Immunomodulatory Effects of Fruiting Body Extract and Solid-State-Cultivated Mycelia of Taiwanofungus camphoratus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Sample Preparation
2.2. Animal Treatment
2.3. Cell Proliferation Assay
2.4. NK Cell Activity
2.5. Phagocytotic Activity
2.6. Cytokine Analysis
2.7. Antibody Production Test
2.8. Statistical Analysis
3. Results
3.1. Effect of LDAC on the Proliferation of Spleen Immune Cells
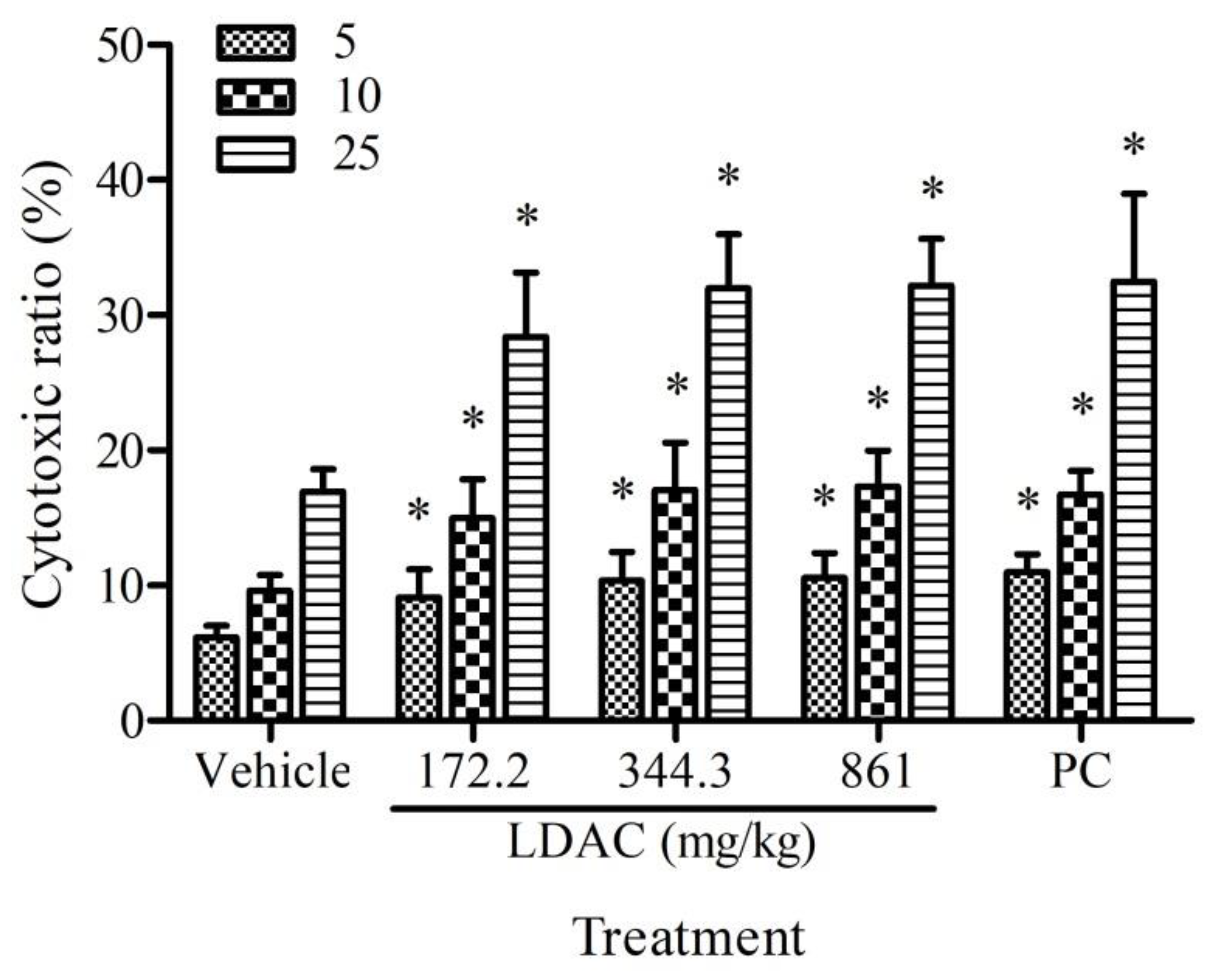
3.2. Effect of LDAC on NK Cell Activity
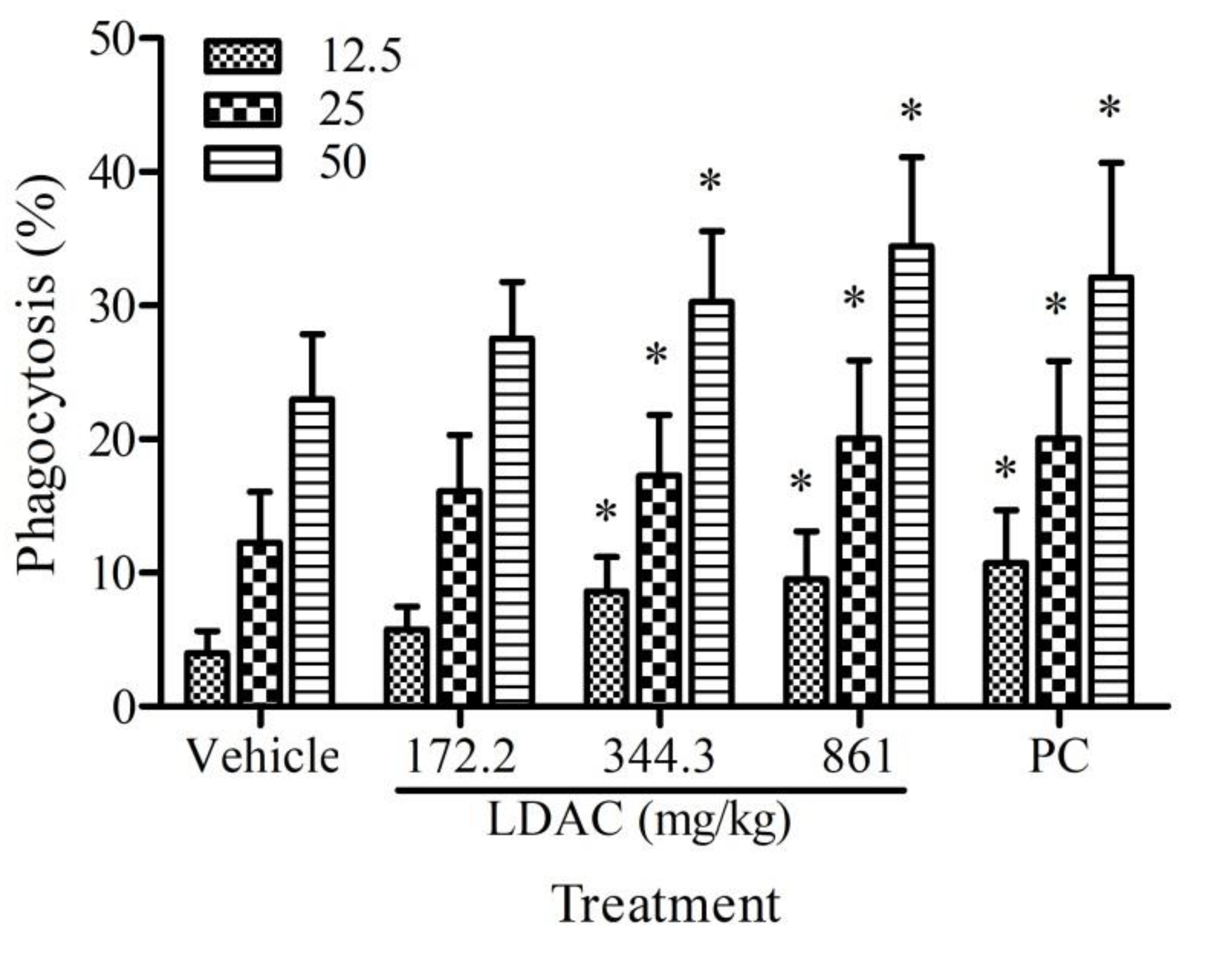
3.3. Effect of LDAC on Phagocyte Activity
3.4. Effect of LDAC on Cytokine Levels
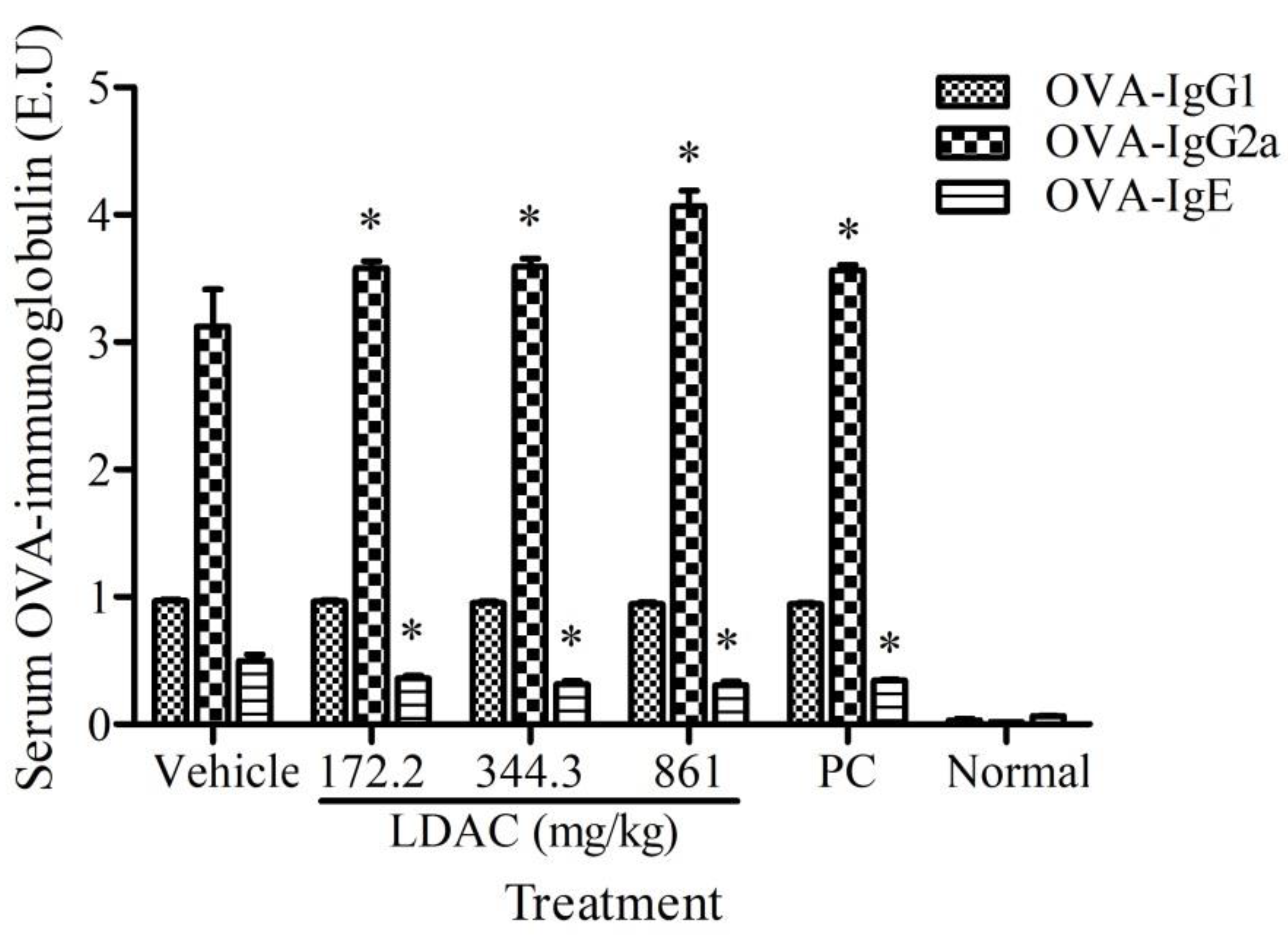
3.5. Effect of LDAC on Serum Antibody Secretion
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaminogawa, S.; Nanno, M. Modulation of Immune Functions by Foods. Evid. Based Complement. Alternat. Med. 2004, 1, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Hsu, Y.J.; Chang, C.J.; Lin, C.S.; Martel, J.; Ojcius, D.M.; Ko, Y.F.; Lai, H.C.; Young, J.D. Immunomodulatory properties of medicinal mushrooms: Differential effects of water and ethanol extracts on NK cell-mediated cytotoxicity. Innate Immun. 2016, 22, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Ryvarden, L.; Chang, T.T. Antrodia camphorata (‘niu-chang-chih’), new combination of a medicinal fungus in Taiwan. Bot. Bull. Acad. Sin. 1997, 38, 273–275. [Google Scholar]
- Geethangili, M.; Tzeng, Y.M. Review of Pharmacological Effects of Antrodia camphorata and Its Bioactive Compounds. Evid. Based Complement. Alternat. Med. 2011, 2011, 212641. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.Y.; Wong, Y.Y.; Chan, T.Y.; Law, C.K.; Tsoi, Y.K.; Leung, K.S. Review of biological and pharmacological activities of the endemic Taiwanese bitter medicinal mushroom, Antrodia camphorata (M. Zang et C. H. Su) Sh. H. Wu et al. (higher Basidiomycetes). Int. J. Med. Mushrooms 2012, 14, 241–256. [Google Scholar] [CrossRef]
