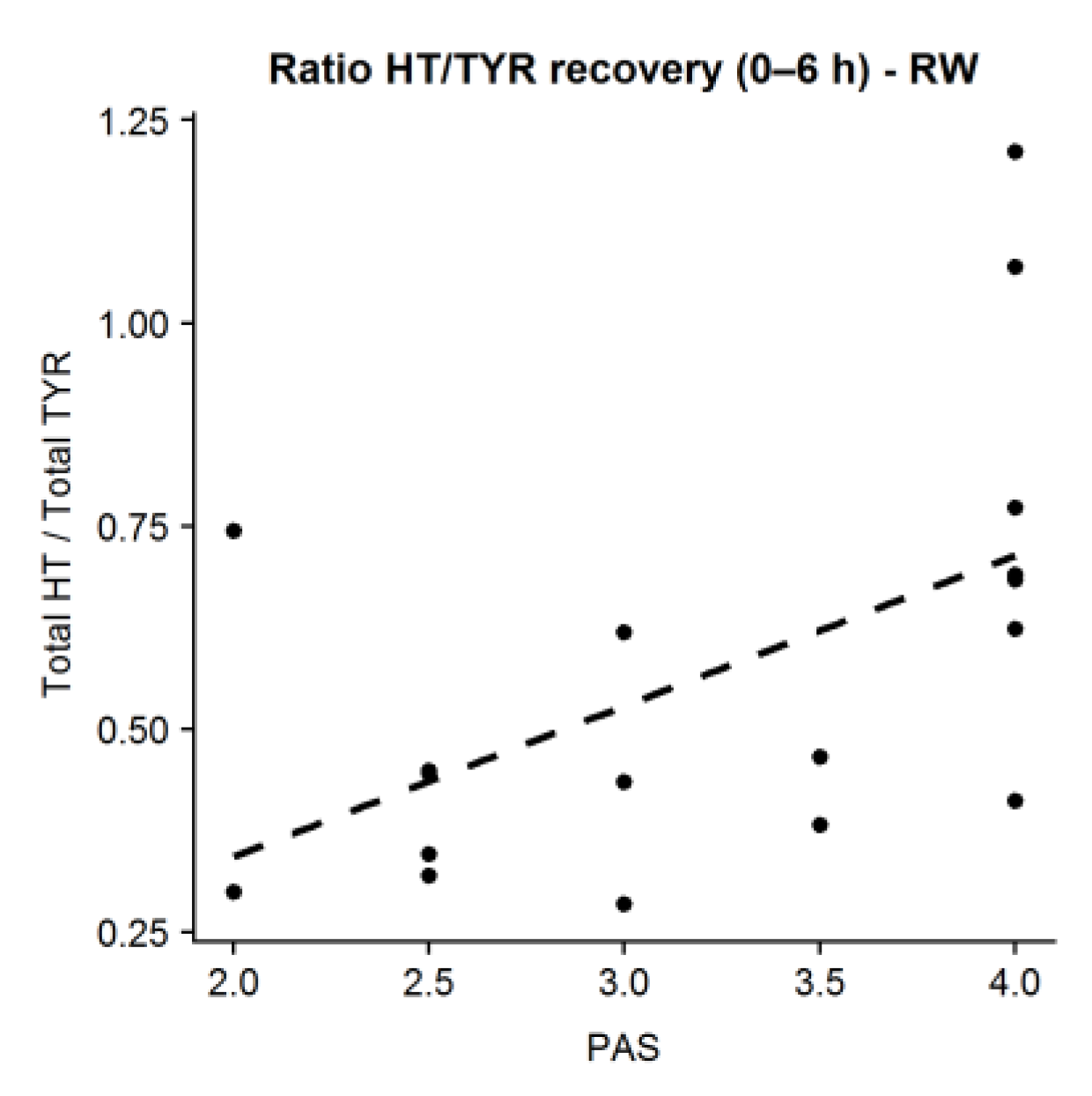
The current work provides further evidence of the mediation of
CYP2A6 and
CYP2D6 on TYR to HT biotransformation [
9,
11]. PAS scores positively correlated with the amount of HT recovered (higher PAS are related to higher HT/TYR ratios in urine) after RW intake. Results were only observable in the case of RW, probably due to its higher TYR absorbed dose and the different composition in polyphenols compared to beer, as other factors than genetics have been described to affect the final activity of
CYP2A6 and
CYP2D6 enzymes (such as certain drugs and dietary and endogenous components) [
10,
14].