- Huang, Y.L.; Chu, Y.L.; Ho, C.T.; Chung, J.G.; Lai, C.I.; Su, Y.C.; Kuo, Y.H.; Sheen, L.Y. Antcin K, an Active Triterpenoid from the Fruiting Bodies of Basswood-Cultivated Antrodia cinnamomea, Inhibits Metastasis via Suppression of Integrin-Mediated Adhesion, Migration, and Invasion in Human Hepatoma Cells. J. Agric. Food Chem. 2015, 63, 4561–4569. [Google Scholar] [CrossRef]
- Kumar, K.J.; Vani, M.G.; Chueh, P.J.; Mau, J.L.; Wang, S.Y. Antrodin C inhibits epithelial-to-mesenchymal transition and metastasis of breast cancer cells via suppression of Smad2/3 and beta-catenin signaling pathways. PLoS ONE 2015, 10, e0117111. [Google Scholar]
- Wang, G.; Wan, Y.; Zhao, J.; Hong, Z. Ethanol extract of Antrodia camphorata inhibits proliferation of HCT-8 human colorectal cancer cells by arresting cell cycle progression and inducing apoptosis. Mol. Med. Rep. 2017, 16, 4941–4947. [Google Scholar] [CrossRef]
- Li, Z.W.; Kuang, Y.; Tang, S.N.; Li, K.; Huang, Y.; Qiao, X.; Yu, S.W.; Tzeng, Y.M.; Lo, J.Y.; Ye, M. Hepatoprotective activities of Antrodia camphorata and its triterpenoid compounds against CCl4-induced liver injury in mice. J. Ethnopharmacol. 2017, 206, 31–39. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Li, L.; Hu, W.; Qu, Y.; Ding, Y.; Meng, L.; Teng, L.; Wang, D. Hepatoprotective Effects of Antrodia cinnamomea: The Modulation of Oxidative Stress Signaling in a Mouse Model of Alcohol-Induced Acute Liver Injury. Oxid. Med. Cell. Longev. 2017, 2017, 7841823. [Google Scholar] [CrossRef]
- Shie, P.H.; Wang, S.Y.; Lay, H.L.; Huang, G.J. 4,7-Dimethoxy-5-methyl-1,3-benzodioxole from Antrodia camphorata inhibits LPS-induced inflammation via suppression of NF-kappaB and induction HO-1 in RAW264.7 cells. Int. Immunopharmacol. 2016, 31, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.Y.; Liu, Y.C.; Kuo, Y.H.; Lin, Y.L.; Wu, Y.S.; Chen, J.W.; Chen, Y.C. Effects of antrosterol from Antrodia camphorata submerged whole broth on lipid homeostasis, antioxidation, alcohol clearance, and anti-inflammation in livers of chronic-alcohol fed mice. J. Ethnopharmacol. 2017, 202, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Cheng, P.C.; Lin, C.N.; Liao, H.F.; Chen, Y.Y.; Chen, C.C.; Lee, K.M. Polysaccharides from Antrodia camphorata mycelia extracts possess immunomodulatory activity and inhibits infection of Schistosoma mansoni. Int. Immunopharmacol. 2008, 8, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.C.; Chang, C.Y.; Cheng, T.L.; Wu, M.J. Immunomodulatory effect of Antrodia camphorata mycelia and culture filtrate. J. Ethnopharmacol. 2008, 120, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.C.; Hwang, S.L.; Chang, F.R.; Chen, Y.H.; Chang, T.T.; Hung, C.S.; Wang, C.L.; Chu, Y.H.; Pan, S.H.; Wu, Y.C. Immunostimulatory effect of Antrodia camphorata extract on functional maturation of dendritic cells. Food Chem. 2009, 113, 1049–1057. [Google Scholar] [CrossRef]
- Liu, K.J.; Leu, S.J.; Su, C.H.; Chiang, B.L.; Chen, Y.L.; Lee, Y.L. Administration of polysaccharides from Antrodia camphorata modulates dendritic cell function and alleviates allergen-induced T helper type 2 responses in a mouse model of asthma. Immunology 2010, 129, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Lo, C.P.; Lin, C.C.; Hsieh, Y.H. Effects of Taiwanofungus camphoratus on non-specific and specific immune activities in mice. Mycology 2018, 9, 129–135. [Google Scholar] [CrossRef]