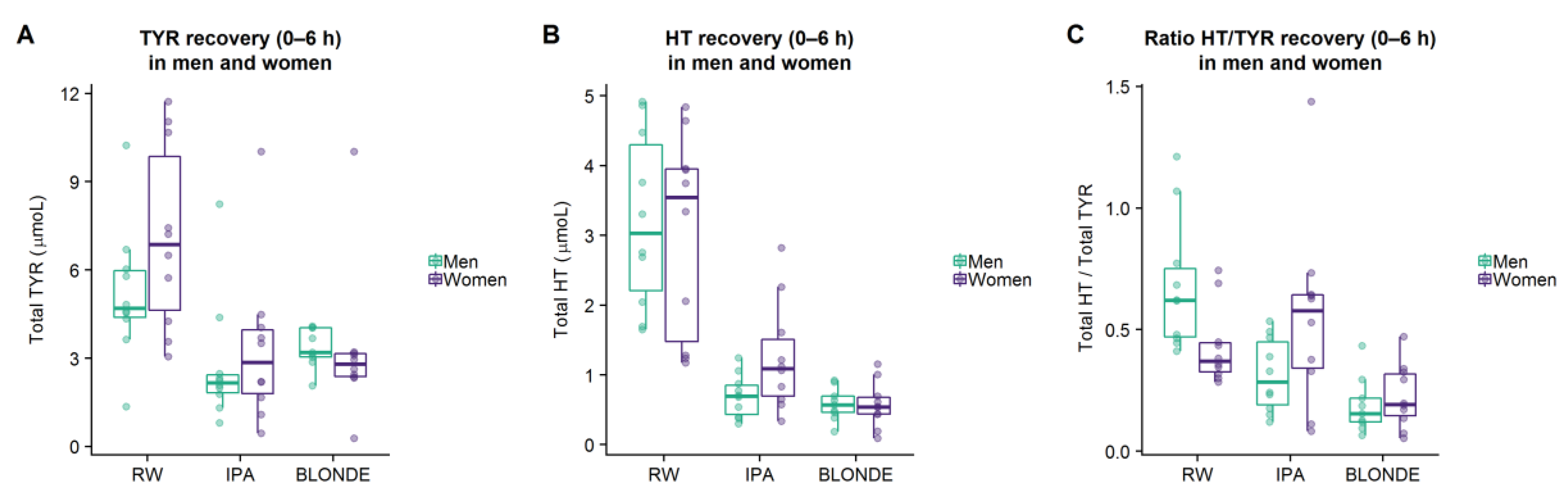
Sex differences in the metabolism of TYR and HT were observed in this study. Men were more efficient converting absorbed TYR to HT (higher HT/TYR ratio) after the administration of RW. This result is consistent with previous findings from the PREDIMED study, in which men presented higher values of HT adjusted by alcohol consumption [
32]. Another clinical study administering olive leaf extracts reported greater concentrations of HT metabolites in men compared to women in response to the same treatment [
33]. In our study, no significant sex differences were observed following IPA, blonde, and free beer treatments, suggesting that other factors could be affecting TYR to HT biotransformation capacity in women. It is well known that
CYP2A6 activity is upregulated by estrogen levels [
10], indicating a higher activity in women compared to men; however, this was not observed in our study. Nonetheless, the influence of estrogen status and the fluctuation intrinsic to the menstrual cycle could affect
CYP2A6 activity distinctly among the experimental sessions which may explain the absence of correlation among treatment ratios in women. Caution should be taken when interpreting these results as women presented lower PAS values than men.