- Lin, C.C.; Kumar, K.J.S.; Liao, J.W.; Kuo, Y.H.; Wang, S.Y. Genotoxic, teratotoxic and oral toxic assessments of Antrodia cinnamomea health food product (Leader Deluxe Antrodia cinnamomea (R)). Toxicol. Rep. 2015, 2, 1409–1417. [Google Scholar] [CrossRef]
- Lin, Y.H.; Kuo, J.T.; Chen, Y.Y.; Kumar, K.J.S.; Lo, C.P.; Lin, C.C.; Wang, S.Y. Immunomodulatory Effects of the Stout Camphor Medicinal Mushroom, Taiwanofungus camphoratus (Agaricomycetes)-Based Health Food Product in Mice. Int. J. Med. Mushrooms 2018, 20, 849–858. [Google Scholar] [CrossRef]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clinic. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Cheng, P.C.; Hsu, C.Y.; Chen, C.C.; Lee, K.M. In vivo immunomodulatory effects of Antrodia camphorata polysaccharides in a T1/T2 doubly transgenic mouse model for inhibiting infection of Schistosoma mansoni. Toxicol. Appl. Pharmacol. 2008, 227, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Pan, I.H.; Li, Y.R.; Pan, Y.G.; Lin, M.K.; Lu, Y.H.; Wu, H.C.; Chu, C.L. The adjuvant effects of high-molecule-weight polysaccharides purified from Antrodia cinnamomea on dendritic cell function and DNA vaccines. PLoS ONE 2015, 10, 0116191. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mg/kg) | Stimulation Index (S.I.) | ||
|---|---|---|---|---|
| Con A 5 μg/mL | LPS 10 μg/mL | OVA 25 μg/mL | ||
| Non-specific immune test | ||||
| Vehicle | - | 1.98 ± 0.19 | 2.15 ± 0.30 | - |
| LDAC | 172.2 | 2.22 ± 0.40 | 2.23 ± 0.31 | - |
| LDAC | 344.4 | 2.42 ± 0.72 * | 2.54 ± 0.53 | - |
| LDAC | 861 | 2.95 ± 0.57 * | 2.66 ± 0.54 * | - |
| PC | 633.5 | 2.78 ± 0.40 * | 2.86 ± 0.68 * | - |
| Specific immune test | ||||
| Vehicle | - | 2.88 ± 0.25 | 2.47 ± 0.34 | 1.35 ± 0.18 |
| LDAC | 172.2 | 3.40 ± 0.35 * | 2.72 ± 0.26 | 1.61 ± 0.28 * |
| LDAC | 344.4 | 3.48 ± 0.18 * | 2.78 ± 0.25 * | 1.62 ± 0.31 * |
| LDAC | 861 | 3.68 ± 0.44 * | 2.92 ± 0.31 * | 1.62 ± 0.21 * |
| PC | 633.5 | 3.71 ± 0.21 * | 3.30 ± 0.29 * | 1.69 ± 0.37 * |
| Group | Dose (mg/kg) | Con A (7.5 μg/mL) | LPS (15 μg/mL) |
|---|---|---|---|
| IL-2, pg/mL | |||
| Vehicle | 4545.2 ± 672.3 | 44.4 ± 7.4 | |
| LDAC | 172.2 | 5227.6 ± 646.9 * | 48.7 ± 5.3 |
| LDAC | 344.4 | 5257.9 ± 932.0 * | 52.7 ± 7.9 |
| LDAC | 861 | 5333.2 ± 219.2 * | 54.0 ± 8.6 |
| PC | 633.5 | 5152.9 ± 602.8 | 53.4 ± 17.0 |
| IL-4, pg/mL | |||
| Vehicle | 237.2 ± 63.9 | 10.4 ± 1.7 | |
| LDAC | 172.2 | 189.1 ± 38.3 * | 8.7 ± 1.1 * |
| LDAC | 344.4 | 171.6 ± 45.2 * | 8.1 ± 1.2 * |
| LDAC | 861 | 149.6 ± 28.3 * | 6.6 ± 1.3 * |
| PC | 633.5 | 162.7 ± 29.1 * | 8.3 ± 1.0 * |
| IL-5, pg/mL | |||
| Vehicle | 197.7 ± 50.9 | 31.8 ± 6.3 | |
| LDAC | 172.2 | 137.7 ± 20.0 * | 25.1 ± 1.7 * |
| LDAC | 344.4 | 111.1 ± 37.4 * | 24.9 ± 2.1 * |
| LDAC | 861 | 98.6 ± 9.0 * | 23.4 ± 3.2 * |
| PC | 633.5 | 101.4 ± 25.1 * | 29.0 ± 3.1 |
| TNF-α, pg/mL | |||
| Vehicle | 208.9 ± 40.0 | 611.1 ± 183.1 | |
| LDAC | 172.2 | 185.2 ± 21.1 | 594.7 ± 107.4 |
| LDAC | 344.4 | 171.5 ± 27.0 * | 548.6 ± 95.2 |
| LDAC | 861 | 167.6 ± 29.5 * | 523.4 ± 138.7 |
| PC | 633.5 | 193.6 ± 35.2 | 507.2 ± 95.1 |
| IFN-γ, pg/mL | |||
| Vehicle | 5512.6 ± 1920.7 | 2267.7 ± 807.1 | |
| LDAC | 172.2 | 6128.6 ± 1744.5 | 3061.3 ± 962.6 |