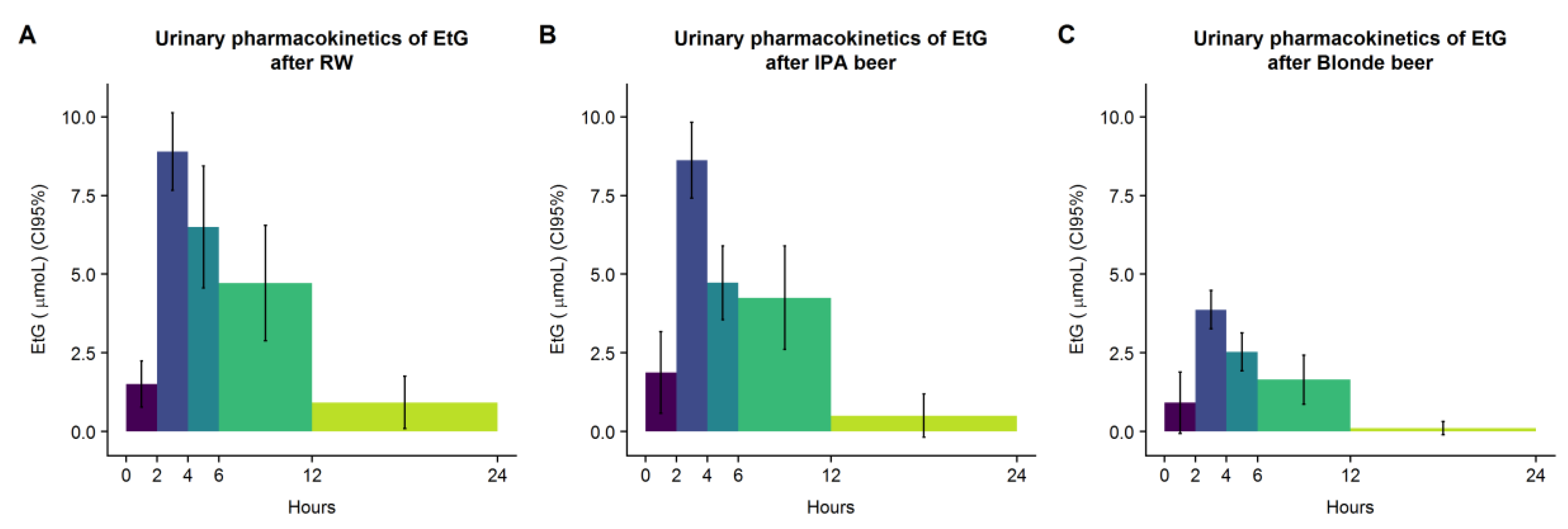
EtG is a widely used biomarker of alcohol consumption (and of compliance to alcohol abstention prior to experimental sessions) and as such was applied in this study [
34]. Its recovery following our treatments matched their alcohol content. Interestingly, marginal differences were observed between genders in EtG recovery, particularly for the IPA beer. This observation would be in line with the existence of variations in alcohol metabolism between men and women [
35].
Our findings enhance understanding of the antioxidant composition of beer and its potential health effects. Currently, beer is the most popular drink in the western world [
36], whilst RW consumption is decreasing. Nevertheless, at similar given doses of TYR and equal doses of alcohol, RW exhibited a higher recovery on TYR and HT metabolites, suggesting a better bioavailability of TYR and a more efficient conversion to HT. Therefore, it is likely that the beneficial effects attributed to RW moderate consumption are superior to the observed following beer consumption. A large body of evidence exists about the health effects attributed to moderate RW consumption, whilst less is known about moderate beer consumption, and data are mostly derived from epidemiological studies [
37]. The study of traditional beer and non-alcoholic beer can help to understand the role of ethanol and its interaction with beer polyphenols and their biological effects. Beer alcoholic content is lower than RW and, as shown in the present study, beer consumption triggered an increase in similar phenols. Non-alcoholic beer led to a mild increment in TYR and HT metabolites due to its lower bioavailability. Nevertheless, it could represent a source of these compounds in cases where alcohol consumption has to be limited.
Our study presents certain strengths and limitations. Firstly, the cross-over design enabled the observation of intra-individual changes, control for potential confounders, and between-subject variation. Secondly, we also evaluated the presence of sex differences in the metabolism of dietary phenolic compounds. This approach goes beyond traditional medical and nutritional studies that have long been performed in men and translated directly to women. Another strength of this study is that the administered doses of RW and beers are achievable in a normal diet and in agreement with moderate alcohol consumption. The current investigation was; however, limited by the low sample size which hampered the simultaneous grouping and comparison of volunteers according to their sex and PAS. This would have been relevant since PAS was not equally distributed between the genders. In addition, treatments were given at a single dose; it is possible that longer administration would have resulted in different results attributed to sex or PAS. Finally, further studies should be performed to assess the relevance of the clinical effects of TYR and endogenous HT formation associated with moderate beer consumption, at a postprandial level, and after chronic consumption.







{kind=link}
{kind=link}
{kind=link}
{kind=link}