| LDAC | 344.4 | 7912.1 ± 1745.1 * | 4605.2 ± 1472.2 * |
| LDAC | 861 | 8745.2 ± 2176.4 * | 5038.4 ± 1874.5 * |
| PC | 633.5 | 8439.9 ± 2441.8 * | 4304.0 ± 1677.9 * |
| Group | Dose (mg/kg) | Con A (5 μg/mL) | LPS (10 μg/mL) | OVA (25 μg/mL) |
|---|---|---|---|---|
| IL-2 (pg/mL) | ||||
| Vehicle | - | 4808.3 ± 957.7 | 94.5 ± 26.6 | 205.1 ± 38.4 |
| LDAC | 172.2 | 5399.7 ± 704.7 | 107.5 ± 20.7 | 234.3 ± 31.6 |
| LDAC | 344.4 | 5464.6 ± 642.3 | 116.6 ± 31.5 | 217.2 ± 45.4 |
| LDAC | 861 | 5777.5 ± 1277.5 * | 125.8 ± 31.5 * | 205.2 ± 43.0 |
| PC | 633.5 | 5519.0 ± 632.5 | 87.8 ± 16.9 | 205.9 ± 50.6 |
| Normal | - | 3366.8 ± 373.3 | 37.3 ± 9.7 | 38.5 ± 16.5 |
| IL-4 (pg/mL) | ||||
| Vehicle | - | 48.5 ± 22.4 | 4.3 ± 0.4 | 5.0 ± 1.1 |
| LDAC | 172.2 | 33.2 ± 18.1 | 3.5 ± 0.5 * | 4.0 ± 0.9 * |
| LDAC | 344.4 | 34.3 ± 16.1 | 3.3 ± 0.3 * | 3.5 ± 0.5 * |
| LDAC | 861 | 22.8 ± 11.3 * | 3.5 ± 0.2 * | 3.2 ± 0.3 * |
| PC | 633.5 | 19.9 ± 7.6 * | 3.4 ± 0.5 * | 3.4 ± 0.7 * |
| Normal | - | 44.5 ± 18.0 | 3.4 ± 0.3 | 2.6 ± 0.4 |
| IL-5 (pg/mL) | ||||
| Vehicle | - | 65.0 ± 12.3 | 14.2 ± 4.7 | 8.4 ± 2.5 |
| LDAC | 172.2 | 57.0 ± 16.7 | 11.1 ± 6.5 | 7.7 ± 2.9 |
| LDAC | 344.4 | 63.2 ± 14.9 | 10.2 ± 3.3 | 6.6 ± 1.5 * |
| LDAC | 861 | 58.9 ± 10.0 | 10.7 ± 2.8 | 6.5 ± 1.4 * |
| PC | 633.5 | 53.8 ± 8.0 | 11.9 ± 5.7 | 5.9 ± 0.7 * |
| Normal | - | 64.0 ± 21.3 | 13.5 ± 5.5 | 4.7 ± 0.4 |
| TNF-α (pg/mL) | ||||
| Vehicle | - | 206.1 ± 84.5 | 583.9 ± 110.2 | 13.7 ± 6.8 |
| LDAC | 172.2 | 192.7 ± 71.0 | 430.2 ± 81.9 * | 10.6 ± 5.8 |
| LDAC | 344.4 | 180.9 ± 58.3 | 396.5 ± 99.3 * | 8.9 ± 4.7 |
| LDAC | 861 | 177.7 ± 78.4 | 394.4 ± 54.5 * | 6.1 ± 4.7 * |
| PC | 633.5 | 137.3 ± 73.0 | 419.6 ± 117.3 * | 10.1 ± 6.1 |
| Normal | - | 140.6 ± 48.3 | 364.9 ± 76.1 | 4.9 ± 1.6 |
| IFN-γ (pg/mL) | ||||
| Vehicle | - | 5128.0 ± 1927.1 | 1389.8 ± 736.4 | 1606.5 ± 758.9 |
| LDAC | 172.2 | 6649.7 ± 2202.2 | 1406.1 ± 199.5 | 1952.8 ± 958.3 |
| LDAC | 344.4 | 6970.9 ± 3132.8 | 1493.7 ± 255.2 | 1920.5 ± 661.3 |
| LDAC | 861 | 7139.3 ± 2550.0 | 1465.9 ± 192.4 | 1897.4 ± 751.3 |
| PC | 633.5 | 6525.7 ± 1991.0 | 1444.1 ± 626.0 | 1814.1 ± 360.9 |
| Normal | - | 4233.7 ± 1487.3 | 1274.0 ± 452.0 | 1050.6 ± 481.0 |
| Group | Dose (mg/kg) | IgG (μg/mL) | IgM (μg/mL) | IgE (μg/mL) |
|---|---|---|---|---|
| Vehicle | - | 511.8 ± 44.9 | 423.6 ± 80.4 | 0.29 ± 0.11 |
| LDAC | 172.2 | 629.1 ± 70.1 * | 428.5 ± 62.4 | 0.17 ± 0.04 * |
| LDAC | 344.4 | 687.6 ± 63.4 * | 481.1 ± 149.8 | 0.14 ± 0.04 * |
| LDAC | 861 | 669.7 ± 73.5 * | 447.7 ± 79.0 | 0.15 ± 0.07 * |
| PC | 633.5 | 665.0 ± 38.1 * | 434.5 ± 91.4 | 0.13 ± 0.05 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.-H.; Chi, C.-H.; Zhang, X.-H.; Chen, Y.-J.; Wang, M.-F. Immunomodulatory Effects of Fruiting Body Extract and Solid-State-Cultivated Mycelia of Taiwanofungus camphoratus. Nutrients 2019, 11, 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092256
Lin L-H, Chi C-H, Zhang X-H, Chen Y-J, Wang M-F. Immunomodulatory Effects of Fruiting Body Extract and Solid-State-Cultivated Mycelia of Taiwanofungus camphoratus. Nutrients. 2019; 11(9):2256. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092256
Chicago/Turabian StyleLin, Liang-Hung, Ching-Hsin Chi, Xiao-Han Zhang, Ying-Ju Chen, and Ming-Fu Wang. 2019. "Immunomodulatory Effects of Fruiting Body Extract and Solid-State-Cultivated Mycelia of Taiwanofungus camphoratus" Nutrients 11, no. 9: 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092256